
基于GYT双标图分析对黄淮海生态区玉米品种综合评价
2024-03-28 05:46:18岳海旺魏建伟刘朋程陈淑萍卜俊周
作物学报 2024年4期
岳海旺 魏建伟 刘朋程 陈淑萍 卜俊周
基于GYT双标图分析对黄淮海生态区玉米品种综合评价
岳海旺 魏建伟 刘朋程 陈淑萍 卜俊周*
河北省农林科学院旱作农业研究所 / 河北省农作物抗旱研究重点实验室, 河北衡水 053000
针对不同环境、多性状条件下优良品种选择效率低下的问题, 探讨整合环境型鉴定技术(envirotyping techniques, ET)和多性状选择对黄淮海夏玉米区试参试品种进行综合评价, 以期为品种合理布局提供理论依据。本研究以2016—2017年黄淮海夏玉米组区域试验数据为材料, 基于当年19个环境协变量信息采用ET将40个试点划分为不同生态区(mega-environments, ME)。采用品种-产量×性状(genotype by yield × trait, GYT)双标图技术对不同生态区(mega-environments, ME)籽粒产量与生育期、株高、穗位高、倒伏率、空秆率、穗长、秃尖、穗行数、穗粒重、百粒重、茎腐病和黑粉病等农艺性状的组合表现进行综合评价, 研究GYT双标图技术在玉米区域试验多性状评价中的作用。AMMI方差分析表明, 2016年被测农艺性状基因型、环境和互作效应均达到了极显著水平(<0.01), 2017年被测农艺性状除穗位高互作效应不显著外, 其余性状基因型、环境和互作效应均达到了极显著水平。根据当年气象因子信息将位于8个省份的40个试点划分为4个ME, 降水亏缺(dbp)、饱和水汽压差(vpd)、相对湿度(rh)和最高温度(max)在5个物候期中呈现出较大的变化趋势。GYT双标图与ME结合, 可以筛选出不同ME的优势品种。2016年参试品种中, 衡玉321和冀丰118在划定的4个ME中均表现出丰产性突出、稳定性较好的特征, 属于丰产稳产型品种。而潞玉36和潞研1502则属于参试品种中丰产性、稳定性均较差的品种。2017年参试品种中, DK56在ME2和ME4试点中产量-性状组合表现较为协调, DK205和衡玉6105分别在ME1和ME3生态区中有较好的表现。对照品种郑单958两年区域试验表现出较好的稳定性但丰产性一般。基于环境型鉴定技术划分生态区与GYT双标图相结合对参试品种的丰产性、适应性和稳定性进行评价, 实现品种推广的精细定位, 为黄淮海夏玉米区品种多性状综合评价提供理论基础。
夏玉米品种; 生态区; 基因型与环境互作; 气候变量; GYT双标图
玉米(L.)在总产方面已经超过了水稻(L.)和小麦(L.), 是世界上最重要的谷物作物之一[1]。我国是全球第二大玉米生产国, 依托于科技创新, 我国玉米单产已由21世纪初的4700 kg hm–2提高至2020年的6310 kg hm–2, 玉米育种对单产的贡献率超过了45%[2]。玉米产量是由多基因控制的复杂数量性状, 由基因型(genotype, G)、环境(environment, E)以及基因型与环境互作(genotype by environment interaction, GEI)等方面共同影响。品种的GEI效应越大, 说明对环境越敏感, 稳定性越差[3-4]。黄淮海夏玉米区是我国玉米三大主产区之一, 也是夏玉米最大集中种植区。该区气象条件复杂多变, 经常遇到高温热害、干旱缺水、光照不足、阴雨连绵, 以及各种病虫害的侵袭, 造成每年产量差异较大[5-8]。因此, 迫切需要更好的了解黄淮海地区基因型与环境互作效应(GEI), 以此选育出具有特殊气候适应性的优良玉米品种, 通过多年多点鉴定试验进行全面评估, 这是应对气候变化背景下提高黄淮海地区夏玉米产量的有效方法。以往有关环境因子影响作物生产的研究主要是通过作物在不同环境中的表现型来预估其综合作用,或对整个生态区的部分环境因子来对比分析, 无法对表现型进行有效预测, 最终无法对各类环境因子开展深层次解析, 成为制约作物生产和作物改良的重要瓶颈。中国农业科学院作物科学研究所徐云碧博士首次在国际上提出环境型鉴定技术(envirotyping techniques, ET), 其定义为对所有环境因子的组成和变异的精确解析和评价。环境型鉴定技术主要应用于环境特征分析、基因型与环境互作分析、表现型预测、精准农业以及精准育种等。该技术为作物育种在内的作物科学提供了解码环境影响的参考技术和途径, 具有广阔的应用前景[9-12]。加性主效应乘积交互作用(additive main effects and multiplicative interaction, AMMI)模型已经广泛用于作物基因型与环境互作分析, 该模型整合方差分析(analysis of variance, ANOVA)和主成分分析(principal component analysis, PCA), 对交互作用主成分轴分解为若干个乘积项, 以提高预测的准确性[13]。随着玉米审定渠道多样化, 对玉米品种精细定位, 确定最佳的推广区域, 充分发挥品种特征优势, 玉米生态区(mega-environments, ME)的划定显得尤为重要[14]。为了便于不同品种多性状间的比较和对比, 品种-性状(genotype by trait, GT)双标图已被广泛用于作物品种多性状的分类评价、形象直观显示目标农艺性状间的相关性, 但在对品种多性状综合评价方面存在不足。同时, 对于目标性状的评价都是建立在丰产性的基础上。针对品种-性状(genotype by trait, GT)双标图在农作物多性状综合评价方面存在的缺陷和应用的局限性, 严威凯博士提出了GYT (genotype by yield × trait biplot, GYT)双标图的概念, GYT双标图可以利用产量和目标性状组合对农作物多种被测农艺性状进行品种分类和综合评价, 具有针对性更强和适用性更广等特点[15-17]。目前环境因子对玉米生育期内产量影响的报道较多, 但基于不同环境因子的玉米生态区划分以及不同生态区内GYT双标图对玉米品种产量-性状组合的综合评价的文章尚未见于报道。本研究的主要目的是利用环境类型学技术在黄淮海地区划定不同生态区, 并在每个生态区中选择基于多性状的、跨年度稳定的优良杂交品种。本研究为更系统和动态地描述试验环境提供了新的见解, 有助于育种者在玉米育种项目中做出更好的战略决策, 实现有效的多元选择。
1 材料与方法
1.1 参试品种和试验点
参试品种包含22个杂交种, 郑单958为对照品种。试验于2016—2017年在黄淮海夏播区的河北、山东、河南、安徽、山西、江苏、湖北和陕西等8个省40个试点进行, 参试品种名称和各试点的经纬度等信息分别见表1和表2。
1.2 试验设计
采用完全随机区组设计, 重复3次。每个小区行长6.7 m, 行距0.6 m, 5个行区, 小区面积20.1 m2。播种期为5月底到6月上旬, 种植密度75,000株 hm–2。收获期为9月底到10月上旬, 试验期间各项农艺措施参照当地大田管理。2016—2017年测定的农艺性状一致包括籽粒产量(kg hm–2)、生育期(d)、株高(cm)、穗位高(cm)、穗长(cm)、秃尖长度(cm)、百粒重(g)、倒伏率(%)、空秆率(%)、穗粒重(g)、穗行数、茎腐病(%)和黑粉病(%)。各性状测定按照《农作物品种试验技术操作规程玉米》[18]要求执行。
1.3 生态区划分
本研究使用R语言软件包EnvRtype[19]中的get_wheater()函数来收集每个试点以日为单位的19个环境协变量(environmental covariables, EC) (表3)。每个试点观测到的19个协变量(environmental covariables, EC)被用来创建称为环境型(envirotype)协变量矩阵, 按照Costa-Neto的方法[19], 使用EnvRtype包的W_matrix() 函数进一步用来计算环境亲缘关系。
为了更好地捕捉环境信息在整个作物生长过程中的时间变化, 根据玉米不同的物候期划分, 作物周期按播种后天数(days after sowing, DAS)分为5个主要物候期, 0~14 d (初始生长期); 15~35 d (叶片扩展期I, V4—V8); 36~65 d (叶片扩展期II, V8—VT); 66~90 d (开花期)和91~120 d (灌浆结实期)。对于每个变量-阶段组合, 我们计算了第一四分位数(25%)、中位数(50%)和第三四分位数(75%)。因此, 环境协变量×物候期×四分位数(19×5×3=285)的285个组合中的每一个都成为环境相关度的一个环境类型描述符。然后, 利用(20行×285列)矩阵, 我们使用EnvRtype软件包的env_kernel()函数, 计算出一个环境内核(enviromic kernel), 如下所示:

式中,E是基于相似环境间的内核,是环境类型描述符矩阵。为了识别生态区, 对E采用了分层聚类(平均法)。

表1 参试品种信息

表2 试点环境信息

表3 本研究中的19种环境协变量
a代表从NASA直接获得的气象数据[20],b代表根据Ghanem等[21]计算过的数据。
arepresents the meteorological data obtained directly from NASA orbital sensors[20];brepresents the data were calculated according to Ghanem et al.[21]
最后, 为了直观地了解环境变量之间的关系以及它们与平均表现和稳定性的关系, 我们用一个包含每个环境/季节的环境变量和籽粒产量的双向表进行了主成分分析。用R软件包factoextra中的fviz_pca_biplot()函数制作双标图, 用来显示研究性状之间的关系。
1.4 AMMI模型分析
AMMI模型整合了标准的方差分析和主成分分析(principal component axis, PCA), 以确定交互主成分(IPC)来计算稳定性参数, AMMI模型可以用以下公式来概括:

式中,Y是第个基因型在第个环境中的目标性状产量;代表总体平均值;α代表第个基因型效应;β代表第个环境效应;λ代表第个主成分轴(PCA)的奇异值;δ和γ分别为基因型、环境和组分的特征向量值;ρ为残差;为主成分轴(PCA)的数量[22]。
1.5 GGE双标图
GGE双标图模型是基于双标图技术引入的, 它是可视化双向数据的有效工具, 经常用于农作物多点测试(Multi-environment trials, METs)的数据分析[23-24]。GGE双标图能够同时显示双向数据表中的基因型主效应(G)和基因型×环境效应(GE)。它的第1个组成部分, 当与基因型主效应(G)高度相关时, 代表仅归因于基因型的解释比例。第2个代表基因型与环境互作(GEI)解释的比例。生成GGE双标图均值矩阵必须以环境为中心, 然后通过奇异值分解(singular values decomposition SVD)为主成分, 然后使用前2个主成分得分(PC1和PC2)生成图形。GGE双标图模型如下:
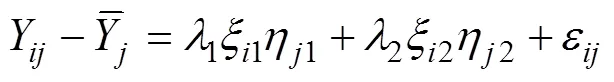

1.6 GT双标图
将2016—2017年参试的22个品种在40个试点的被测农艺性状, 籽粒产量(kg hm–2)、生育期(d)、株高(cm)、穗位高(cm)、穗长(cm)、秃尖长度(cm)、百粒重(g)、倒伏率(%)、空秆率(%)、穗粒重(g)、穗行数、茎腐病(%)和黑粉病(%)组建品种×性状(genotype by trait, GT) 双标图。GT双标图通过将每个品种和性状的前2个主成分(PC1和PC2)来构建的,模型公式如下:

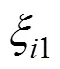
1.7 GYT双标图的构建
参考Yan和Frégeau-Reid的方法[15]进行GYT双标图的构建。首先, 穗长(ear length, EL)、百粒重(hundred-seed weight, HSW)、穗粒重(grains weight per ear, GWE)和穗行数(Kernel row number, KRN)等农艺性状, 产量-性状组合数值是通过将产量与每个性状相乘(例如, Y×EL, Y×HSW, Y×GWE, Y×KRN), 对于以上产量-性状组合数值是越大越好。而对于茎腐病(stalk rot, SR)、黑粉病(common smut, CS)、倒伏率(lodging rate, LR)和秃尖(bare tip length, BTL)等农艺性状的产量-性状组合值是越小越好, 表达方式为在产量-性状组合后面加后缀(–1), 比如Y×SR (–1)等。其次, 是对GYT表格进行标准化数据处理。每个品种的标准化产量-性状组合平均值为该品种的理想指数(superiority index, SI), 可以用来对品种排序。标准化处理公式如下:

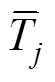
最后, GYT双标图的形成与GT双标图过程一致,只是把GT双标图中的“性状”更改为“产量-性状组合”即可。
1.8 统计分析
Microsoft Excel 2020用于数据录入和整理, DPS V19.05版本用于AMMI模型分析[27], R software version 4.2.3软件进行生态区划分, GT双标图和GYT双标图分析通过GGEbiplot软件完成。
2 结果与分析
2.1 AMMI模型分析
对2016—2017年参试品种测定的13个农艺性状进行AMMI模型分析, 结果表明, 被测农艺性状变异来源中, 基因型(G)和环境(E)效应均达到了极显著水平(<0.01)。其中, 2016年除穗粒重、秃尖长度和百粒重等性状基因型与环境互作效应达到了显著水平(<0.05)外, 其余性状互作效应均达到了极显著水平。籽粒产量、百粒重、生育期、穗位高、空秆率、穗粒重和穗长等性状环境效应占比最高, 分别为55.31%、46.53%、44.44%、50.64%、66.19%和37.24%, 是变异的主要来源。秃尖长度、倒伏率、穗行数、茎腐病和黑粉病等性状互作效应在总平方和中占比最高, 分别为40.88%、60.77%、38.03%、76.78%和52.78%。对交互效应进行分解为第1主成分轴(PC1)和第2主成分轴(PC2), 显著性检测表明13个性状的PC1和PC2均达到了极显著水平, 前两项互作主成分占交互效应平方和百分比最大的是茎腐病(87.50%), 最小的是穗行数(38.14%) (表4)。2017年除穗位高等性状外, 其余12个性状互作效应均达到了极显著水平。籽粒产量、生育期、穗长、百粒重、空秆率和穗粒重等性状在总平方和占比方面趋势一致, 环境效应占比最大, 分别为64.94%、85.46%、42.23%、55.71%、78.75%和66.94%。茎腐病、秃尖长度、倒伏率、穗行数和黑粉病的互作效应平方和占比最大, 分别为47.22%、60.60%、64.41%、91.41%和64.93%。13个被测农艺性状PC1和PC2均达到了极显著水平, 穗行数和百粒重分别是PC1和PC2占比交互效应比例最高(98.36%)和最低(32.92%) (表5)。通过2年AMMI模型分析, 大部分被测性状互作效应在总平方和中占比要大于基因型效应, 要重视互作效应在品种选育过程中的作用, 为品种的合理布点提供科学依据。
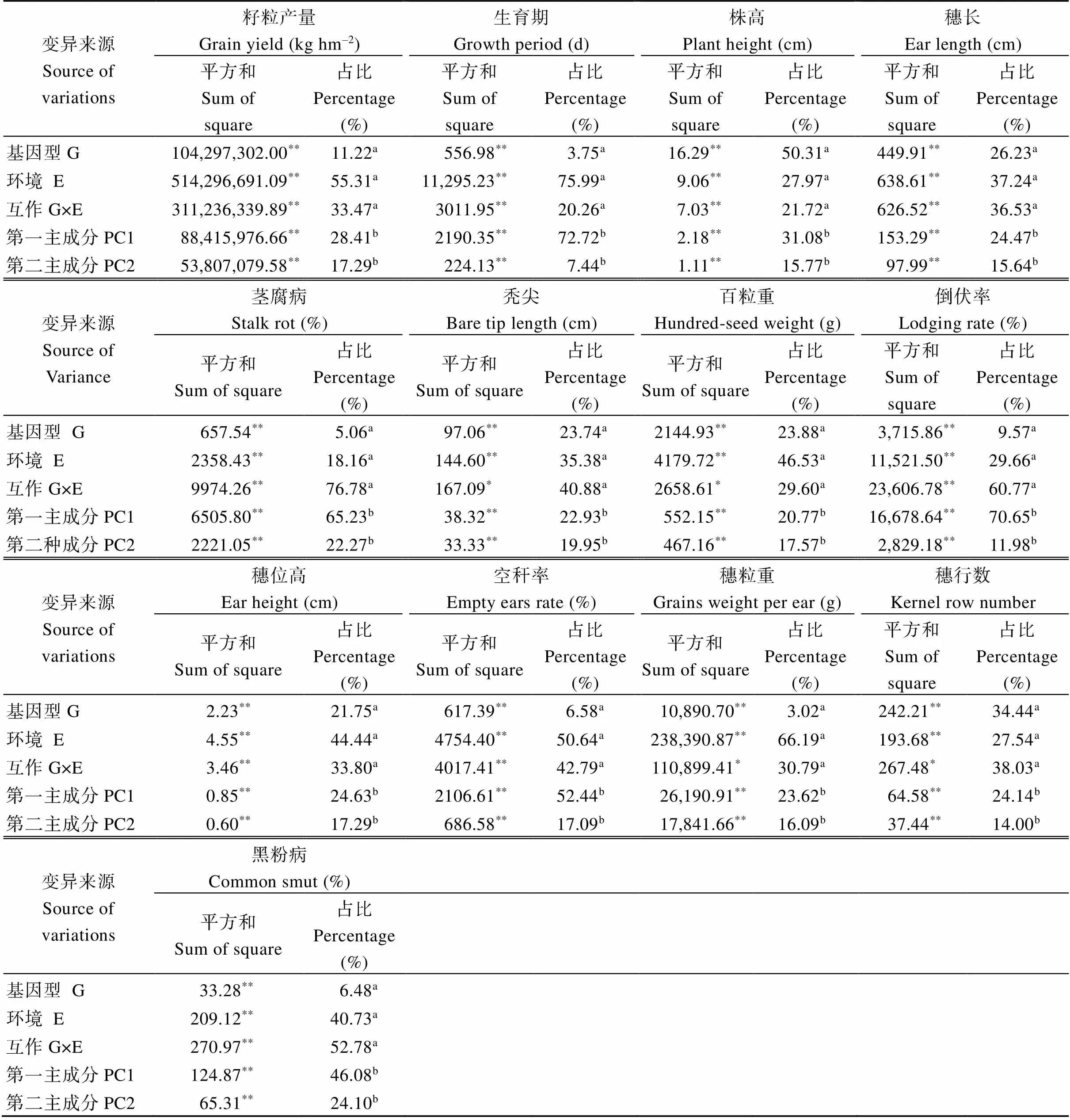
表4 2016年参试品种农艺性状AMMI模型分析
*和**分别表示在5%和1%概率水平差异显著。G和E分别代表基因型因素和环境因素。a表示的是总平方和占比,b表示的是互作效应占比。
*and**indicate significant difference at the 5% and 1% probability levels, respectively. G and E represent genotype and environment, respectively.aandbindicate percentage of total sum of square and sum of square of GE, respectively.

表5 2017年参试品种农艺性状AMMI模型分析
*和**分别表示在5%和1%概率水平差异显著,ns表示不显著。a表示的是总平方和占比,b表示的是互作效应占比。
*and**indicate significant difference at the 5% and 1% probability levels, respectively.ns: not significant.aandbindicate percentage of total sum of square and sum of square of GE, respectively.
2.2 生态区划分
2016和2017两年根据285个环境协变量的信息将位于8个省份的40个试点划定了4个生态区(ME)。2016年所有山东试点章丘、潍坊、平度、临沂、临朐、聊城、莱州、济宁、嘉祥、菏泽和德州, 河南试点中的巩义、荥阳、鹤壁、滑县、焦作、洛阳、新乡、小冀和原阳, 河北试点中的邯郸等22个试点属于第1生态区(ME1)。河南省南部的驻马店、周口、商丘和南阳, 安徽的泗县、埇桥、界首, 江苏的宿迁、徐州等9个试点属于第2生态区(ME2)。山西的运城、陕西的杨凌和泾阳等3个试点属于第3生态区(ME3)。剩余河北的高阳、深州、泊头、武强、藁城和赵县等6个试点属于第4生态区(ME4) (图1-A)。2017年所有河南试点包括原阳、西平、新乡、荥阳、西华、南阳、洛阳、临颍、焦作、辉县、滑县、鹤壁和长葛, 安徽埇桥、界首, 江苏徐州和湖北襄阳等17个试点划归为第1生态区(ME1), 所有山东试点包括淄博、潍坊、泰安、平度、临沂、临朐、聊城、莱州、济宁、嘉祥、东营、德州和茌平, 河北邯郸等14个试点划归为第2生态区(ME2), 河北的赵县、永年、行唐、武强、深州和藁城等6个试点划归为第3生态区(ME3), 陕西的杨凌和泾阳, 山西的运城等3个试点划归为第4生态区(ME4) (图1-B)。
2.3 2年环境因子变化
各气候因子在2年中呈现较大的变化趋势。2016年第1生态区(ME1)降水亏缺(dbp)最大值和最小值分别为126.02mm d–1和–13.97 mm d–1, 潜在蒸散量(etp)为1.22~13.97mm d–1, 相对湿度(rh) 32.62%~95.50%, 水平表面上的日射(sihs)、最高温度(max)、平均温度(mean)、最低温度(min)和饱和水汽压差(vpd)的最高、最低区间分别为30.18~2.70 MJ m–2d–1、39.7~17.8℃、32.5~16.4℃、27.6~10.7℃和3.28~0.18 kPa d–1。第2生态区(ME2)平均温度(mean)范围为17.8~32.3℃, 最高温度(max)范围为20.8~ 38.8℃, 最低温度(min)范围为13.8~28.7℃, 降水亏缺(dbp)在–13.71~95.57 mm d–1, 潜在蒸散量(etp)区间范围在1.65~13.71mm d–1, 饱和水汽压差(vpd)区间范围在0.25~3.03kPa d–1, 水平表面上的日射(sihs)最大和最小值分别为29.63 MJ m–2d–1和3.80 MJ m–2d–1。相对湿度(rh)最大和最小值分别为94.12%和36.38%。第3生态区(ME3)最大降水亏缺(dbp)和最大相对湿度(rh)是4个生态区中最小的, 分别为43.35 mm d–1和90.12%, 潜在蒸散量(etp)范围为1.72~14.22 mm d–1,平均温度(mean)区间范围在16.55~32.76℃, 饱和水汽压差(vpd)为0.16~3.07 kPa d–1。第4生态区(ME4)降水亏缺(dbp)和潜在蒸散量(etp)分别为–14.20~ 112.01 mm d–1和1.25~14.20 mm d–1, 最小相对湿度(rh)是4个生态区中最小的为26.38%, 平均温度(mean)为15.2~31.9℃ (附图1-A)。2017年最低相对湿度(rh)和最高相对湿度均出现在第1生态区(ME1)分别为34.50%和93.94%, 降水亏缺(dbp)最高值出现在第2生态区(ME2)为73.55 mm d–1, 最高温度(max)、平均温度(mean)、最低温度(min)的最低值均出现在第3生态区(ME3)分别为11.16、9.33和6.71℃,平均温度(mean)的最高值出现在第4生态区(ME4)为34.99℃ (附图1-B)。
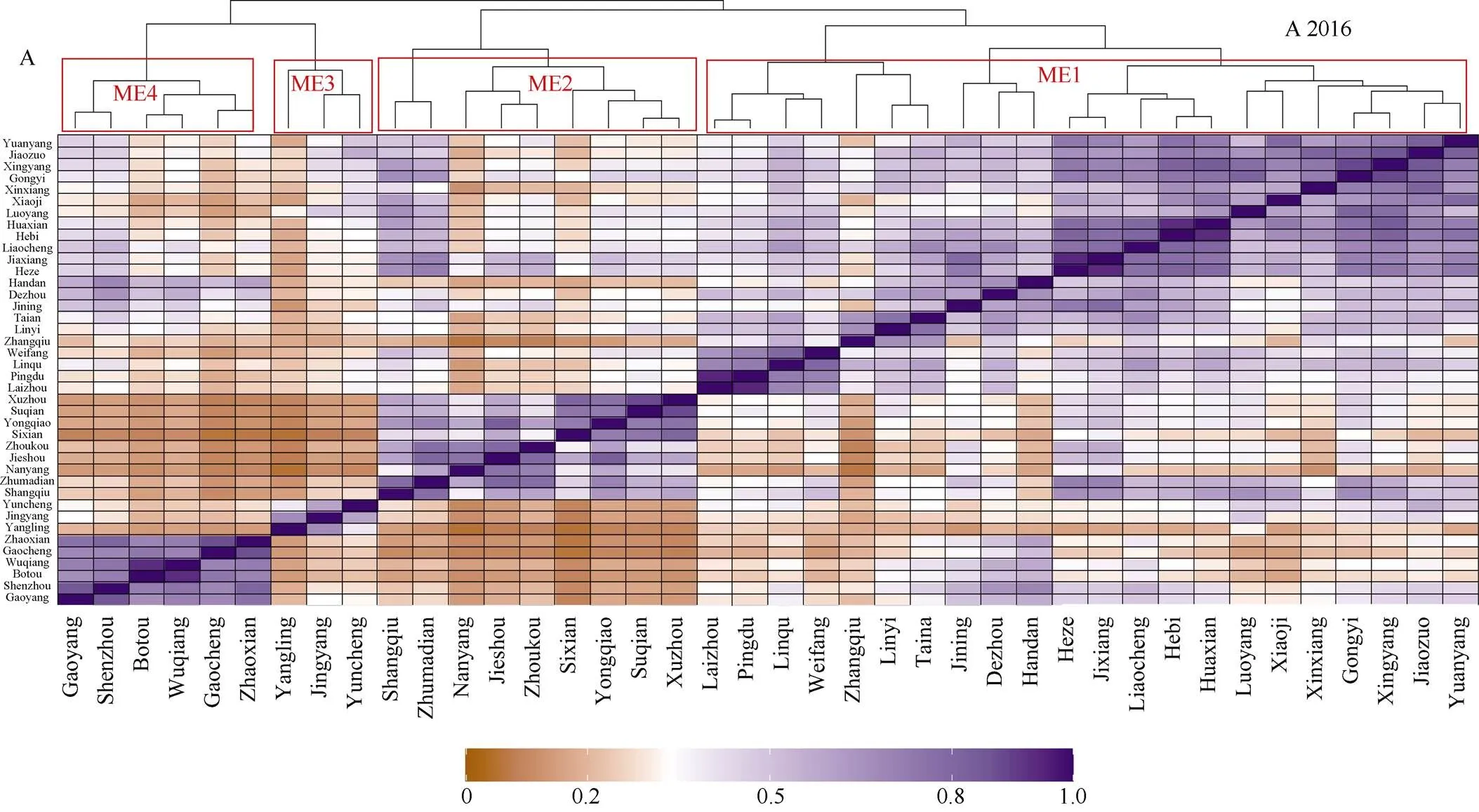
(图1)
ME1、ME2、ME3和ME4分别代表第1生态区、第2生态区、第3生态区和第4生态区。
ME1, ME2, ME3, and ME4 indicate mega-environments 1, mega-environments 2, mega-environments 3, and mega-environments 4, respectively.
2.4 2年气象因子相关性和主成分分析
2016年平均温度(mean)与饱和蒸汽压曲线斜率(svpc)、最低温度(min)、最高温度(max)呈现出高度正相关关系, 而与降雨量(prec)、降水亏缺(dbp)呈负相关。相对湿度(rh)与降雨量(prec)、饱和水汽压差(vpd)、潜在蒸散量(etp)呈高度负相关, 降雨量(prec)与潜在蒸散量(etp)、水平表面上的日射(sihs)、饱和水汽压差(vpd)、温度范围(range)呈高度负相关。2017年水平表面上的日射(sihs)与平均温度(mean)、最高温度(max)、最低温度(min)、潜在蒸散量(etp)、饱和蒸汽压曲线斜率(svpc)呈正相关, 而与降水亏缺(dbp)、饱和水汽压差(vpd)、降雨量(prec)呈负相关。相对湿度(rh)与温度范围(range)、饱和水汽压差(vpd)呈负相关, 而与降雨量(prec)呈正相关(图2)。
2016年主成分分析显示, 前2个主成分(PC1、PC2)累计贡献了72.7%的互作效应变异信息。第1生态区(ME1)呈现出较高的籽粒产量(GY), 第2生态区(ME2)主要特征是较高的饱和蒸汽压曲线斜率(svpc)、平均温度(mean)、最低温度(min)、温度对辐射利用效率的影响(frue)、水平表面上的日射(sihs)和相对湿度(rh)。第3生态区(ME3)呈现出较高的饱和水汽压差(vpd)和温度范围(range)。相反, 第4生态区(ME4)呈现出较低的饱和蒸汽压曲线斜率(svpc)、平均温度(mean)和温度对辐射利用效率的影响(frue) (图3-A)。2017年主成分分析PC1解释了互作效应的41.5%, PC2解释了互作效应的31.1%, 共计解释了互作效应变异来源的72.6%。第1生态区(ME1)呈现出较高的降水亏缺(dbp)、相对湿度(rh)和降雨量(prec)。相反, 第3生态区(ME3)呈现出较低的降水亏缺(dbp)、相对湿度(rh)、降雨量(prec)和较高的饱和水汽压差(vpd)。第2生态区(ME2)主要特征是较高的潜在蒸散量(etp)和水平表面上的日射(sihs)。第4生态区(ME4)主要特征是较低的最低温度(min)和温度对辐射利用效率的影响(frue) (图3-B)。
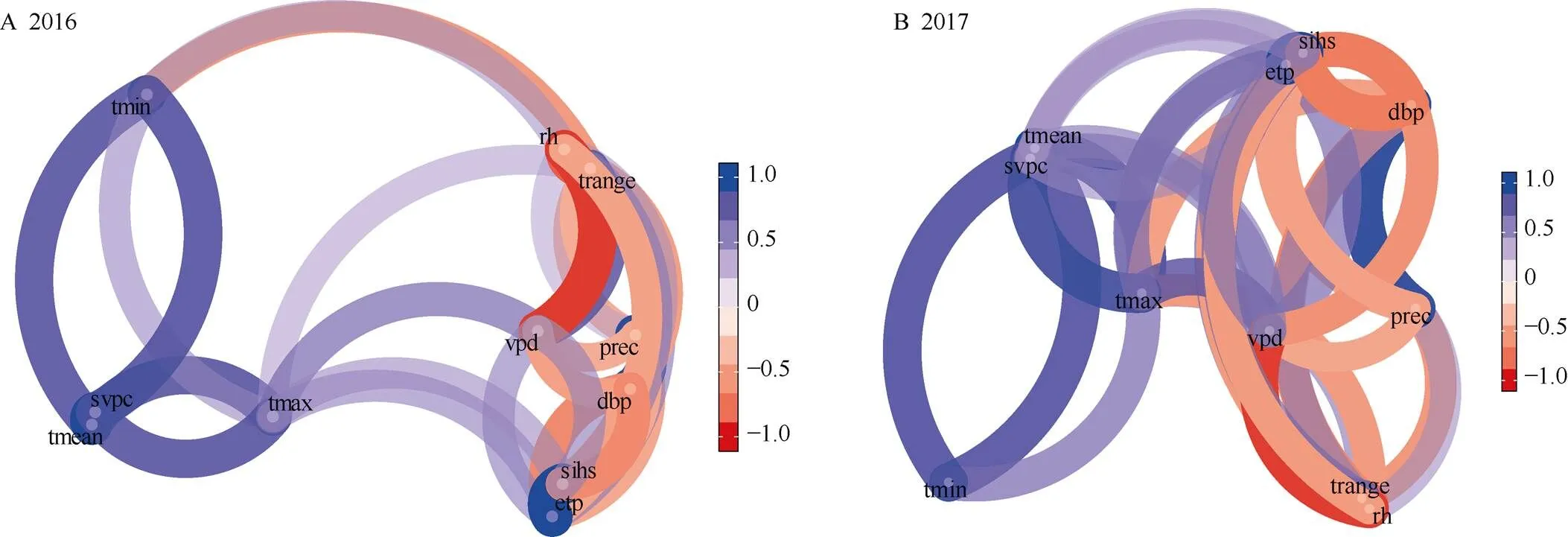
图2 2016年(A)和2017年(B)气象因子相关性分析
etp: 潜在蒸散量; dbp: 降水亏缺; rh: 大气湿度; sihs: 水平表面上的日射; svpc: 饱和蒸汽压曲线斜率;max: 最高温度;min: 最低温度;mean: 平均温度;range: 温度范围; vpd: 饱和水汽压差; prec: 降雨量。
etp: potential evapotranspiration; dbp: deficit by precipitation; rh: relative humidity; sihs: all sky insolation incident on a horizontal surface; svpc: slope of saturation vapor pressure curve;max: maximum temperature;min: minimum temperature;mean: mean temperature;range: daily temperature range; vpd: vapor pressure deficit; prec: rainfall precipitation.
2.5 不同气象因子物候期比较
2016年第2生态区(ME2)和第3生态区(ME3)分别在初始生长期(1-intial growing)和灌浆结实期(5-grain filling)表现出较高的降水亏缺(dbp), 而ME3和ME2分别在叶片扩展期I (2-leaf expansion I)和灌浆结实期(5-grain filling)呈现出较低的降水亏缺(dbp)。4个生态区在叶片扩展期II (3-leaf expansion II)均有较高的降水亏缺(dbp) (附图2)。2017年ME2和ME3在叶片扩展期II (3-leaf expansion II)表现出较高的降水亏缺(dbp), ME4在开花期(4-flowering)、ME1在灌浆结实期(5-grain filling)均呈现出较高的降水亏缺(dbp) (附图3)。
2016年ME4在初始生长期(1-intial growing), 60%以上的天数最高温度大于34.1℃, 其他生态区在这一时期最高温度大于34.1℃的比例在20%以下。ME3在叶片扩展期II (3-leaf expansion II)、开花期(4-flowering), 一半以上的天数最高温度大于34.1℃, ME2在开花期(4-flowering)接近一半的天数出现大于34.1℃的高温(附图4)。2017年ME3和ME4分别在叶片扩展期I (2-leaf expansion I)和叶片扩展期II (3-leaf expansion II)有一半以上的天数出现了大于35.2℃的高温, ME1在叶片扩展期II (3-leaf expansion II)接近一半的天数出现大于35.2℃的高温(附图5)。
2.6 黄淮海夏玉米参试品种基因型×性状(GT)双标图分析
2016年黄淮海夏玉米试验13个参试品种的GT双标图前2个主成分共计解释了互作效应变异来源的49.1% (PC1=28.8%, PC2=20.3%), 可以近似表达黄淮海夏玉米参试品种被测性状间的相关性以及基因型和性状间的互作效应。由图4-A得出以下信息: (1) 由原点(0, 0)出发连接每个被测性状形成不同的向量, 根据性状向量之间的夹角和分布将被测农艺性状划分为若干个相关向量群, 每个向量群内的性状彼此呈现出正相关关系, 不同向量群内的被测性状呈现出负相关或零相关。倒伏率、百粒重和秃尖长度群, 黑粉病和穗行数群, 籽粒产量、穗粒重、穗长、株高和穗位高群。(2) 生育期、茎腐病和空秆率之间彼此呈现出微弱正相关和负相关关系, 其中生育期与茎腐病、黑粉病、穗行数、穗位高等性状呈现出显著负相关关系。空秆率与籽粒产量、穗粒重、穗长、株高、穗位高呈现出显著负相关关系。(3) 通过GT双标图可以直观看出品种的特征性状精确表达。例如, 唐玉6678 (TY6678)位于茎腐病和生育期图标附近, 且远离籽粒产量图标, 说明这个品种抗病性较差、生育期较长且产量较低。衡玉321 (Hengyu 321, HY321)和邯玉9112 (Hanyu 9112, HY9112)的穗粒重和穗长等性状较高且籽粒产量较高, 属于丰产型品种。中农大626 (Zhongnongda 626, ZND626)、中农大696 (Zhongnongda 696, ZND696)和潞玉36 (Luyu 36, LY36)等品种倒伏率较高、秃尖长度较大和黑粉病发病重, 属于丰产性较差的品种。
me: 生态区。各气象因子缩写同图2。
me: mega-environments. Abbreviations of the meteorological factors are the same as those given in Fig. 2.

图4 2016年(A)和2017年(B)参试品种GT双标图
HY321: 衡玉321; JF118: 冀丰118; LY1502: 潞研1502; LY36: 潞玉36; HY9112: 邯玉9112; SD510: 宿单510; JY518: 冀玉518; ZND626: 中农大626; ZND696: 中农大696; TY6678: 唐玉6678; JY196: 冀玉196; SD617: 宿单617; JY902: 冀玉902; JY757: 冀玉757; HY6105: 衡玉6105; ZD111: 中单111; LY1611: 潞研1611; TY6461: 唐玉6461; HY5177: 邯玉5177; ZD958: 郑单958。GY: 籽粒产量; CS: 瘤黑粉病; SR: 茎腐病; EER: 空秆率; PH: 株高; EH: 穗位高; GP: 生育期; LR: 倒伏率; EL: 穗长; KRN: 穗行数; BTL: 突尖长度; GWE: 穗粒重; HSW: 百粒重。
HY321: Hanyu 321; JF118: Jifeng 118; LY1502: Luyan 1502; HY9112: Hanyu 9112; SD510: Sudan 510; JY518: Jiyu 518; ZND626: Zhongnongda 626; ZND696: Zhongnongda 696; TY6678: Tangyu 6678; JY196: Jiyu 196; SD617: Sudan 617; JY902: Jiyu 902; JY757: Jiyu 757; HY6105: Hengyu 6105; ZD111: Zhongdan 111; LY1611: Luyan 1611; TY6461: Tangyu 6461; HY5177: Hanyu 5177; ZD958: Zhengdan 958. GY: grain yield; CS: common smut; SR: stalk rot; EER: empty ears rate; PH: plant height; EH: ear height; GP: growth period; LR: lodging rate; EL: ear length; KRN: kernel row number; BTL: bare tip length; GWE: grains weight per ear; HSW: hundred-seed weight.
2017年黄淮海夏玉米13个参试品种前2个主成分共计解释了58.1%的品种性状变异信息。籽粒产量与穗粒重、百粒重、穗长和生育期呈现出正相关关系, 而与倒伏率、空秆率呈现出显著负相关关系(图4-B)。株高除与穗位高和黑粉病呈正相关外, 和其余性状呈现负相关或微弱正相关关系。潞研1611 (Luyan 1611, LY1611)、冀丰118 (Jifeng 118, JF118)和唐玉6461 (Tangyu 6461, TY6461)等品种百粒重、穗长和穗粒重较高且籽粒产量较高, 属于丰产性较好的品种。中单111 (Zhongdan 111, ZD111)等品种除空秆率和倒伏率较高外, 其余性状表现较差, 属于丰产性较差的品种。虽然GT双标图可以展示品种性状优势, 但不能对品种综合评价, 对哪些品种值得推广种植说服力不够。
2.7 参试品种基因型×产量×性状(GYT)双标图分析
2016年参试品种GYT双标图前2个主成分解释了品种-产量-性状组合总变异信息的82.8% (PC1= 71.8%, PC2=11.0%) (图5-A)。鉴于每个向量-性状组合都含有产量组分, 除产量×黑粉病组合(Y×CS(-1))和产量×茎腐病组合(Y×SR(-1))外, 其余产量-性状组合间均呈正相关关系。在GT双标图中穗粒重与穗长和株高之间表现出极显著正相关关系以及产量与空秆率和倒伏率之间的极显著负相关关系在GYT双标图中得到了体现。结合表6和附表1可知, 除产量×秃尖长度组合(Y×BTL(-1))与产量×茎腐病组合(Y×SR(-1))为负相关外, 其余产量-组合间呈现出正相关关系。产量×黑粉病(Y×CS(-1))与其余产量-性状组合间差异均不显著, 产量×茎腐病(Y×SR(-1))除与产量×空秆率(Y×EER(-1))和产量×穗位高(Y× EH)间差异显著(<0.05)外, 与其余产量-性状组合间差异不显著。GYT双标图可以展现出品种间产量-性状互作关系, 邯玉9112 (HY9112)、宿单510 (SD510)和冀玉196 (JY196)等品种在产量×茎腐病(Y×SR(-1))组合上表现较好, 衡玉321 (HY321)在Y×KRN、Y×HSW、Y×GWE和Y×EL等产量-组合上表现较好, 冀丰118 (JF118)在Y×CS(-1)组合表现较好。参试品种理想指数是各品种产量-性状组合经过标准化处理后的平均值, 排列顺序和品种在ATA轴上的坐标位置一致。衡玉321 (HY321)和冀丰118 (JF118)是参试品种中理想指数较高的2个品种, 理想指数分别为1.24和1.01, 潞研36 (LY36)、潞研1502 (LY1502)和唐玉6678 (TY6678)理想指数较差, 对照品种郑单958理想指数在参试品种中位于中间位置。

表6 2016年产量与性状组合皮尔逊相关性分析
*和**分别表示在5%和1%概率水平差异显著,ns表示不显著, 表中产量指的是籽粒产量。GY: 籽粒产量; CS: 瘤黑粉病; SR: 茎腐病; EER: 空秆率; PH: 株高; EH: 穗位高; GP: 生育期; LR: 倒伏率; EL: 穗长; KRN: 穗行数; BTL: 突尖长度; GWE: 穗粒重; HSW: 百粒重。
*and**indicate significant difference at the 5% and 1% probability levels, respectively.ns: not significant. Yield in the table refers to grain yield. GY: grain yield; CS: common smut; SR: stalk rot; EER: empty ears rate; PH: plant height; EH: ear height; GP: growth period; LR: lodging rate; EL: ear length; KRN: kernel row number; BTL: bare tip length; GWE: grains weight per ear; HSW: hundred-seed weight.
2017年黄淮海参试品种GYT双标图前2个主成分累计解释了品种-产量-性状组合72.6% (PC1= 59%, PC2=13.6%)的总变异信息。由图5-B和表7、附表2可见, 除Y×CS(-1)、Y×KRN、Y×SR(-1)和Y× BTL(-1)外, 其余产量-性状组合间向量夹角均为锐角, 呈现出正相关关系。其中, Y×HSW、Y×GWE、Y×LR(-1)、Y×PH、Y×EH、Y×EL和Y×EER(-1)等产量-性状组合间彼此呈现出显著(<0.05)正相关, Y×CS(-1)与其余产量-性状组合呈现出负相关关系。DK56在Y×SR(-1)和Y×BTL(-1)等产量-组合上表现较好, 而在Y×CS(-1)组合上表现较差。宿单617 (Sudan 617, SD617)除在组合Y×CS(-1)上表现尚可外, 其余产量-性状组合表现均较差。冀玉902 (Jiyu 902, JY902)在所有的产量-性状组合上经标准化后的数值均是正值, 理想指数是参试品种中最高(0.88)的品种, 其次是DK56 (0.78)和PT1212 (0.59)。邯玉5177 (Hanyu 5177, HY5177)理想指数最低(–1.31)是所有参试品种中表现最差的品种, 对照品种郑单958除在Y×CS(-1)组合上表现较好外, 其余产量-性状组合表现较差, 理想指数仅次于邯玉5177 (HY5177)排在参试品种后面。2年数据显示, 与GT双标图相比, GYT双标图前2个主成分可以解释更多、且拟合度较好, 结果更有说服力。
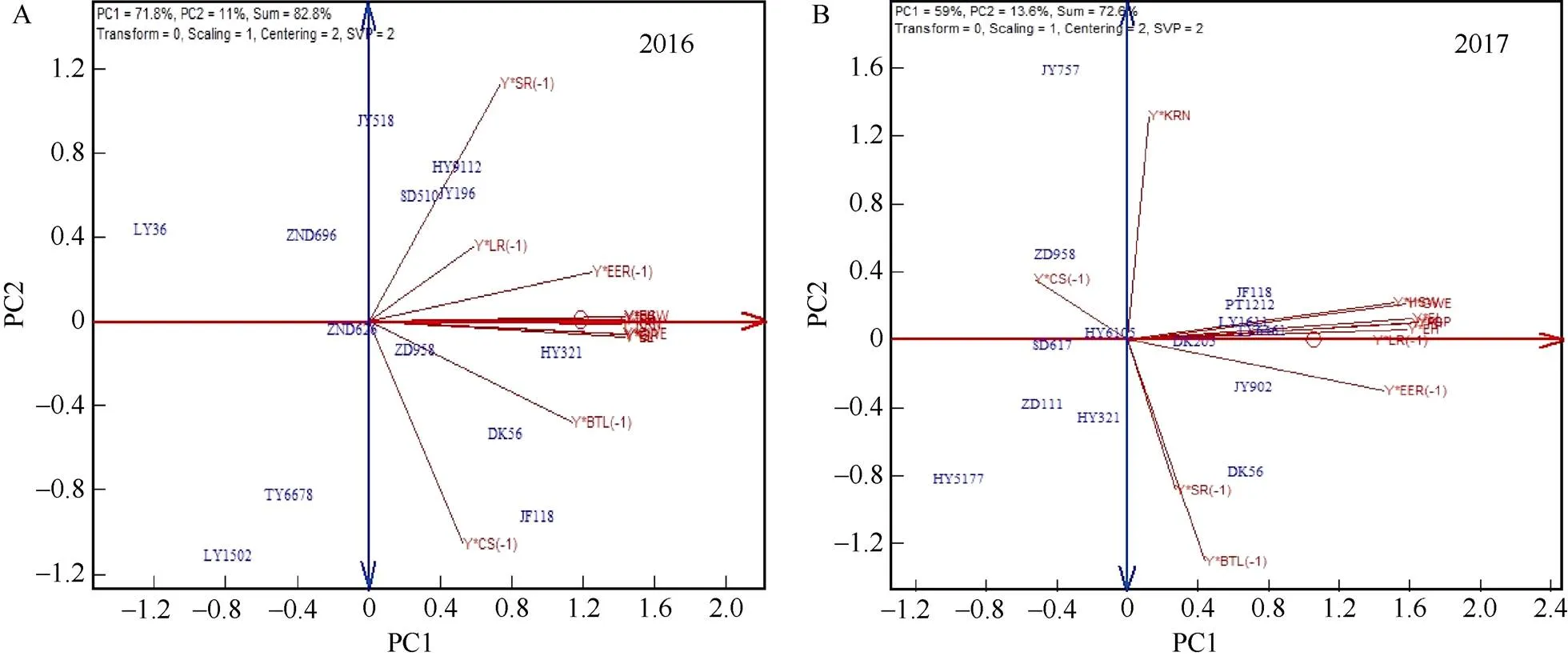
图5 2016年(A)和2017年(B)参试品种农艺性状GYT双标图
HY321: 衡玉321; JF118: 冀丰118; LY1502: 潞研1502; LY36: 潞玉36; HY9112: 邯玉9112; SD510: 宿单510; JY518: 冀玉518; ZND626: 中农大626; ZND696: 中农大696; TY6678: 唐玉6678; JY196: 冀玉196; SD617: 宿单617; JY902: 冀玉902; JY757: 冀玉757; HY6105: 衡玉6105; ZD111: 中单111; LY1611: 潞研1611; TY6461: 唐玉6461; HY5177: 邯玉5177; ZD958: 郑单958。Y× BTL (-1): 产量×秃尖; Y×CS (-1): 产量×瘤黑粉病; Y× EER (-1): 产量×空秆率; Y×EH: 产量×穗位高; Y×EL: 产量×穗长; Y×GP: 产量×生育期; Y×GWE: 产量×穗粒重; Y× HSW: 产量×百粒重; Y×KRN: 产量×穗行数; Y×LR (-1): 产量×倒伏率; Y×PH: 产量×株高; Y× SR (-1): 产量×茎腐病。
HY321: Hanyu 321; JF118: Jifeng 118; LY1502: Luyan 1502; HY9112: Hanyu 9112; SD510: Sudan 510; JY518: Jiyu 518; ZND626: Zhongnongda 626; ZND696: Zhongnongda 696; TY6678: Tangyu 6678; JY196: Jiyu 196; SD617: Sudan 617; JY902: Jiyu 902; JY757: Jiyu 757; HY6105: Hengyu 6105; ZD111: Zhongdan 111; LY1611: Luyan 1611; TY6461: Tangyu 6461; HY5177: Hanyu 5177; ZD958: Zhengdan 958. Y×BTL (-1): grain yield×bare tip length; Y×CS (-1): grain yield×common smut; Y× EER (-1): grain yield×empty ears rate; Y×EH: grains yield×ear height; Y×EL: grains yield×ear length; Y×GP: grains yield×growth period; Y×GWE: grains yield×grains weight per ear; Y× HSW: grains yield×hundred-seed weight; Y×KRN: grains yield×kernel row number; Y×LR (-1): grains yield×lodging rate; Y×PH: grains yield×plant height; Y× SR (-1): grains yield×common smut.

表7 2017年产量与性状组合皮尔逊相关性分析

(续表7)
*和**分别表示在5%和1%概率水平差异显著,ns表示不显著, 缩写同表6。
*and**indicate significant difference at the 5% and 1% probability levels, respectively.ns: not significant. Yield in the table refers to grain yield. GY: grain yield; CS: common smut; SR: stalk rot; EER: empty ears rate; PH: plant height; EH: ear height; GP: growth period; LR: lodging rate; EL: ear length; KRN: kernel row number; BTL: bare tip length; GWE: grains weight per ear; HSW: hundred-seed weight. Abbreviations are the same as those given in Table 6.
2.8 不同生态区参试品种评价
将参试品种与平均环境轴 (average environment axis, AEA)作一垂线, 垂线长短代表着该品种稳定性强弱, 垂线越短该品种越稳定, 越靠近AEA轴正方向的品种代表着丰产性越高。由图6可知, 2016年黄淮海参试品种在第1生态区(ME1)、第2生态区(ME2)、第3生态区(ME3)和第4生态区(ME4)前2个主成分分别解释了73.8%、79.4%、81.1%和71%的品种-产量-性状组合变异信息。在4个生态区中, 衡玉321 (HY321)和冀丰118 (JF118)丰产性和稳定性均在参试品种前列, 属于丰产稳产型品种。潞玉36 (LY36)和潞研1502 (LY1502)靠近AEA轴最左侧, 与AEA轴垂线较长, 这2个品种属于参试品种中丰产性稳定性均较差的品种。AEA轴将参试品种和产量-性状组合分为上下2部分, 位于同一侧的品种与之相邻的产量-性状组合都有较好的表现, 反之,表现较差。例如, 衡玉321 (HY321)和DK56在Y×HSW、Y×KRN、Y×PH、Y×EH、Y×GP、Y×EL和Y×GWE等产量-性状组合上表现较好, 而在Y×EER和Y×LR等产量-性状组合上表现较差。

图6 基于GYT双标图的2016年参试品种丰产性和稳定性分析
ME1: 第1生态区; ME2: 第2生态区; ME3: 第3生态区; ME4: 第4生态区。
ME1: mega-environments 1; ME2: mega-environments 2; ME3: mega-environments 3; ME4: mega-environments 4.
由图7可知, 2017年黄淮海参试品种前2个主成分分别在ME1、ME2、ME3和ME4上解释了66.1%、72.8%、63.8%和71.2%的品种-产量-性状组合变异信息。DK205、唐玉6461 (TY6461)、潞研1611 (LY1611)和DK56、冀玉902 (JY902)、PT1212分别是ME1和ME2丰产性较好的品种, 邯玉5177 (HY5177)是ME1和ME2中丰产性最差的品种。衡玉6105 (HY6105)、冀丰118 (JF118)、衡玉321 (HY321)和邯玉5177 (HY5177)、冀丰118 (JF118)、冀玉902 (JY902)分别是ME3和ME4表现较好的品种, 而中单111 (ZD111)和宿单617 (SD617)分别是ME3和ME4表现较差的品种。除ME2外, 衡玉321 (HY321)与AEA轴垂线较短, 呈现出较好的稳定性。DK56和对照品种郑单958在4个生态区稳定性表现较好。与2016年品种图标在GYT双标图中的位置类似, 2017年衡玉321 (HY321)和DK56同为AEA轴一侧, ME1、ME2和ME3中在Y×PH和Y× HSW等产量-性状组合上表现较好, 而在Y×CS(-1)、Y×LR(-1)等产量-组合上表现较差。
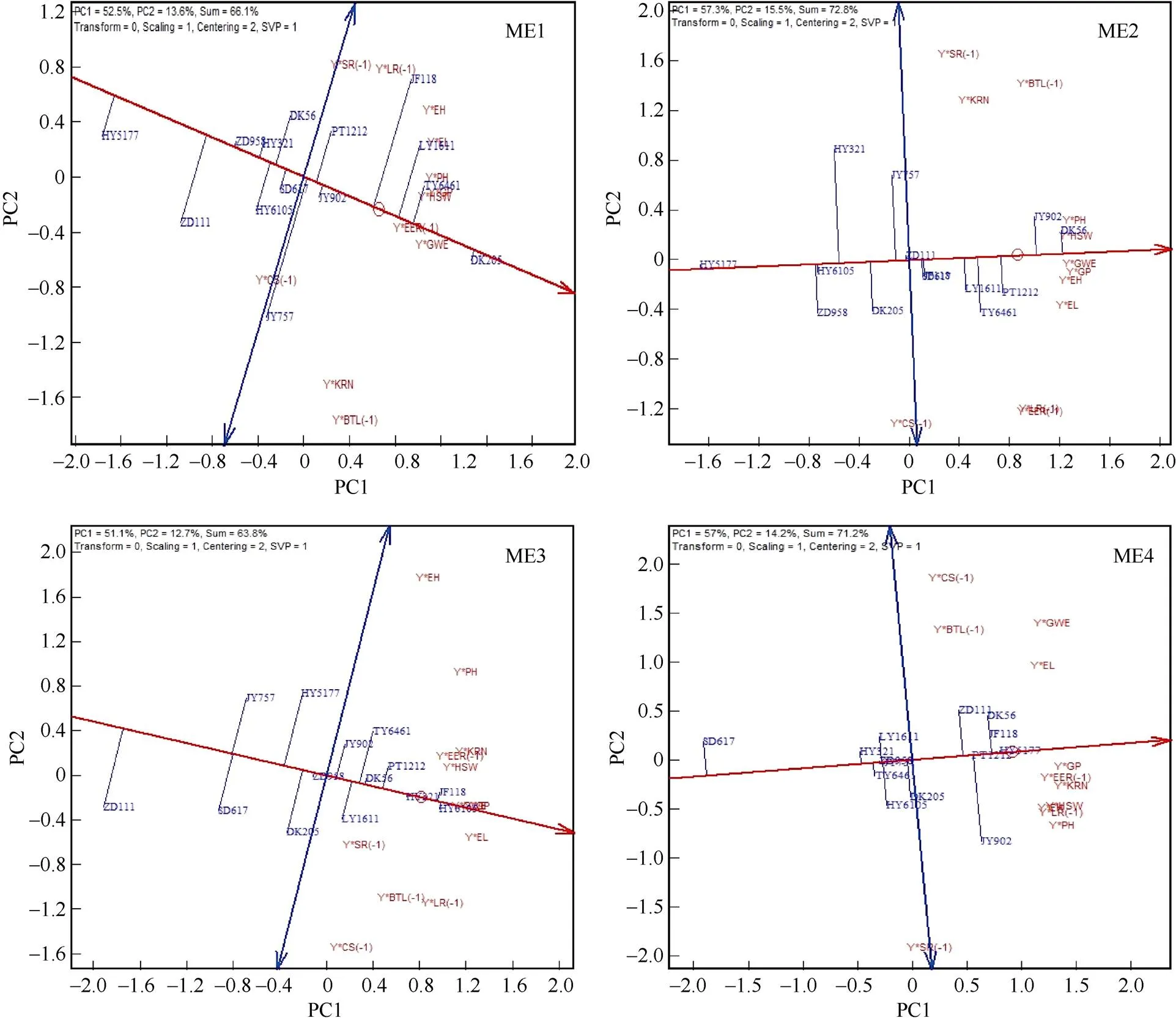
图7 基于GYT双标图的2017年参试品种丰产性和稳定性分析
ME1: 第1生态区; ME2: 第2生态区; ME3: 第3生态区; ME4: 第4生态区。
ME1: mega-environments 1; ME2: mega-environments 2; ME3: mega-environments 3; ME4: mega-environments 4.
3 讨论
3.1 不同年份生态区间的环境因子变化
2016—2017两年中包括的40个试点根据每年天气数据以19个环境协变量(EC)评价环境相似性, 最终被划分为4个不同生态区(ME)。同一试点在2年的气候因子表现是不同的。例如, 邯郸试点作为河北省试点中唯一一个与其他试点划分为不同的生态区,整个生长周期的大气湿度、饱和水气压差和最高温度等因子高度不同。同时期对比, 2016年灌浆结实期有较高的饱和水气压差(附图6), 开花期相对湿度较大(附图7), 初始生长期最高温度大部分时间>35℃, 而2017年饱和水汽压差较小(附图8), 相对湿度较小(附图9), 最高温度较低。邯郸试点2年间的气候变化较邯郸试点和莱州试点之间的变化要明显得多, 尽管这2个试点相距甚远, 且处于不同的海拔高度。这与Tao等[28]和Qin等[29]研究结果一致, 显示了黄淮海地区复杂多变的气候特征。这一结果可以利用植物生态生理学知识, 帮助确定基因型与环境相互作用的主要来源。在温暖的天气里, 通过蒸腾作用损失的水分要比寒冷的天气大。平均而言, ME3经历了更高的饱和水气压差, 通过将相对湿度和温度结合成一个单一的数量, 这是预测植物蒸腾作用的准确措施。值得注意的是, ME1作为2016年和2017年试点产量最高的生态区, 在开花期均呈现出较高的饱和水气压差。类似这种环境下的高产主要是由于在开花期由于降水不足以导致蒸腾作用受限, 在高饱和水气压差以及缺水地区的杂交种经常表现这种特征[30]。作为一种C4代谢植物, 玉米的光合作用最适温度比C3植物高, 这是因为二氧化碳浓缩系统的运行抑制了Rubisco加氧酶的活性。以前的研究表明, 玉米的净光合作用只有在叶温高于38℃时才会受到抑制, 而且光系统II的最大量子产量对45℃以下的叶温相对不敏感。当叶温逐渐升高时, Rubisco的激活和净光合作用通过新的激活酶多肽的表达而适应[31]。这种适应可能发生在ME1中, 并可能解释为什么2017年原阳和焦作等试点呈现出高产量, 即使约60%的开花期经历温度>35.2℃。另一个可以解释原阳试点在2017年高产的气候变量是最低温度。研究表明, 最低温度与籽粒产量之间呈负相关, 而原阳是2017年在籽粒灌浆阶段呈现最低温度<18.7℃的较小时期的试点。以前的研究表明, 温度低于20℃时, 光系统II的主要电子受体(QA)的氧化还原状态会突然上升, 并增加叶绿素荧光的非光化学淬灭, 表明在这种条件下电子传输受到限制[32]。
3.2 较高的降水量并不能确保较高的产量
2016年ME1试点在作物周期内呈现出较高的累积降水量(图3), 范围从平度的~310 mm到章丘的~630 mm。因此, 这些环境显示出较高的降水亏损(正亏损意味着更多的水供应), 以及较低的蒸汽压亏损。然而, ME1中的章丘、邯郸(~610)等试点没有出现预期的结果, 即较高的降水量带来较高的产量。邯郸和章丘在ME1中属于产量较低的试点, 分别为9004.15 kg hm–2和7928.65 kg hm–2。与同生态区的平度试点相比, 章丘比平度平均产量少2063 kg hm–2, 即使降水量高出约2倍。降水充沛的地区产量较低的原因可能是蒸腾作用受到限制。水从土壤中进入植物根部, 并通过细胞壁上的毛细管力导致的木质部内的负压, 移动到叶片。被太阳加热的水变成水蒸气, 通过气孔流出, 同时允许吸收二氧化碳进行光合作用[33]。这一过程的推动力由叶片细胞间空间和外部大气质量之间的水蒸气浓度差异来调节, 这一过程的能量由可用的辐射量提供。例如, 在2016年ME1中的章丘试点观察到较高的相对湿度, 在灌浆结实期有将近一半的时间出现约80%的数值。考虑到较低的太阳辐射量, 在ME1的试点中观察到较低的潜在蒸散量, 这可以极大地减少植物的蒸腾作用。通过在高蒸汽压缺失的情况下限制蒸腾作用以提高玉米的耐旱性[34], 但考虑到蒸腾驱动的质量流概念[35], 在湿度升高的情况下, 产量降低可能与矿物营养的获取受阻有关, 主要是那些通过质量流供应给植物根系的营养(NO3–、Ca+2和Mg+2)。
3.3 环境类型化有助于更好地理解基因型与环境的相互作用
在研究的所有评价性状中, 观察到基因型、环境及其相互作用之间存在显著差异, 证实了环境对性状表型表达的影响, 以前的研究中也观察到类似的报告[36-39]。伴随着过去几十年全球气候因子的变化, 未来粮食生产将受到环境条件的威胁这受到了越来越多的人共识[40-42]。因此, 在一定环境下, 气候变量对玉米产量和基因型变异的影响尤为必要。所研究年份之间的气候信息显示出明显的多样性, 使得在多种环境条件下选择优良的玉米基因型是有利的。在不可预测的复杂天气变化中, 种植产量表现稳定的品种, 对确保粮食安全至关重要。美国生产规模大、机械化程度高、生物技术发达, 再加上合理密植、科学施肥、生物防治病虫害、节水灌溉等措施和技术手段, 美国的玉米产量水平长期处于世界前列。2020年中国的玉米产量为6350 kg hm–2, 大约是美国产量水平的60%[43]。鉴于中美两国玉米产量的巨大差异, 如何选育出适应不同生态区域气候特点的优良玉米杂交种, 是保证中国玉米产业健康稳定发展的关键[44]。本研究中, 我们证实了之前其他研究中报道的黄淮海地区复杂气候条件, 并表明使用环境类型技术来定义基于气候变量的生态区, 有助于育种者更好地理解基因型与环境的相互作用。一些研究根据基因型在单一年份的反应来定义生态区[45-47], 但由于定义基因型效应的环境模式在不同年份可能会有很大变化, 这可能导致错误的建议。在大多数评估多个地点和年份的基因型的研究中, 跨年度的平均产量被用来填充一个双向表(基因型×地点), 进一步用于GGE双标图分析[48]。
3.4 GYT双标图在生态区中的应用
鉴于黄淮海地区复杂的气候特点, 选育出的玉米品种要想适应每年、每个地区的生态条件是不现实的, 但是通过生态区的划分可以明晰品种的特殊适应性, 即适应某一特定的生态区。品种-性状(GT)双标图是GGE双标图在多种农艺性状分析上的一种表现形式, 可以显示不同性状间的相关性, 可以对品种特性进行分类, 不足是难以按多性状对品种进行综合评价[15]。同时, 丰产性是评价目标性状的前提, 脱离作物产量单纯讨论品种稳定性是没有意义的。针对GT双标图的缺陷, 研究人员开发了一种新型的基于产量-目标性状组合的方式即品种-产量×性状(GYT)双标图对参试品种进行综合评价。GYT双标图技术将品种产量和目标性状以图形的方式相结合从而对品种进行评判, 对品种的优点和缺点可以同时显示。这种技术受到国内外越来越多研究人员的青睐[49-51]。基于气象因子划定不同的生态区, 采用GYT双标图技术评价参试品种的丰产性和稳定性可以做到品种推广的精细定位。本研究中, 2016年参试品种中衡玉321和冀丰118在划定的4个生态区中均表现出丰产性突出、稳定性较好的特征, 属于丰产稳产型品种。而潞玉36和潞研1502则属于参试品种中丰产性、稳定性均较差的品种。2017年参试品种中, DK56在ME2和ME4试点中表现出较好的丰产性和稳定性, DK205和衡玉6105分别在ME1和ME3环境中有较好的表现。对照品种郑单958两年区域试验表现出较好的稳定性但丰产性一般。在上述参试品种中衡玉321、DK205和衡玉6105等品种已通过国家审定, 在生产上得到了大面积的示范推广, 逐步成为当地主栽品种, 进一步印证了本研究方法的科学性和可行性。
4 结论
本研究是国内首次将环境分型技术和多性状选择相结合的案例, 以更好地了解基因型与环境的相互作用, 旨在为被测农艺性状获得满意的选择收益。评估了在2个作物季节中生长在40个试点的12个(2016年)和14个(2017年)玉米杂交种的13个农艺性状。大多数农艺性状均受到环境、基因型和基因型与环境相互作用的显著影响。2年研究结果表明, 通过使用当年285个环境协变量, 为每个研究年度划定了4个生态区。在本研究中, 利用GYT双标图技术, 在每个生态区中筛选出丰产性、稳定性不同的品种。2016年试验中衡玉321和冀丰118被4个生态区同时选中, 属于丰产性和稳定性较突出的品种。2017年试验中DK56在ME2和ME4试点环境中表现出较好的丰产性和稳定性, ME1和ME3中选品种分别是DK205和衡玉6105。
致谢:本研究中GGEbiplot软件由加拿大农业和农业食品部渥太华研发中心严威凯博士提供, 在此表示感谢。
附表和附图 请见网络版: 1) 本刊网站http://zwxb. chinacrops.org/; 2) 中国知网http://www.cnki.net/; 3) 万方数据http://c.wanfangdata.com.cn/Periodicalzuowxb.aspx。
[1] Haarhoff S J, Swanepoel P A. Plant population and maize grain yield: a global systematic review of rainfed trials., 2018, 58: 1819–1829.
[2] 王行川, 费继飞, 刘东胜, 高飞, 王爱芬, 原志强, 陈瑞杰, 孙招, 张长征, 王义波. 基于玉米新品种联创808选育的商业育种问题探讨. 玉米科学, 2020, 28(5): 14–19. Wang X C, Fei J F, Liu D S, Gao F, Wang A F, Yuan Z Q, Chen R J, Sun Z, Zhang C Z, Wang Y B. Breeding of a corn hybrid Lantron 808 and its implication for commercial breeding., 2020, 28(5): 14–19 (in Chinese with English abstract).
[3] Diouf I, Derivot L, Koussevitzky S, Carretero Y, Bitton F, Moreau L, Causse M. Genetic basis of phenotypic plasticity and genotype× environment interactions in a multi-parental tomato population., 2020, 71: 5365–5376.
[4] Ligarreto M G, Pimentel L C. Grain yield and genotype × environment interaction in bean cultivars with different growth habits., 2022, 25: 232–241.
[5] Yue H W, Gauch H G, Wei J W, Xie J L, Chen S P, Peng H C, Bu J Z, Jiang X W. Genotype by environment interaction analysis for grain yield and yield components of summer maize hybrids across the Huanghuaihai region in China., 2022, 12: 602.
[6] Shirazi S Z, Mei X, Liu B, Liu Y. Estimating potential yield and change in water budget for wheat and maize across Huang-Huai- Hai Plain in the future., 2022, 260: 107282.
[7] Ling M, Han H, Hu X, Xia Q, Guo X. Drought characteristics and causes during summer maize growth period on Huang-Huai- Hai Plain based on daily scale SPEI., 2023, 280: 108198.
[8] Yue H, Wei J, Xie J, Chen S, Peng H, Cao H, Jiang X. A study on genotype-by-Environment interaction analysis for agronomic traits of maize genotypes across Huang-Huai-Hai region in China., 2022, 91: 57–81.
[9] 徐云碧. 作物科学中的环境型鉴定(Envirotyping)及其应用. 中国农业科学, 2015, 48: 3354–3371. Xu Y B. Envirotyping and its applications in crop science., 2015, 48: 3354–3371 (in Chinese with English abstract).
[10] Xu Y. Envirotyping for deciphering environmental impacts on crop plants., 2016, 129: 653–673.
[11] Costa-Neto G, Fritsche-Neto R, Crossa J. Nonlinear kernels, dominance, and envirotyping data increase the accuracy of genome- based prediction in multi-environment trials., 2021, 126: 92–106.
[12] Smith D T, Potgieter A B, Chapman S C. Scaling up high- throughput phenotyping for abiotic stress selection in the field., 2021, 134: 1845–1866.
[13] 岳海旺, 魏建伟, 谢俊良, 姚文影, 曹红梅, 陈淑萍, 彭海成, 卜俊周. 基因型和环境互作对黄淮海夏玉米品种籽粒产量的影响. 中国农业大学学报, 2022, 27: 31–43. Yue H W, Wei J W, Xie J L, Yao W Y, Cao H M, Chen S P, Peng H C, Bu J Z. Effect of interaction between genotype and environment on the grain yield of summer maize hybrids in Huanghuaihai region., 2022, 27: 31–43 (in Chinese with English abstract).
[14] Yan W, Nilsen K T, Beattie A. Mega-environment analysis and breeding for specific adaptation., 2023, 63: 480–494.
[15] Yan W K, Frégeau-Reid J. Genotype by yield × trait (GYT) biplot: a novel approach for genotype selection based on multiple traits., 2018, 8: 8242.
[16] Elfanah A M, Darwish m A, Selim A I, Shabana M M, Elmoselhy O M, Khedr R A, Abdelhamid M T. Spectral reflectance indices’ performance to identify seawater salinity tolerance in bread wheat genotypes using genotype by yield × trait biplot approach., 2023, 13: 353.
[17] 许乃银, 赵素琴, 张芳, 付小琼, 杨晓妮, 乔银桃, 孙世贤. 基于GYT双标图对西北内陆棉区国审棉花品种的分类评价. 作物学报, 2021, 47: 660–671. Xu N Y, Zhao S Q, Zhang F, Fu X Q, Yang X N, Qiao Y T, Sun S X. Retrospective evaluation of cotton varieties nationally registered for the Northwest Inland cotton growing regions based on GYT biplot analysis., 2021, 47: 660–671 (in Chinese with English abstract).
[18] NY/T 1209–2020. 农作物品种试验与信息化技术规程: 玉米. 北京: 中国农业出版社, 2020. NY/T 1209–2020. Regulations for the Variety Tests and Information of Field Crop: Maize (L.). Beijing: China Agriculture Press, 2020 (in Chinese).
[19] Costa-Neto G, Galli G, Carvalho H F, Crossa J, Fritsche-Neto R. EnvRtype: a software to interplay environs and quantitative genomics in agriculture., 2021, 11: jkab040.
[20] Sparks A H. Nasapower: a NASA POWER global meteorology, surface solar energy and climatology data client for R., 2018, 3: 1035.
[21] Ghanem M E, Marrou H, Sinclair T R. Physiological phenotyping of plants for crop improvement., 2015, 20: 139–144.
[22] Munda S, Sarma N, Lal M, Aromatic M, Group E P. G × E interaction of 72 accessions with three year evaluation ofJowitt. using regression coefficient and Additive Main effects and Multiplicative Interaction model (AMMI)., 2020, 146: 112169.
[23] Yan W, Holland J B. A heritability-adjusted GGE biplot for test environment evaluation., 2010, 171: 355–369.
[24] Ghazvini H, Bagherikia S, Pour-Aboughadareh A, Sharifalhossaini M, Razavi S A, Mohammadi S, Khakizade G. GGE biplot analysis of promising barley lines in the cold regions of Iran., 2022, 36: 461–472.
[25] Yan W, Hunt L A, Sheng Q, Szlavnics Z. Cultivar evaluation and mega-environment investigation based on the GGE biplot., 2000, 40: 597–605.
[26] Abu N E, Uguru M I, Obi I U. Genotype by trait relations of yield and yield components in aromatic peppers () based on GT biplot., 2011, 3: 382–390.
[27] Tang Q Y, Zhang C X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research., 2013, 20: 254–260.
[28] Tao F, Xiao D, Zhang S, Zhang Z, Rötter R P. Wheat yield benefited from increases in minimum temperature in the Huang- Huai-Hai Plain of China in the past three decades., 2017, 239: 1–14.
[29] Qin J, Xu R, Li H. Yang C Y, Liu D, Liu Z X, Zhang L F, Lu W G, Frett T, Chen P Y, Zhang M C. Evaluation of productivity and stability of elite summer soybean cultivars in multi-environment trials., 2015, 206: 759–773.
[30] Shekoofa A, Sinclair T R, Messina C D, Cooper M. Variation among maize hybrids in response to high vapor pressure deficit at high temperatures., 2016, 56: 392–396.
[31] Crafts-Brandner S J, Salvucci M E. Sensitivity of photosynthesis in a C4plant, maize, to heat stress., 2002, 129: 1773–1780.
[32] Sowiński P, Fronk J, Jończyk M, Grzybowski M, Kowalec P, Sobkowiak A. Maize response to low temperatures at the gene expression level: a critical survey of transcriptomic studies., 2020, 11: 576941.
[33] Slam M S, Hasan M K, Islam m R, Chowdhury M K, Pramanik M H, Iqbal M A, Rajendran K, Iqbal R, Soufan W, Kamran M, Sabagh A. Water relations and yield characteristics of mungbean as influenced by foliar application of gibberellic acid (GA3)., 2023, 11: 1048768.
[34] Yang Z, Sinclair T R, Zhu M, Messina C D, Cooper M, Hammer G L. Temperature effect on transpiration response of maize plants to vapor pressure deficit., 2012, 78: 157–162.
[35] Cramer M D, Hawkins H J, Verboom G A. The importance of nutritional regulation of plant water flux., 2009, 161: 15–24.
[36] Kamutando C N, Muungani D, Masvodza D R, Gasura E. Exploiting genotype × environment interaction in maize breeding in Zimbabwe., 2013, 8: 4058–4066.
[37] Mebratu A, Wegary D, Mohammed W, Teklewold A, Tarekegne A. Genotype × environment interaction of quality protein maize hybrids under contrasting management conditions in Eastern and Southern Africa., 2019, 59: 1576–1589.
[38] Arunkumar B, Gangapp E, Ramesh S L, Savithramma D, Nagaraju N, Lokesha R. Stability analysis of maize (L.) hybrids for grain yield and its attributing traits using Eberhart and Russel model., 2020, 39: 52–63.
[39] Adu G B, Badu-Apraku B, Akromah R, Haruna A, Amegbor I K, Amadu M K. Grain yield and stability of early-maturing single- cross hybrids of maize across contrasting environments., 2019, 33: 776–796.
[40] Ceglar A, Kajfež-Bogataj L. Simulation of maize yield in current and changed climatic conditions: addressing modelling uncertainties and the importance of bias correction in climate model simulations., 2012, 37: 83–95.
[41] Das J, Poonia V, Jha S, Goyal M K. Understanding the climate change impact on crop yield over Eastern Himalayan Region: ascertaining GCM and scenario uncertainty., 2020, 142: 467–482.
[42] de Araujo M J, de Paula R C, Campoe O C, Carneiro R L. Adaptability and stability of eucalypt clones at different ages across environmental gradients in Brazil., 2019, 454: 117631.
[43] Guo Q, Huang G, Guo Y, Zhang M, Zhou Y, Duan L. Optimizing irrigation and planting density of spring maize under mulch drip irrigation system in the arid region of Northwest China., 2021, 266: 108141.
[44] Yue H W, Wei J W, Xie J L, Chen S P, Peng H C, Bu J Z. A study on genotype × environment interactions for the multiple traits of maize hybrids in China., 2021, 113: 4889–4899.
[45] Singh C, Gupta A, Gupta V, Kumar P, Sendhil R, Tyagi B S. Genotype × environment interaction analysis of multi-environment wheat trials in India using AMMI and GGE biplot models., 2019, 19: 309–318.
[46] Mushayi M, Shimelis H, Derera J, Shayanowako A I T, Mathew I. Multi-environmental evaluation of maize hybrids developed from tropical and temperate lines., 2020, 216: 1–14.
[47] Enyew M, Feyissa T, Geleta M, Tesfaye K, Hammenhag C, Carlsson A S. Genotype by environment interaction, correlation, AMMI, GGE biplot and cluster analysis for grain yield and other agronomic traits in sorghum (L. Moench)., 2021, 16: e0258211.
[48] Shojaei S H, Mostafavi K, Bihamta M R, Omrani A, Mousavi S M N, Illés Á. Stability on maize hybrids based on GGE biplot graphical technique., 2022, 12: 394.
[49] 岳海旺, 韩轩, 魏建伟, 郑书宏, 谢俊良, 陈淑萍, 彭海成, 卜俊周. 基于GYT双标图分析对黄淮海夏玉米区域试验品种综合评价. 作物学报, 2023, 49: 1230–1247. Yue H W, Han X, Wei J W, Zheng S H, Xie J L, Chen S P, Peng H C, Bu J Z. Comprehensive evaluation of maize hybrids tested in Huang-Huai-Hai summer maize regional trial based on GYT biplot analysis., 2023, 49: 1230–1247 (in Chinese with English abstract).
[50] Shojaei S, Mostafavi K, Ansarifad I, Bihamta M, Zeinalzadeh-Tabrizi H, Omrani A, Mousavi S M N. Comparison of genotype × trait and genotype × yield-trait biplots in Sunflower cultivars., 2023, 7: 136–147.
[51] Gholizadeh A, Ghaffari M, Siahbidi Z, Rezaeizad A. Multi-trait selection in sunflower hybrids using double and triple biplots., 2022, 14: 29–41.
Comprehensive evaluation of maize hybrids in the mega-environments of Huanghuaihai plain based on GYT biplot analysis
YUE Hai-Wang, WEI Jian-Wei, LIU Peng-Cheng, CHEN Shu-Ping, and BU Jun-Zhou*
Dryland Farming Institute, Hebei Academy of Agriculture and Forestry Sciences / Hebei Provincial Key Laboratory of Crops Drought Resistance Research, Hengshui 053000, Hebei, China
The selection of superior maize hybrids under different climatic types and multiple traits has been a difficult problem for crop breeders. It is necessary to explore the combination of envirotyping techniques (ET) and multi-trait selection for the comprehensive evaluation of the participating hybrids in the summer maize regional trials in the Huanghuaihai plain, which can provide a theoretical basis for the rational layout of hybrids. In this study, based on the data of the Huanghuaihai summer maize group from 2016 to 2017 were used, 40 sites were divided into different mega-environments (ME) by envirotyping techniques based on 19 environmental covariates in the same year. The combination performance of agronomic traits such as growth period, plant height, ear height, lodging rate, empty ears rate, ear length, bare tip length, kernels row number, grains weight per ear, hundred-seed weight, stalk rot, and common smut in the different mega-environments was comprehensively evaluated by using the GYT biplot technique. The ANOVA of the Additive Main effects and Multiplicative Interaction (AMMI) model showed that genotype (G), environment (E), and GE effects for the evaluated agronomic traits reached highly significant level (<0.01) in 2016 and that genotype, environment, and GE effects for the evaluated agronomic traits reached highly significant level in 2017, except for the GE effect of ear height, which was not significant. The 40 sites located in eight provinces were divided into four MEs based on the meteorological factor information of the current year, and meteorological factors deficit by precipitation (dbp), maximum temperature (max), vapor pressure deficit (vpd), and relative humidity (rh) showed a large trend during the five phenological periods. Among the hybrids evaluated in 2016, Hengyu 321 and Jifeng 118 both showed outstanding productivity and good stability in the four mega-environments, which belonged to the productive and stable genotypes. Among the genotypes evaluated in 2017, DK 56 showed a more harmonious yield-trait combination in ME2 and ME4, while DK205 and Hengyu 6105 performed better in ME1 and ME3, respectively. The control hybrid Zhengdan 958 had a good stability but an average productivity in the two-year regional trials. In conclusion, based on the combination of environtyping techniques to divide mega-environments and GYT biplot, we evaluated the high-yielding, the adaptability and stability of the evaluated hybrids, which could realize the fine positioning of genotype promotion, and provide a theoretical basis for the comprehensive evaluation of multiple traits of genotypes in the summer maize area of the Huanghuaihai plain.
summer maize hybrid; mega-environment; genotype-environment interaction; climatic variables; genotype × yield × trait biplot (GYT biplot)
10.3724/SP.J.1006.2024.33034
本研究由河北省“三三三人才工程”人才培养项目(A202101056), 河北省重点研发计划项目(20326305D), 财政部和农业农村部国家现代农业产业技术体系建设专项(玉米, CARS-02), 河北省玉米现代种业科技创新团队(21326319D), 河北省农林科学院科技创新专项课题(2023KJCXZX-HZS-1)和河北省农林科学院科技创新专项课题(2023KJCXZX-HZS-12)资助。
This study was supported by the “Three-Three-Three Talent Project” Funded Project in Hebei Province (A202101056), the Key Research and Development Projects of Hebei Province (20326305D), the China Agriculture Research System of MOF and MARA (Maize, CARS-02), Hebei Provincial Maize Modern Seed Industry Science and Technology Innovation Team (21326319D), the HAAFS Science and Technology Innovation Special Project (2023KJCXZX-HZS-1), and the HAAFS Science and Technology Innovation Special Project (2023KJCXZX-HZS-12).
卜俊周, E-mail: bujunzhou@126.com
E-mail: yanjiu1982@163.com
2023-05-22;
2023-10-23;
2023-11-29.
URL: https://link.cnki.net/urlid/11.1809.S.20231129.1031.002
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
房地产导刊(2021年10期)2021-11-22 07:43:34
西部论丛(2020年2期)2020-10-21 09:27:54
江西建材(2018年4期)2018-04-10 12:36:58
IT时代周刊(2015年8期)2015-11-11 05:50:49
生物进化(2015年4期)2015-02-14 02:12:08
现代检验医学杂志(2015年6期)2015-02-06 01:44:02
实验动物与比较医学(2014年5期)2014-02-28 14:53:10
中国糖料(2013年1期)2013-01-22 12:28:23
中国烟草学报(2012年4期)2012-04-09 07:11:30
医学综述(2011年18期)2011-12-10 01:09:01
