
小麦SUS 基因家族鉴定与生物信息学分析*
2024-04-01 14:12孔斌雪马靖福窦佳欣张沛沛杨德龙
云南农业大学学报(自然科学) 2024年1期
孔斌雪,李 娜,马靖福,窦佳欣,陈 涛,张沛沛,刘 媛,杨德龙,**
(1.甘肃农业大学 生命科学技术学院,甘肃 兰州 730070;2.省部共建干旱生境作物学国家重点实验室,甘肃 兰州 730070)
蔗糖是光合作用的产物,其作为最初的能量供体与植物生长发育的各种代谢途径相关,对植物生长发育至关重要[1-2]。蔗糖参与调节细胞分裂、种子萌发、开花诱导、花青素合成、果实发育等一系列生理代谢过程[3]。蔗糖代谢研究对于了解植物生理具有重要意义。蔗糖合成酶(sucrose synthase,SUS)是植物体内调节蔗糖代谢的重要酶之一,在蔗糖运输到库器官之前将其分解[4],首先在小麦胚芽中被发现[5]。SUS 在植物代谢过程中具有关键作用,其活性与库强度、生物量积累以及碳分配相关[2]。研究发现:SUS 活性可影响水稻籽粒大小及产量[6];玉米sh1突变体和豌豆rug4突变体胚乳的SUS 活性与野生型相比均有所下降,表现出籽粒干瘪和粒质量下降[7-8];拟南芥SUS基因突变体叶片和茎中的SUS 活性与野生型相比下降15%[9];SUS 受到RNAi 抑制后,马铃薯块茎中的淀粉含量降低,导致干质量下降,番茄中的蔗糖卸载能力也降低,座果数下降[10-11]。除参与广泛的代谢过程外,SUS 还与非生物胁迫响应相关。研究发现:SUS基因的转录水平随着氧气含量的降低而增强[12-14]。SUS 也在热应激条件下植物的生理代谢中发挥作用[15-16]。
在高等植物中,SUS 由1 个小的多基因家族编码。前人对多种植物的SUS基因家族成员进行了鉴定和研究,结果显示:拟南芥(Arabidopsis thaliana)[17]、水稻(Oryza sativa)[13]、烟草(Nicotiana sylvestris)[18]和巴西橡胶树(Hevea brasiliensis)[19]有6 个SUS基因家族成员,石榴(Punica granatum)[20]和葡萄(Vitis vinifera)[21]有5 个SUS基因家族成员,亚洲棉(Gossypium arboreum)有7 个SUS基因家族成员[22],杨树(Populus trichocarpa)有15 个SUS基因家族成员[23]。前人对SUS基因家族的研究发现:SUS基因遵循基因家族进化,表现为结构保守但功能分化[24]。
小麦(Triticum aestivumL.)是全球最重要的粮食作物之一[25],其产量受灌浆速率和灌浆持续期的影响[26]。小麦籽粒灌浆是蔗糖等光合产物向库器官运输的过程[27],是决定粒质量、产量和品质的重要时期。目前,仅有少数小麦SUS基因被鉴定,TaSUS1、TaSUS2和TaSUS3在发育的籽粒中表达,在整个小麦主要灌浆期均检测到TaSUS的转录本[28-29]。研究还发现:水稻籽粒胚乳的SUS活性决定水稻籽粒库强,对水稻籽粒灌浆过程起调节作用[30],这表明小麦SUS基因家族在调节籽粒灌浆和产量方面可能起关键作用。本研究利用小麦全基因组序列信息鉴定小麦SUS基因家族成员,并对其生物信息学特征进行分析,为进一步研究小麦SUS基因家族调节植物生长发育,特别是在籽粒发育中的功能提供理论基础。
1 材料与方法
1.1 小麦SUS 基因家族成员的鉴定与系统发育分析
从Ensemble Plants 数据库[31]下载小麦全基因组数据、蛋白质序列和注释文件,鉴定小麦SUS基因家族的成员。从PFAM 数据库[31]下载SUS保守结构域HMMER 文件(PF00862),并将其作为查询序列用于搜索小麦蛋白数据库中的SUS基因家族成员(阈值≤1×10-10)。为了保证数据的准确性,进一步通过NCBI-CDD、Inter Pro 和SMART 数据库[32-34]验证其是否具有SUS 结构域。根据上述结果,剔除重复基因和不含SUS 结构域的基因,然后使用MEGA 软件[35]对拟南芥、水稻和小麦SUS基因家族蛋白序列进行多重序列比对,构建系统发育树,并在EvolView[36]网站美化系统进化发育树。
1.2 小麦SUS 基因的染色体定位与序列分析
利用Ensembl Plants 数据库获得TaSUS基因在染色体上的位置,利用TBtools 软件进行定位。利用 MEME Suite 在线工具[37]分析TaSUS基因家族成员的保守结构域,参数motif 数量设置为10,使用 TBtools 软件绘制基因结构图和motif 图。利用TBtools 软件提取TaSUS基因编码区上游2 000 bp序列,并提交到PlantCARE 数据库[38]确定顺式作用元件的种类和数量。通过ExPASY 在线工具[39]预测最终得到的TaSUS基因家族成员的等电点和分子质量。亚细胞定位通过Gpos-mPLoc 在线工具[40]进行预测。
1.3 小麦SUS 基因复制及共线性分析
片段复制和串联重复对基因家族扩张非常重要[41],为分析不同物种间SUS基因家族的共线性,从Ensemble Plants 数据库[31]下载不同物种的基因组信息,并采用TBtools 中的One Step MCScanX 模块进行共线性分析以及可视化。
1.4 小麦SUS 基因的表达模式分析
从小麦表达数据库expVIP[42]下载小麦品种中国春的表达数据,分析TaSUS基因在其籽粒、根、茎、叶和穗5 个不同组织的转录水平。基因表达量热图利用TBtools 软件绘制。
2 结果与分析
2.1 TaSUS 基因家族成员的鉴定与命名
从小麦全基因组中鉴定到24 个SUS基因(表1),命名为TaSUS1~TaSUS24。小麦SUS 蛋白质氨基酸数量在560~1 074 之间,分子质量为63.61~118.27 ku,理论等电点为5.69~8.69,除Ta-SUS2、TaSUS4、TaSUS6、TaSUS19、TaSUS21和TaSUS24 外,其他TaSUS 蛋白质的等电点都小于7,呈酸性。亚细胞定位预测分析表明:除TaSUS17 和TaSUS22 位于细胞质和细胞膜外,其他TaSUS 蛋白均位于细胞质。TaSUS基因家族成员均含有SUS 结构域。

表1 小麦SUS 基因家族的理化性质Tab.1 Physicochemical properties of SUS gene family in wheat
2.2 TaSUS 基因的系统进化
由TaSUS基因与6 个拟南芥AtSUS基因、6 个水稻OsSUS基因的蛋白序列构成的系统发育树(图1)可知:SUS基因家族可被分为3 个亚家族。其中,Ⅰ类基因20 个,包括拟南芥基因4 个、水稻基因4 个和小麦基因12 个;Ⅱ 类基因13 个,包括拟南芥基因2 个、水稻基因2 个和小麦基因9 个;Ⅲ 类基因只包含3 个小麦基因。部分小麦SUS基因家族成员与水稻的遗传关系比其与拟南芥更密切。
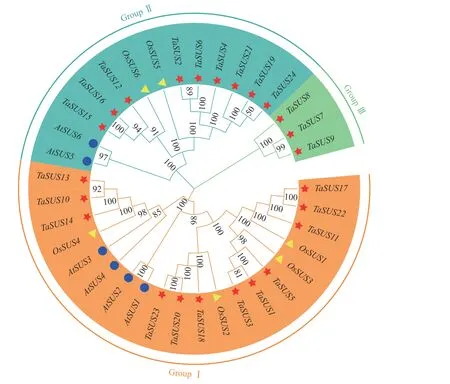
图1 小麦、拟南芥和水稻SUS 基因家族的系统进化树Fig.1 Phylogenetic tree of SUS gene family in wheat,Arabidopsis and rice
2.3 TaSUS 蛋白保守结构域及基因结构
由图2 可知:每个TaSUS 蛋白序列都包含多个(4~10 个)保守基序(motif),其中motif1、mo-tif2、motif3 和motif8 保守分布于TaSUS 蛋白序列;同属于第Ⅲ亚组的TaSUS7、TaSUS8 和TaSUS9 的motif 较为特异,motif 长且分散。这表明同一家族中的小麦TaSUS 蛋白高度保守,并且在进化过程中,不同亚群中的成员可能发生了功能分化。基因结构分析显示:除TaSUS12基因只含有9 个外显子外,其余基因外显子数量为12~17个;18 个TaSUS基因具有完整的非翻译区(untranslated region,UTR),但TaSUS1和TaSUS12基因的3′和5′端均没有UTR,TaSUS17、TaSUS20、TaSUS22和TaSUS23的3′端不含UTR;同一亚组的TaSUS基因结构分布相对保守。

图2 小麦SUS 基因家族的蛋白保守基序和基因结构Fig.2 Protein conserved motif and gene structure of SUS gene family in wheat
2.4 TaSUS 基因的染色体定位和共线性
染色体定位结果显示:除1、5 号染色体外,TaSUS基因在其他染色体均有分布,其中,4A、7A 和7D 染色体上各有3 个TaSUS基因,3、6 号染色体上仅各有1 个TaSUS基因;共线性分析鉴定出20 对片段重复基因,且7 号染色体上的片段重复基因占比最多(8 个),每一对片段重复基因都位于相同的TaSUS亚组,且小麦SUS基因家族中不存在串联重复基因 (图3a)。物种间水稻、拟南芥和小麦SUS基因的共线性分析结果(图3b)显示:1 对TaSUS基因与AtSUS基因具有共线性,16 对TaSUS基因与OsSUS基因具有共线性,说明小麦SUS基因家族与水稻SUS基因家族亲缘关系较近,可能具有相似的生物学功能。

图3 SUS 基因家族在小麦染色体上的定位(a)以及物种间的共线性(b)Fig.3 Location on the chromosomes of wheat (a) and synteny among species (b) of SUS genes family
2.5 TaSUS 基因启动子区的顺式作用元件
TaSUS基因的启动子区共鉴定出45 种顺式作用元件,可分为激素响应元件(hormone responsive elements,HRE)、光响应元件(light responsive elements,LRE)、胁迫响应元件(stress responsive elements,SRE)、组织特异性元件(tissue specific elements,TSE)和其他响应元件(other responsive elements,ORE)(图4)。植物激素、光响应和胁迫响应相关的元件存在于所有TaSUS基因的启动子区,其中,激素相关的顺式作用元件有生长素(auxin,IAA)、水杨酸(salicylic acid,SA)、脱落酸(abscisic acid,ABA)、茉莉酸甲酯(methyl jasmonate,MeJA)和赤霉素(gibberellin,GA)。在小麦SUS基因家族成员中,有80%的成员都包含ABA 相关的作用元件ABRE 以及MeJA 相关的作用元件CGTCA-motif 和TGACGmotif;LRE 在24 个TaSUS基因启动子中普遍存在;每个TaSUS基因启动子都包含1~8 个SRE,对低温、缺氧、干旱等胁迫做出响应;在TSE中,有RY-element、CAT-box 和GCN4_motif 顺式作用元件,分别与种子、分生组织和胚乳发育相关;TaSUS基因启动子中还包含ORE,如细胞周期调节、昼夜节律调控、玉米醇溶蛋白代谢调节和MYB 结合位点。说明TaSUS基因在调节植物生长发育中发挥着多种作用。
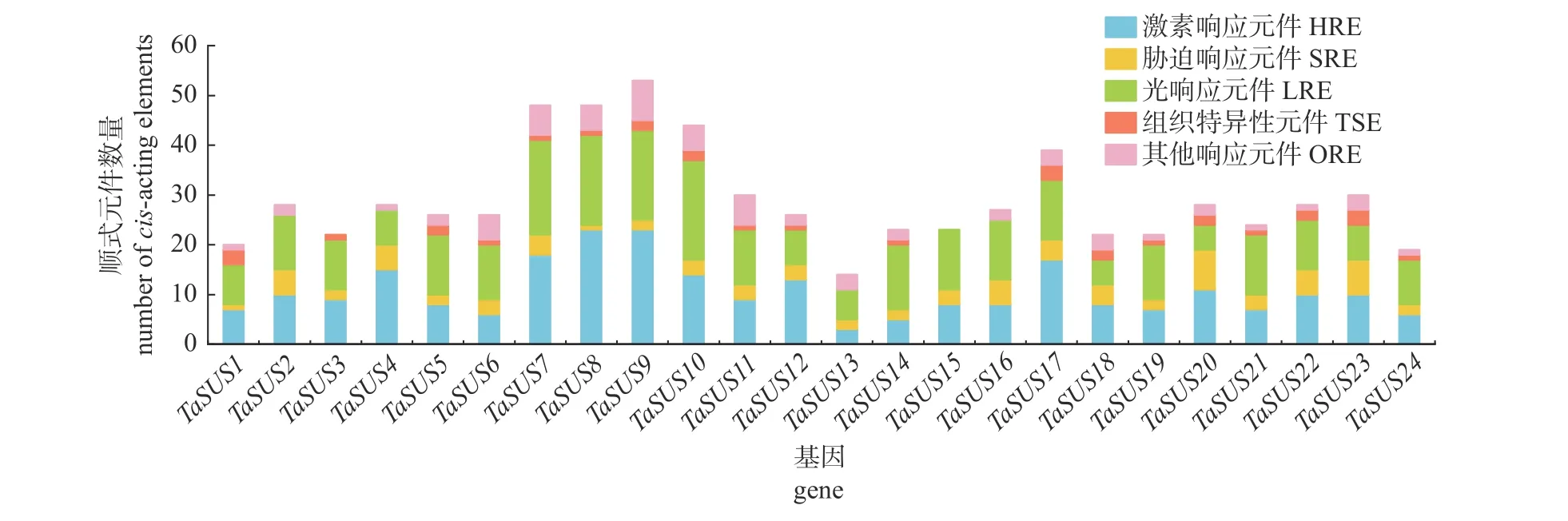
图4 TaSUS 基因中的顺式作用元件Fig.4 Cis-acting elements in TaSUS genes
2.6 TaSUS 基因的表达模式
由图5 可知:大多数TaSUS基因在小麦穗中显著表达,在叶、茎和根中的相对表达量较低,Ta-SUS3、TaSUS5和TaSUS6在小麦籽粒中显著表达。
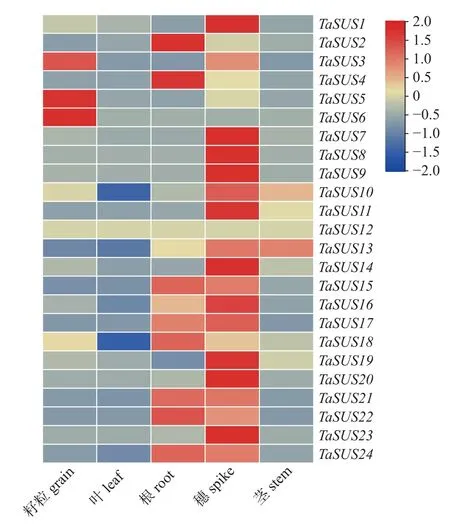
图5 小麦 SUS 基因在不同组织中的表达量Fig.5 Expression levels of SUS gene in different tissues of wheat
3 讨论
SUS基因家族对植物的生长发育至关重要,但鲜有小麦中SUS基因的鉴定与功能研究。本研究共鉴定到24 个TaSUS成员,明显多于其他植物,其原因可能是小麦作为六倍体物种,基因组庞大,同源基因较多。前人研究表明:片段复制和串联重复是植物中常见的基因复制事件,在基因家族扩展过程中扮演重要角色[43]。在鉴定到的24 个TaSUS基因中,有20 对基因存在片段复制,说明在小麦SUS基因家族扩增过程中重复事件发挥了重要作用,分析其原因可能是小麦SUS基因家族成员较多。系统发育树分析结果表明:36 条 SUS 蛋白序列分为Ⅰ、Ⅱ 和Ⅲ组,这与其他植物中SUS的报道[2-3,17]一致,且Ⅰ组中的基因数量最多,Ⅲ组中不包含OsSUS和AtSUS基因,表明小麦中亚家族Ⅰ比其他基因组扩展得多。
SUS 蛋白一般是平均分子质量约为90 ku(约800 个氨基酸)的同源四聚体[2],如水稻OsSUS1-6[13]、拟南芥AtSUS1-5[17]和石榴PsSUS1-4[20],但其他SUS 异构体的分子质量则不同,如葡萄VvSS5 分子质量为102.7 ku[21],拟南芥AtSUS6分子质量为106.86 ku[17],甘蔗SoSuSy4 分子质量为106.81 ku[3]。本研究表明:大多数小麦SUS 蛋白分子质量约为90 ku,且都含有酸性氨基酸,有少数蛋白的分子质量大于100 ku,其物理化学性质与其他植物相似。已有研究表明:SUS Ⅰ和SUS Ⅱ基因有15 个外显子,而SUS Ⅲ基因因存在3′端延伸有17 个外显子[24]。本研究发现:小麦SUS家族基因的外显子为 9~17 个,表明小麦进化过程中SUS基因家族外显子存在增加或缺失。
基因表达模式分析可在一定程度上用于预测基因参与的生理过程[21,44-45]。本研究发现:大多数TaSUS基因在小麦穗中显著表达,TaSUS3、Ta-SUS5和TaSUS6在小麦籽粒中显著表达,表明TaSUS基因可能在调节小麦生长发育过程中发挥重要作用;TaSUS12基因虽未在小麦品种中国春的组织中表达,但其可能在其他小麦品种生长发育过程中发挥作用。SUS基因对果实发育的调节作用已在其他植物中得到证实,如:VvSS3在营养组织浆果中高表达,对调节浆果中的糖积累起关键作用[21];在17 个PbrSUS基因中,有8 个在梨幼果中的表达量较高,且其表达量随着果实的成熟逐渐降低[46];MdSUS1.1/1.2主要在幼果和成熟果实中表达,在果实发育过程中MdSUS2.1的表达量达到最高[47];AtSUS2在拟南芥的成熟种子中特异性表达[17]。本研究表明:TaSUS基因在小麦穗和籽粒中的表达水平高于叶、茎和根,说明SUS可能参与调节籽粒发育过程。
4 结论
本研究共鉴定到24 个TaSUS基因,可分为3 个不同的亚组。同一家族中的小麦TaSUS 蛋白高度保守,推测TaSUS基因可能参与小麦的生长发育和逆境胁迫响应。下一步研究可致力于对TaSUS基因的过表达和抑制分析,确定TaSUS在籽粒发育中的特定功能。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
科学与财富(2021年3期)2021-03-08
——以多重共线性内容为例
长沙航空职业技术学院学报(2019年2期)2019-07-13
温州大学学报(自然科学版)(2019年2期)2019-06-04
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
现代工业经济和信息化(2016年2期)2016-05-17
电子工业专用设备(2015年4期)2015-05-26
汽车维修与保养(2015年8期)2015-04-17
