
基于环境因子的渔业资源评估研究进展
2024-04-01 01:50鲁红月陈新军
海洋湖沼通报 2024年1期
鲁红月,张 畅,陈新军,2,3,4,5*
(1.上海海洋大学海洋科学学院,上海 201306;2.青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室,山东 青岛 266071;3.上海海洋大学大洋渔业资源可持续开发教育部重点实验室,上海 201306;4.上海海洋大学国家远洋渔业工程技术研究中心,上海 201306;5.上海海洋大学农业农村部大洋渔业开发重点实验室,上海 201306)
引 言
海洋渔业资源的开发和保护是众多专家学者一直以来所关注的重点话题,海洋环境对渔业资源的影响研究已成为国际上的热点话题[1]。环境因子对鱼类的影响贯穿整个生活史过程,补充量、渔获量和资源量等都深受其影响[2]。研究环境对渔业资源的影响,应对不断变化的海洋环境,建立更科学的渔业资源评估模型,更好的开发和利用渔业资源[3]。
评估模型是对渔业资源进行定量评估的重要工具,科学有效的资源评估,是对渔业资源合理管理,实现渔业资源可持续利用的重要手段。在交叉学科和计算机信息技术的支持下,资源评估模型不断改善,目前国内外学者将环境因子与模型中的单位捕捞努力量渔获量(Catch per unit effort,CPUE)[4-5]、资源承载能力联系起来取得较好的效果。基于环境因子的评估模型将成为未来渔业资源评估的重要发展趋势之一。为此,本研究将基于已有研究结果,将环境对鱼类早期生活史的影响、环境因子与模型参数的关系及渔业资源评估模型改进和应用进行分析,以期为我国渔业资源评估与管理学科的发展提供参考[6-7]。
1 环境对渔业资源的影响
1.1 环境对鱼类早期生活史的影响
环境变化对鱼类资源的影响是通过对其生活史过程的影响来实现的,鱼类在精卵结合到衰老死亡整个生命过程中的胚胎期、仔鱼期和稚鱼期,统称为鱼类早期生活史阶段。该阶段的鱼类作为补充群体,直接关系到物种年际补充量大小,是造成种群数量变动的主要原因,对整个资源群体有很重大意义,且各发育期与环境因子的联系各具特色[8]。
鱼类早期生活史阶段相比其他阶段对环境的敏感性高,产卵场分布、摄食、个体生长发育、自然死亡等极易受环境影响。该阶段自然死亡发生率较高,直接影响资源补充量多少,对资源量大小造成影响。自然死亡率除年龄、生长率、性成熟、摄食因素外,与海表温度(Sea surface temperature,SST)、海表面盐度(Sea surface salinity,SSS)等其他海洋环境密切相关[9]。产卵场温度的变化影响物种胚胎发育、生长和补充量,产卵场盐度变化影响物种的洄游路径和索饵场的分布。研究环境对鱼类早期生活史的具体影响形式,量化环境在不同生活史阶段造成的差别,才能更准确评估资源量[10-11]。
目前资源评估中,产卵场的环境与补充量关系得到较多的关注,对早期生活史阶段的影响机理研究较少。为可持续利用和科学管理渔业资源,需考虑环境变化对产卵场生物量的影响,重视环境对其幼体、仔稚鱼等不同生命阶段中的不同影响。
1.2 案例分析
以北太平洋柔鱼(Ommastrephesbartramii)为例,早期生活史阶段会造成不同阶段性影响:柔鱼仔稚鱼至成鱼期,其生长速率、个体大小、洄游模式、同类相食和死亡率等受环境的影响[15],胚胎期至仔鱼期,其孵化时间、卵存活率和孵化成功率受环境影响[12],产卵期至孵化期,其产卵时间、产卵地点、产卵数量和死亡率受影响[13]。
Vijai等[14]认为柔鱼早期生活史阶段鱼类死亡对资源补充量影响较大。Cao等[15]对北太平洋的冬生群体西部补充量变化的研究中,认为SST影响补充量的分布,补充群体更多分布在适宜SST范围。ICHII等[16]认为在过渡区叶绿素浓度(Chlorophyllconcentration,Chl-a)较高生产力海区是资源群体最为有利的育肥场和索饵场,且产卵场的位置随着Chl-a浓度的变化而变化[17-18]。陈新军等[19]利用北太平洋柔鱼冬春生西部群体产卵场和索饵场的适宜SST范围,解释了柔鱼资源量变化。除Chl-a、SST和SSS外,黑潮势力和路径的变化以及亲潮交汇的此消彼长,厄尔尼诺和拉尼娜等异常天气现象对柔鱼资源变动产生影响。
除北太平洋柔鱼外,产卵场SST变化对福克兰群岛附近的巴塔哥尼亚枪乌贼(Doryteuthis gahi)[20]、西北大西洋皮氏枪乌贼(Doryteuthispealeii)和滑柔鱼(Illex illecebrosus)[21]、茎柔鱼(Dosidicusgigas)[22]、太平洋褶柔鱼(Ommastrephes pacificus)[23]的胚胎发育、生长和补充量,以及在索饵场的分布均产生影响。
2 基于环境因子的渔业资源评估模型
2.1 剩余产量模型
(1)模型构建的理论基础和假设条件
剩余产量模型在评估中忽略种群年龄和体长等演化特征,将其作为一个综合独立的聚合单元。其所需最少数据为时间序列上的相对资源丰度指数和渔获量相关数据[8,24]。该模型以种群增长S型曲线一般模型为理论基础,将资源群体的生长和自然死亡综合起来作为资源群体大小的一个单变量函数进行分析[25]。常假设物种资源丰度受环境影响存在年度波动且模型参数恒定。
模型构建修正的参数如表1,该模型最常用的是Schaefer模型。Schaefer模型以逻辑斯蒂(logistic)方程为基础,捕捞死亡率与资源量呈线性关系。Schaefer模型[26-30]:
(1)
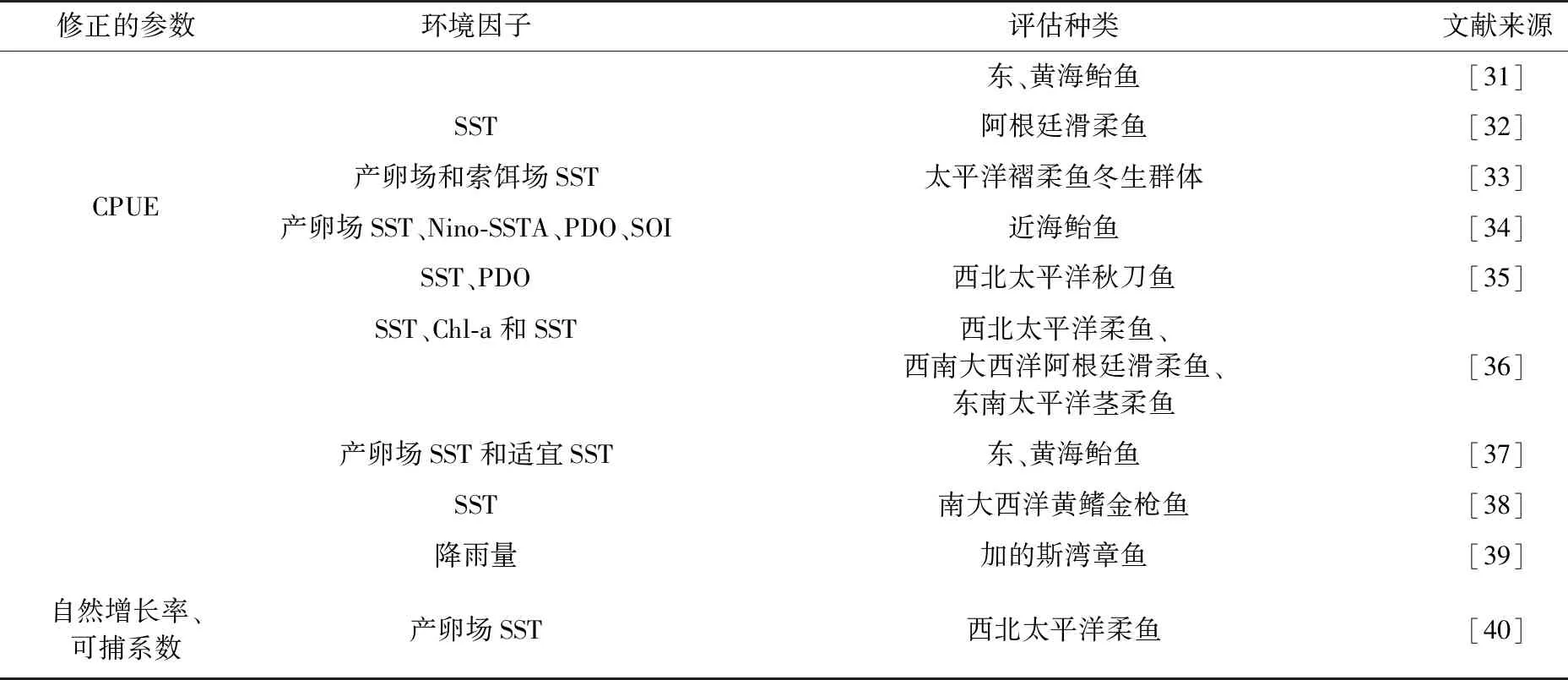
表1 基于环境因子的剩余产量模型参数统计表
其中,Bt代表t年资源量;K为承载力;r是指内禀自然增长率;Ct-1为t-1年的渔获量;r、K为模型中的参数。在模型中,常假设为常数而影响评估结果,实际中的参数尤其是渔获量、自然增长率是伴随着生活史过程进行动态变化。
(2)对评估模型的修正
模型评估中克服无法反映环境对鱼类种群动态变化的缺点[26-27],将环境因子引入评估模型中,具体因子如表1所示。多环境指数与CPUE、渔获量和捕捞努力量等用交相关分析、logistic回归分析、灰色系统的关联分析等进行选定,提高模型拟合程度[28]。方差、均方差、均方根、相关系数R2和赤池信息量(Akaike information criterion, AIC)用于评价模型拟合结果[41]。贝叶斯理论较充分的考虑模型与参数值的不确定性,降低后期管理策略实施中的风险。
(3)修正模型的应用
该模型强调环境与资源补充量和资源量的影响,考虑环境变化后模型拟合效果较好。具体应用见表1,模型多应用于近海和远洋头足类,还有白虾和其他鱼类。
2.2 年龄结构模型
(1)模型构建的理论基础和假设条件
年龄结构模型可结合多个数据源且考虑生物和环境过程的复杂情况,允许种群具有空间结构,并用海洋环境数据建立与资源补充量等参数的关系。需假设所有年份有一个恒定的捕捞水平且捕捞死亡率逐年不变并具有相同的捕捞能力。该模型应用较多的是:Stock synthesis(SS3为最新版本)模型。Stock synthesis模型表达式如下:
(2)
式中,Na,1是在年份1中年龄a的数量;amax是建模的最大年龄;M为瞬时自然死亡率。假设在未开发种群中,不存在任何捕捞死亡率。观测将捕捞死亡率包括进去的影响如下式:
(3)
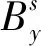
(2)对评估模型的修正
年龄结构模型中,修正的参数如表2,模型中参数多为常数。实际种群分析模型(Virtual population analysis,VPA)和SS3模型尝试将死亡率与环境因子联系起来,考虑种群空间分布的同时考虑海洋环境数据对死亡率、资源补充量等参数的影响,用AIC评估模型。

表2 基于环境因子的年龄结构模型参数统计表
(3)修正模型的应用
探究环境、资源补充量和死亡率之间的关系,环境因子见表2,模型参数考虑生活史中的死亡率,创建一个连续捕鱼死亡率的种群动态。该模型应用物种见表2,该模型模拟的种群动态与真实发展存在较大差异,年龄验证耗时较大,实验数据难以获得,该模型有待继续研究与发展。
2.3 亲体补充量关系
(1)模型构建的理论基础和假设条件
亲体补充量关系是用补充曲线图描述渔业资源群体的资源量与补充量之间函数关系的数学模型。研究亲体补充量是否由产卵场环境变化影响。假设环境因素的影响恒定,不存在高密度时完全消失之点,且补充量随着亲体量的增加而逐渐下降。亲体补充量关系与非平衡剩余产量、年龄结构等模型相结合对资源进行评估。亲体补充量模型应用较多且受环境影响较明显的是Ricker模型[44]:
Ricker的亲体补充量关系(Stock and recruitment relationship, SRR)的表达式如下:
R=αS×e-βs
(4)
式中,α、β为模型中的待定参数;R为补充量;S为亲体量;α为与非密度依赖死亡有关的参数;β是与密度以来死亡有关的参数。
对其补充量R进行优化:
ln(R)-lnS=ln(α)-βS+θV
(5)
式中,V表示环境因子(SST、Pacific Decadal Oscillation PDO);θ为环境因子系数。该模型对环境变化的分析较为显著,加入模型的环境因子见表3,把握环境对渔业资源补充量变化,除成鱼对仔稚鱼的同类同食、密度依赖的其他因素影响和早期生活史中的产卵量的变化外,环境因子也对该模型中的参数造成影响。

表3 基于环境因子的亲体补充量关系模型环境因子参数统计表
(2)对评估模型的修正
Ricker模型参数β依赖于死亡系数,参数α是依赖非密度死亡的参数。而补充量的波动主要还是环境变化导致的。将参数α作为表示不同环境变化的变量,两者常采取分解法求解(多元线性回归),用Akaike信息法则对模型进行比较。亲体补充量关系修正的参数见表3。但在评估过程中有的参数通过模型难以直接得到,需要借助生物量动态模型[45]获取特有参数。
(3)修正模型的应用
该模型强调在亲体量变化水平相当的情况下,环境因子对其资源补充量影响表现较为显著。目前该模型已应用研究见表3评估种类。该模型研究显示加入环境因子的模型对补充量估计效果更好[46]。
2.4 多物种渔业资源评估模型
(1)模型构建的理论基础和假设条件
单物种渔业资源评估过程中,对资源量的自然死亡率指数很难估计。在种间关系的研究进展中揭示资源丰度与自然死亡率变化情况的联系,降低估计自然死亡率系数的不确定性[47]。将适应性系数(Suitability coefficients)等参数设为常数,假设恒定摄食量和单位捕食消耗量与被捕食者资源量无关等[48]。
目前除基于Schaefer模型的单物种评估外,多物种实际种群分析(Multispecies virtual population analysis,MSVPA)[51]是目前较为成功的多物种评估模型之一,对MSVPA进行调整的MSVPA-X模型方法[52],其表达式为:
(6)
Eias=αiaexp(βiats)
(7)
式中,ts表示季节SST;β和α为季节SST实验标准。在评估过程中考虑了物种相互作用,并对其进行敏感性分析。
(2)对评估模型的修正
多物种渔业资源评估模型修正的参数如表4所示,模型拟合过程中考虑生活史中的死亡率参数[51],建立食物有效性和捕食者消费率之间的修正函数关系,将自然死亡率归因于环境等影响,对参数进行敏感性分析。多物种间的复杂关系存在简单假设缺少必要的数据支撑和生物量参考点[53]。

表4 基于环境因子的多物种渔业资源评估模型环境因子参数统计表
(3)修正模型的应用
MSVPA模型方法的扩展,考虑SST依赖性代谢产物,分析开发物种的捕食者-饵料生物之间的相互作用,对模型与参数变化进行敏感性分析。该方法作为从单一物种资源评估到生态系统模型的过渡阶段的关键环节。该模型已评估种类见表4,由于生态系统的复杂性,缺少必要的数据支撑,使其还未成为评估主流模型。
2.5 基于生态系统的渔业资源评估模型
(1)模型构建的理论基础和假设条件
基于生态系统的渔业评估模型中包含复杂的种群生物之间的相互作用机制。引入多个数据,分析环境变化对种群时空动态的影响,建立环境变化与种群时空动态之间的定量关系[55],假设资源量受时空动态影响存在变化。
(2)对评估模型的修正
该模型修正的参数如表5所示,模型中的种群动态与环境联系密切,用多个数据集进行试验,提供预测的范围,评估过程中引入环境要素增加资源预测的不确定性,这些不确定性需降低过程不确定性来抵消[56]。考虑环境变化的动态性和可预测性、资源量对生态系统驱动因素的响应强度和生物资源的生命史[57-58]。资源评估模型中的假设性检验,在评估过程中考虑更全面的生态系统要素。

表5 基于环境因子生态系统的渔业资源评估模型环境因子参数统计表
该模型应用较为成功的模型为空间生态-种群动力学模型(Spatial ecosystem and population dynamics model,SEAPODYM)和基于个体生物模型(Individual-based model,IBM)[59]。利用SEAPODYM对金枪鱼类种群进行研究见表5,耦合生物物理环境,扩展生活史过程中的自然死亡率和生境指数。生态位模型利用物种分布区域所关联的环境因子来模拟物种分布[60]。其中,Elton生态位倾向于较小尺度的生物类环境变量,而Grinnell一般出现在较大尺度的气候变量模型中[61]。例如,巴家文等[61]基于32个环境因子预测了淡水钩虾和湖泊钩虾的适宜分布区域。因此,该模型更多侧重于资源分布地理区域的预测,对于资源评估方面还有待发展。
(3)修正模型的应用
该模型越来越多地被用来区分气候变化和捕鱼对整个生态系统以及对物种种群的影响,SEAPODYM评估模型选择环境时考虑早期生活史幼仔鱼存活率相关系数,分析种群空间动态[63-64]。该模型已评估物种见表5,评估效果最好的是太平洋金枪鱼,该模型现用于运行多物种、多渔场模拟[46],今后研究应倾向于多物种多渔业模拟。
3 讨论与分析
3.1 存在问题
(1)环境因子单一。渔业资源进行种群评估过程中,单一环境与模型中参数结合对种群关系研究受限,不能综合反映环境和物种资源量之间的关系。我国近海鲐鱼、阿根廷滑柔鱼和西北太平洋秋刀鱼在综合产量模型评估中,出现此类问题,环境的滞后作用得不到考虑。模型参数对其早期生活史中死亡率和个体生长等考虑较少[47],年龄结构模型考虑死亡率较多,剩余产量模型中考虑欠佳,降低资源量预测的准确度,忽视海洋环境对不同生长阶段的影响程度。
(2)拟合效果不理想。克服单一的环境,多环境与模型参数进行拟合,出现过度拟合的现象[27]。剩余产量模型和生态系统的评估模型出现拟合效果差,西白令海峡鳕鱼环境综合分析出现数据过度拟合。环境因子选择、模型参数相关的环境因子数据序列较短影响模型的拟合效果。
(3)预测结果不确定性。模型的结构和功能不同使得评估难度增加,模型参数的标准化过程会造成结果不同,增加预测不确定性的风险。环境因子与资源丰度和系统承载能力等相关参数进行研究时,考虑生态驱动要素,增加预测的不确定性。且评估过程中,未进行不同管理方案下资源的长期预测,使得预测结果的不确定性增加[65]。
3.2 今后展望
目前基于环境因子的渔业资源评估模型以单物种模型为主。随着模型的开发,渔业资源评估研究将环境因子纳入不同生态系统模型中,是不断变化环境中适应渔业管理的关键步骤[66],我们对此还具有很大的局限性。环境因子获取完整程度影响模型的选取,因此应研究环境因子选择问题以及环境与参数分析,进一步开发新的模型,保证研究者将选取合适的模型为基于环境因子的渔业资源进行评估。因此环境因子选择时要考虑生活史阶段,考虑环境因子交叉分析,考虑模型中环境因子对模型本身产生影响,对不同海域的海洋环境进行综合分析,对不同时间段的情况进行分段分析[67-68],此外还应考虑生物因素的影响和内部动力过程[69]。
综上,生态系统评估模型逐渐成为渔业评估与管理的主流方向[70]。掌握环境变化对种群生长的影响,是今后进行环境因子的资源评估模型研究的重要内容,也是开发新的生态系统模型的关键。鱼类种群的适应性对未来海洋环境变化保持同步是海洋物种可持续发展的关键环节,管理系统也应适应不断变化的生态系统,提高对渔业生产变化的响应能力和准确性[71]。基于环境因子的资源评估模型是目前对资源可持续开发的一个重要环节[72],生态系统模型在今后渔业资源评估管理研究中将不断深入,基于环境因子的多物种资源评估模型也将不断优化升级。
猜你喜欢
今日农业(2022年15期)2022-09-20
当代水产(2022年7期)2022-09-20
中老年保健(2021年4期)2021-08-22
今日农业(2021年5期)2021-05-22
科学之谜(2020年6期)2020-08-11
海洋信息技术与应用(2020年2期)2020-07-27
当代水产(2019年8期)2019-10-12
今日农业(2019年14期)2019-09-18
当代水产(2019年3期)2019-05-14
红土地(2018年7期)2018-09-26
