
钙在作物生长发育中的功能及应用
2024-03-28 02:35:44郜耿东葛萌萌葛贤宏周广生傅廷栋
作物学报 2024年4期
王 宇 郜耿东 葛萌萌 常 影 谭 静 葛贤宏 王 晶 汪 波 周广生 傅廷栋
综述
钙在作物生长发育中的功能及应用
王 宇 郜耿东 葛萌萌 常 影 谭 静 葛贤宏 王 晶 汪 波 周广生*傅廷栋
华中农业大学植物科学技术学院, 湖北武汉 430070
钙是作物必需的中量元素之一, 广泛存在于根、茎、叶、花、果实、种子中, 对作物的生长发育具有重要意义。钙属于再利用难的元素, 其吸收、转运受制于蒸腾作用, 因此农作物常发生生理性缺钙, 从而导致抗逆性减弱, 产量和品质降低。作物体内的钙具有双重功能, 既参与细胞壁与细胞膜的构成, 还可作为细胞内第二信使参与多种环境刺激和内部生长发育信号的响应。细胞中钙的吸收和转运对于维持细胞内钙离子稳态和钙信号精准传导至关重要。近年来, 钙在作物生长与发育中的功能和在作物生产中的应用得到了广泛研究。本文阐述了作物体内钙元素的分布、吸收转运和需求状况, 介绍了作物缺钙症状和原因, 并综述了钙的营养结构功能和第二信使功能及钙信号产生、传导和解码机制, 总结了钙在作物生长发育中的作用, 包括对产量、品质和抗逆性等方面的影响, 同时对其未来研究方向提出了展望。
作物; 缺钙; 钙吸收; 钙转运蛋白; 第二信使; 抗逆性
作物从土壤中汲取水分和矿质元素, 充足的矿质元素是作物正常生长和高产的前提。作物所需矿质元素种类和数量根据品种、种植环境、栽培目的有所不同, 因此认识和掌握矿质元素的特性和使用方法对于农作物高产、高质、高效栽培至关重要。氮磷钾是作物所必需的3种大量营养元素, 氮磷钾对于植物生长发育的影响也因此成为经久不衰的研究热点; 而关于中微量元素的肥料效应研究起步较晚, 且相对分散。氮磷钾的肥效研究和应用经过长足发展, 目前对作物产量的提升效果在一定程度上已达到了瓶颈, 且养分报酬递减现象愈发严重, 氮磷钾过度使用的弊端也逐渐暴露。20世纪末和21世纪初, 人们开始考虑中、微量元素的效应和应用, 包括钙、镁、硅、硫、锌、钼、硼等元素, 同时开发中、微量元素配合、大量元素和中微量元素配合等配施技术, 取得不错的效果[1], 例如花生中可以实现以钙促氮、降低氮肥用量, 协调干物质积累与产量, 硅钙磷镁钾肥配施能增加小麦产量, 同时改良土壤[2-3]。
钙是作物除氮磷钾外所需的第四大重要营养元素, 属于中量元素, 同时也是一种特殊营养元素, 具有区别于其他营养元素的信使功能[4]。钙主要通过根系被作物从土壤中吸收, 广泛分布于植物的根、茎、叶、花、果实和种子等部位, 钙在作物中调控细胞极性生长和细胞壁生物合成, 对根毛发育、花粉管伸长、茎秆和果实发育等方面具有重要调节作用, 同时在提高作物光合效率、抗逆性及抗倒性等方面也具有重要意义[5]。本文就作物钙分布、吸收、功能和应用研究等进展进行了综述。
1 作物钙吸收转运机制
作物的矿质元素供给主要靠根系从周围土壤溶液中吸收, 通过共质体或质外体途径运输至中柱, 再经木质部导管长距离向上运输至整个地上部植株[6]。作物蒸腾作用产生蒸腾拉力, 从而在植物和土壤之间形成从叶片开始沿茎、根到土壤的水势梯度, 水分由此进入根系内部, 许多矿质元素也因此随水分移动至根表面被作物吸收, 这种现象也称质流。钙就主要以质流形式移至根表, 再经横向短距离运输至木质部, 该过程主要依靠质外体途径运输。但是根系内皮层细胞壁上存在木栓化的凯氏带, 会阻止钙进入内皮层, 所以钙的吸收主要发生在凯氏带尚未(完全)形成的根尖处, 或通过细胞膜上的Ca2+通道进入内皮层细胞, 进而通过共质体途径穿过内皮层细胞进入中柱木质部[7]。进入木质部的钙受蒸腾作用影响, 随木质部液流向上运输至各组织部位附近, 随后通过Ca2+通道卸载流入组织细胞, 因此各组织部位钙含量通常与蒸腾作用强度呈正相关[8]。有趣的是, 在果树和荚果类植物的钙吸收研究中发现, 钙流入果实并不完全通过木质部/共质体途径, 韧皮部/共质体途径也参与果实钙吸收过程[9]。对荔枝、柑橘和龙眼进行母枝环剥处理, 韧皮部途径被切断, 会显著降低果实钙含量, 采用染料示踪试验也发现果柄中的钙多分布在韧皮部[10]。
钙在植物体内的移动性较差[11], 一方面因为植物细胞壁中存在许多羧基残基, 能够与钙离子形成果胶酸钙, 因此质外体钙在运输过程中会被截获一部分, 同时果胶酸钙的形成又会降低细胞壁透性, 也会限制钙的质外体运输[12]; 另一方面, 钙通过共质体运输时, 胞质钙浓度增加, 除了一部分通过胞质连丝参与共质体运输, 还有一部分会被钙泵排至胞外, 或进入细胞器中形成草酸钙和磷酸钙被贮存起来[13]。类似钙这类移动性较差的营养元素, 根系营养或叶面营养都无法保证结实部位的营养供给, 生产上经常采用针对性喷施, 尤其是果实和荚果类作物, 可以直接将钙施在果实上, 通过非维管束吸收途径直接补充养分[14]。
2 作物体内钙分布情况
植物体内的钙含量约为0.1%~5.0%, 主要以游离钙和结合钙的形式存在, 两者可相互转换[13,15]。钙在作物中的移动和钙信号的传导主要通过游离钙实现[16];而结合钙又根据结合对象大小可分为2类: 一类是与草酸、磷酸等小分子有机酸及简单分子配体结合, 其中主要是与有机酸结合的草酸钙; 另一种就是与蛋白质、酶、多糖和脂质等生物大分子或大分子复合物结合, 主要是存在细胞壁中的果胶酸钙[17-18]。根据作物的物种、部位和器官不同, 钙含量差异很大, 总体表现为双子叶作物的钙含量高于单子叶作物, 老叶中的钙含量比新叶中的多, 果实和种子的钙含量相对较少, 另外在顶端分生组织中的钙含量较高[19-20]。例如油菜收获时各器官钙含量大小顺序为落叶 > 角壳 > 根、茎 > 籽粒[21]。而在细胞水平上, 钙主要存在于细胞壁和细胞器中, 为了保证胞内钙信号的正常传导和防止钙离子毒性, 细胞质内钙含量则较少[14,22]。细胞壁是最大的钙库, 细胞壁中的钙主要与果胶结合; 叶绿体、液泡、内质网等细胞器也有钙的分布, 其钙浓度也是细胞质的几百甚至上千倍, 其中液泡中的钙含量最高, 能达到80 mmol L–1以上, 主要以草酸钙和磷酸钙形式存在, 是重要的钙贮存库[22-23]。
3 作物钙需求及缺钙症状
作物在不同发育时期以及不同组织部位对钙的需求特性存在较大差异。作物从营养阶段到生殖阶段, 直至成熟阶段, 钙积累量总体上呈现先增加后降低的趋势。例如, 油菜在苗期钙积累量增加缓慢, 而在薹花期积累速率会迅速增加, 直至进入角果发育期达到最大钙积累值, 而角果成熟期钙积累量有所下降, 这与油菜干物质累积动态趋势基本一致, 说明薹花期是油菜钙吸收的最大效率期。成熟期油菜各组织部位的钙含量占比分别为: 籽粒13%~ 18%、茎秆30%~34%、角壳48%~55%, 平均每100 kg油菜籽的形成需要吸收约6.2 kg CaO[24]。不同作物钙需求特性也不尽相同, 一般双子叶作物的需钙量要高于单子叶作物, 如属于十字花科的油菜、白菜等作物钙含量比较高, 可以达到5000 mg kg–1以上, 典型喜钙作物花生的植株钙含量能达到12,000 mg kg–1以上, 平均每100 kg花生荚果的形成需要约2.5 kg CaO, 而高粱、玉米和水稻等禾本科作物的钙含量大多在2000 mg kg–1以下[25-26]。十字花科作物的高钙含量可能与其较高的果胶含量有关, 细胞壁中的果胶可以结合钙离子。不同作物的缺钙临界值也有所不同, 含钙量高的作物更易发生缺钙现象, 比如白菜、番茄(叶片)的缺钙临界值是体内钙含量少于1.5%, 大豆、花生(叶片)的临界值约在1.0%, 而水稻、小麦等禾本科作物因为需钙量少, 缺钙临界值多在0.5%以下[27-29]。
作物的缺钙通常首先发生在幼嫩组织, 一般表现为顶芽、侧芽和根尖等分生组织发生卷曲畸形, 叶缘开始变黄并逐渐坏死, 严重时出现生长点坏死。例如油菜缺钙时, 表现为植株矮小, 主侧根生长被抑制, 新生叶片向上卷曲并伴随斑点出现, 角果数减少[30-31]。玉米、水稻缺钙症状表现为幼根畸形, 叶片卷曲且伴随锯齿状的叶缘, 叶片易扭曲粘连, 导致新生叶无法正常伸展, 逐渐干枯坏死, 形成类似“牛尾巴”症状[32-34]。番茄缺钙导致植株发育不良, 叶片萎蔫且边缘发黄, 生长点坏死, 脐腐病发病率增加[35]。另外, 钙缺乏经常发生在作物钙积累速率最快的时期, 比如果树在落花后20~30 d和果实膨大期所需钙分别占全生育期的30%和50%, 此时钙供应不足会导致果实品质下降, 出现畸形果和果实开裂的现象, 苦痘病发病率增加[36]。油菜薹花期和角果期的钙积累量快速增加, 分别占全生育期钙总积累量的32%和36%, 此时缺钙易导致顶花脱落, 花蕾黄化干枯, 胚珠受精率低、易败育, 茎秆呈紫红色且伴随开裂现象[37]。花生荚果发育期缺钙会造成荚果成熟期会继续开花, 出现“老来青”现象, 这是因为缺钙降低光合产物运输效率, 更多同化物会滞留在叶片、茎秆和果壳中[38]。上述缺钙引起症状最终都会体现为植株生长发育受阻, 产量或品质衰减, 大幅度地降低了作物的经济效益。因此充足的钙供应是作物产量与品质进一步提升的潜在突破口。
4 作物钙缺乏原因
土壤缺钙及植株生理性缺钙是植物缺钙的两大主要原因。土壤是作物的主要钙来源, 土壤中的钙分为有机钙、矿物态钙、交换态钙和水溶性钙, 其中植物可吸收利用的主要是交换态和水溶性钙, 而土壤中钙则主要以植物难吸收的矿物态钙形式存在[39]。中国土壤中的钙浓度(以土壤中CaO含量计算)总体上呈现由西北向东南递减的趋势, 这种分布状况与中国的干湿地区划分以及降水量分布高度相关[40]。就全国而言, 土壤表层(0~25 cm)钙浓度的中位值在1.96%, 其中干旱、半干旱、半湿润和湿润地区的钙浓度中位值分别为4.61%、2.73%、1.37%和0.56%[40]; 在西北等干旱地区, 钙可作为蒸发岩矿物质沉积在土壤中, 导致土壤钙含量高, 但矿物态钙占比高, 作物可吸收的钙比例低; 而在东南部湿润地区, 水溶性钙易直接溶于地下水或地表水中而流失, 一些酸性离子也可将交换态钙置换为水溶性钙, 使钙流失。土壤各形态钙占比也受到土壤pH值和营养成分的影响。不同pH值的土壤中, 作物钙肥利用效率表现为盐碱土壤 > 酸性土壤 > 中性土壤。土壤中水溶性钙与水溶性钾、水溶性镁分别表现为拮抗作用与协同作用, 而交换性钙则与交换性钾、交换性镁表现为拮抗作用与协同作用[14,41-42]。土壤中钙元素分布还受到人类活动影响, 一方面工业活动带来的酸雨会降低土壤pH, 大幅增加土壤中的硫酸根和硝酸根离子含量, 从而溶解矿物态钙, 生成水溶性钙而流失, 另一方面由于高密、高产等栽培技术的长期应用, 土壤中的钙被作物大量吸收, 但未得到及时补充[43-44]。
生理性缺钙有2个方面的原因。一方面由于钙元素的难再分配特性, 作物无法通过韧皮部实现衰老组织到年轻组织的钙元素再分配, 各组织只有依赖蒸腾作用和水分运动, 通过木质部的运输获得钙供应, 这也导致一些蒸腾作用较弱的组织(幼果、心叶等)更容易出现缺钙症状[45]。这种特性也使作物对钙的吸收受到自然天气的约束, 在阴雨时节或高温干旱时, 作物蒸腾作用都会受到影响从而限制作物对钙离子的吸收和运输[46]。另一方面K+、Mg2+和NH4+等阳离子与Ca2+存在离子拮抗关系, 会限制作物对钙的吸收和分配, K+和Mg2+既有拮抗关系又存在协同关系, 因此钾/镁、钾/钙、钙/镁、氮/钙等营养平衡状态也会影响钙的吸收利用[34,41,47]。因此, 生产中经常出现土壤并不缺少钙元素, 作物却表现出缺钙症状的现象。
土壤可吸收利用钙含量和作物对钙的吸收转运能力共同决定着作物是否缺钙, 因此针对缺钙的不同情况, 需要调整应对措施, 通过施钙、调节土壤pH、营养元素比例和水分管理保证土壤中足够的可吸收利用钙含量。特别是生理性缺钙通常发生在灌浆结实或坐果阶段, 这时通过外源喷施钙溶液可以有效防止果实开裂、避免钙缺乏症和保证产量、品质。
5 钙在作物生长发育中的角色
5.1 钙作为结构成分
5.1.1 维持细胞壁结构 植物细胞壁从外到内依次为中胶层、初生细胞壁和次生细胞壁: 中胶层处在2个相邻细胞之间, 包含大量果胶质, 能使细胞彼此粘连, 同时有一定的可塑性; 初生细胞壁位于中胶层内侧, 主要包含纤维素、半纤维素和果胶; 次生壁沉积在初生壁内侧, 是由初生壁持续生长增厚所形成的, 主要包含纤维素和少量半纤维素, 常含有木质素[48-50]。细胞壁中这些成分之间相互交联共同维持细胞形态, 为植株提供机械支撑[51]。
钙是作物维持细胞壁完整性所必需的, 决定细胞壁的刚度。作物体内的钙主要贮藏在细胞壁之中, 除游离形式的Ca2+, 钙还以负载离子的形式参与交联负电荷, 能够与去甲酯化的多聚半乳糖醛酸(homogalacturonan, HG)形成分子间Ca2+键, 加强果胶分子的交联, 赋予了细胞壁显著的结构刚性(图1),因此在缺乏钙时, 会导致细胞壁软化, 无法实现支撑功能[12,52]。例如, 钙盐处理能延迟采摘后芒果的果皮软化, 是因为细胞壁降解被抑制[53]; 利用EDTA (一种Ca2+螯合剂)能够改变芍药次生壁生物合成, 降低其花序茎的机械强度[54]; 外源钙处理延缓了非洲菊茎的弯曲, 提高乳草花花序茎的机械强度, 这可能是由于果胶分子的偶联导致细胞壁更硬[55-56]。花粉管在柱头中的伸长过程受到Ca2+浓度([Ca2+])严格限制, 其伸长容许[Ca2+]范围在10 μmol L–1到10 mmol L–1之间, 在容许[Ca2+]范围内, 花粉管能保持细胞完整性, 同时还具有一定可塑性, 如[Ca2+]在10 μmol L–1以下, 花粉管则会因为管壁太薄而在伸长过程中破裂, 而在10 mmol L–1以上, 花粉管生长也经常停止, 这可能是由于Ca2+与果胶分子过度交联[57]所致。
5.1.2 保证膜的稳定性 细胞膜是所有生物细胞共有的结构, 是细胞的边界, 是细胞与胞外信息交流和物质交换的重要场所[58]。根据1972年Singer和Nicolson提出的流动镶嵌模型学说, 细胞膜主要由磷脂双分子层和全部或部分镶嵌其中的蛋白质分子组成, 磷脂的流动性和蛋白质的运动性使细胞膜结构处于不断变动状态, 同时也是选择透过性的基础, 因此如何保证细胞膜稳定性是非常关键的。
钙的参与是保证细胞膜稳定性的重要环节, 在细胞膜中, 钙通过结合羧基实现磷脂和蛋白质的联结(图1), 同时能在膜表面结合位点处和其他阳离子进行有效交换[59]。在拟南芥根中, 钙缺乏显著地加剧了质膜内陷、核内小体的形成, 同时还发现磷脂酰胆碱(phosphatidyl choline, PC)、溶素-PC、磷脂酰乙醇胺(phosphatidyl ethanolamine, PE)、溶素-PE、磷脂酰肌醇(phosphatidylinositol, PI)和甘油三酯(triglyceride, TAG)生物合成增加, 而磷脂酸(phosphatidic acid, PA)和甘油二酯(diglyceride, DAG)生物合成减少, TAG含量和PC/PE摩尔比的增加表明缺钙可能导致膜脂组成比例失衡[60]。钙还能与膜联蛋白(annexins)结合从而促进annexins与膜脂的结合, AnxA2 (annexin A2)以钙依赖方式通过诱导脂质再分配来调节质膜特性, 这可能会导致膜的有序或无序的增加[61]。
5.2 钙作为胞内第二信使
钙不仅作为作物生长的一种必需营养元素, 它也承担着区别于其他营养元素的第二信使角色。近年来, 随着对于钙在各种生理、发育以及应激相关过程充当第二信使的研究愈发深入, 不少学者认为钙作为营养元素所带来的一些效应也可能离不开这一功能[4]。
5.2.1 钙信号的编码 细胞质内的钙浓度([Ca2+]cyt)受到严格控制, 主要是为了防止高[Ca2+]cyt带来的离子毒性, 以及保证钙信号的精准传导。当来自胞外的“第一信使” (包括激素、机械刺激、非生物和生物胁迫等)被细胞膜上的类受体蛋白激酶(receptor-like kinases, RLKs)等受体所感知, 从而激活质膜上的Ca2+通道, 通过从外部空间流入或从细胞内储存释放来介导Ca2+进入细胞质, 产生短暂、持续或间歇的钙振荡, 为了防止细胞毒性, 多余的Ca2+需通过钙泵或离子交换器从细胞质中移除(图1)[62]。Ca2+外排或内流状态的通道正是编码过程的关键, 同时也是植物吸收、转运Ca2+的关键环节, 这些通道可按照其功能分为内流系统和外排系统。
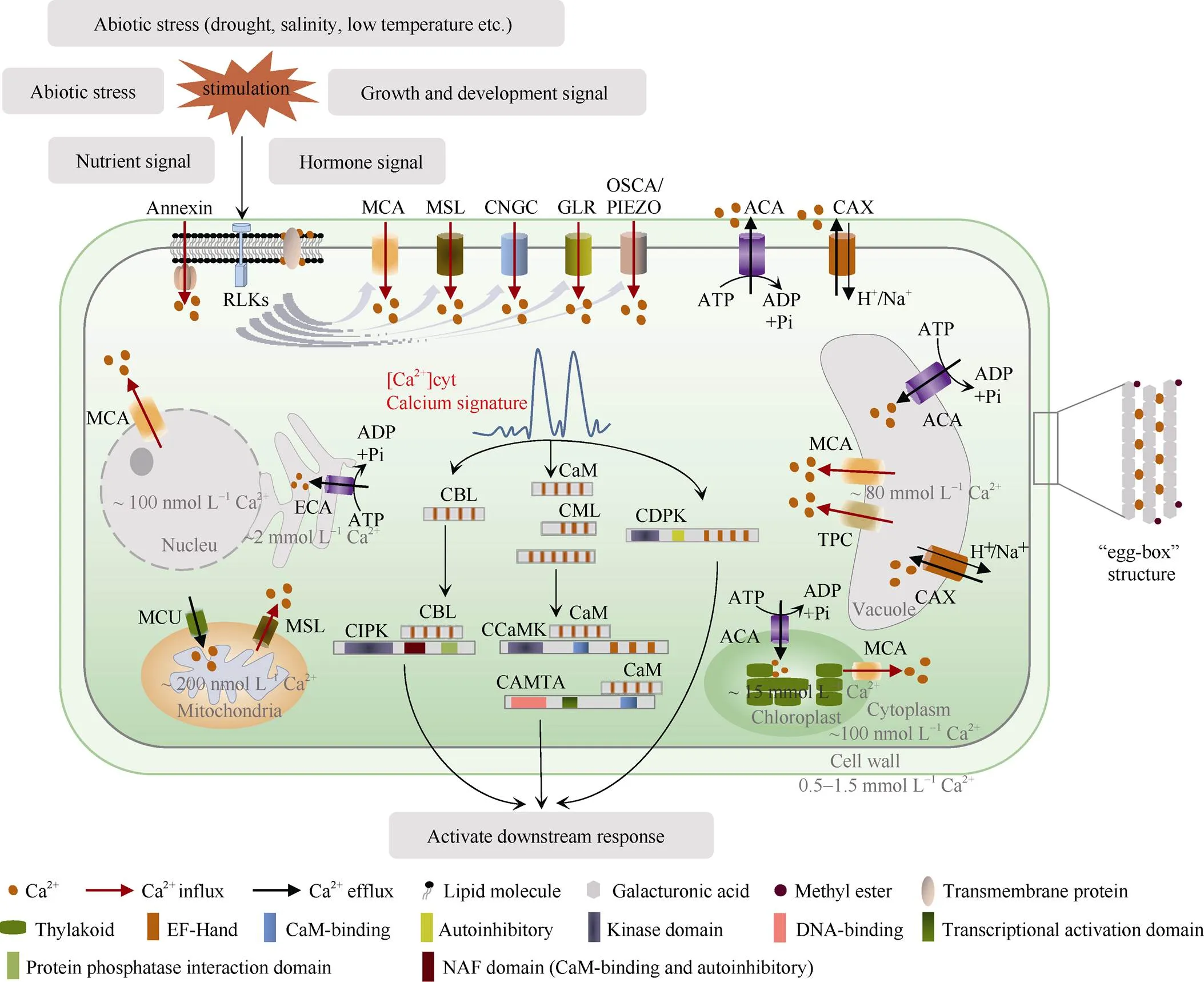
图1 钙在作物细胞中的角色、分布和运动及钙信号传导
Ca2+通过与羧基残基结合, 能够联结磷脂和膜蛋白保持细胞膜稳定, 并能交联多聚半乳糖醛酸形成“蛋盒”结构增加细胞壁强度。Ca2+在细胞壁、细胞膜、细胞质、细胞核、叶绿体、线粒体、内质网上均有分布, 细胞质中的Ca2+浓度受到严格控制, 静息状态下维持在100 nmol L–1左右, 细胞核和线粒体中的Ca2+浓度比较接近胞质Ca2+浓度, 而细胞壁、液泡和叶绿体的Ca2+浓度达到了mmol L–1级别, 远高于胞质Ca2+浓度, 是重要的Ca2+储存场所。RLKs等类受体激酶感知膜外刺激, 磷酸化激活钙离子内流系统, 包括环核苷酸门控通道(CNGCs)、谷氨酸受体样通道(GLRs)、机械敏感通道(MSLs)、机械敏感的“Mid1互补活性”通道(MCAs)、机械敏感压电通道(PIEZO)和高渗压诱导[Ca2+]cyt通道(OSCAs), Ca2+从细胞间隙流入细胞质中, 产生钙峰, 随即被Ca2+-ATPase (ACAs/ECAs)和Ca2+/阳离子反向转运蛋白(CAXs)逆浓度梯度转移至胞外, 如此形成短暂或持续的钙振荡, 此时具有EF-hands的钙调素/类钙调素(CaMs/CMLs)、钙调磷酸酶B类蛋白(CBLs)等感受器能结合钙离子, 并识别和解码产生的钙峰或钙振荡, 从而激活对应靶蛋白(CIPKs、CCaMKs、CAMTA等)的催化或转录激活活性, 进而激活下游反应以响应刺激。钙依赖蛋白激酶(CDPKs)自身同时拥有EF-hands和催化结构域, 能不依赖其他钙感受器直接结合Ca2+并激活自身激酶活性。图片根据参考文献[4, 14, 23, 52, 62, 65, 70]进行改制。
Ca2+can bind phospholipids and membrane proteins to maintain cell membrane stability by binding to carboxyl residues, and can cross-link homogalacturonan to form an “egg box” structure to strengthen cell wall. Ca2+is widely distributed in cell wall, membrane, cytoplasm, nucleus, chloroplast, mitochondria, and endoplasmic reticulum. [Ca2+]cytis strictly controlled and maintained at about 100 nmol L–1at resting state. The concentration of Ca2+in the nucleus and mitochondria is relatively close to [Ca2+]cyt. Ca2+concentration in cell wall, vacuole, and chloroplast reaches the level of mmol L–1, which is much higher than cytoplasmic Ca2+concentration, and is an important for Ca2+storage site. Receptor-like kinases (RLKs) sense extracellular stimulation and activate Ca2+influx systems by phosphorylation. Ca2+influx systems include cyclic nucleotide-gated channels (CNGCs), glutamate receptor-like channels (GLRs), mechanosensitive-like channels (MSLs), Mid1-complementary activity channels (MCAs), mechanosensitive piezoelectric channels (PIEZO), and hyperosmo-induced [Ca2+]cytchannels (OSCAs). Ca2+flows from intercellular spaces into the cytoplasm through Ca2+influx systems, producing calcium peaks that are then transferred extracellular by Ca2+-ATPase (ACAs/ECAs) and Ca2+/cationic exchangers (CAXs) against concentration gradients, resulting in transient or sustained calcium oscillations. At this time, receptors such as calmodulin/calmodulin-like (CaMs/CMLs) and calcineurin B proteins (CBLs) with EF-hands can bind Ca2+, identify, and decode the generated calcium peaks or calcium oscillations, and thus activating the catalytic or transcriptional activation activity of target proteins (CIPKs, CCaMKs, CAMTA, etc.). This in turn activates the downstream response in response to the stimulus. Calcium-dependent protein kinases (CDPKs) possess both EF-hands and catalytic domains, and can directly bind Ca2+and activate kinase activity independently of other calcium receptors. The picture is restructured according to the references [4, 14, 23, 52, 62, 65, 70].
内流系统: Ca2+流入细胞质主要依靠各种Ca2+通道, 通道的活动受到细胞内和细胞外信号的严格调节。由膜电位或电压控制的通道被称为电压门控通道(voltage-gated calcium ion channels, CaVs); 对配体分子结合敏感的被称为配体门控通道(ligand- gated calcium channels, LGCCs); 一些通道对细胞膜的机械变化敏感, 因此被称为机械敏感通道[62]。有趣的是, 研究者发现随着植物离开海洋, 在陆地上定居, CaVs类通道几乎在植物中逐渐消失了[63], 如编码具有双孔的电压门控通道的(two-pore channel 1)基因在拟南芥中只存在单个拷贝, 这种现象可能是因为陆地植物在组织和细胞水平上缺乏运动[62,64]。与之对应是一些LGCCs类型在植物中的数量之多超乎意料, 如环核苷酸门控通道(cyclic nucleotide-gated channel, CNGCs)和谷氨酸受体样通道(glutamate receptor-like proteins, GLRs), 在拟南芥基因组中至少存在20个基因家族成员和20个基因家族成员, 这2类通道都是非选择性的阳离子通道[65]。机械敏感通道主要包括机械敏感通道(mechanosensitive-like channels, MSLs)、机械敏感的“Mid1互补活性”通道(mid1-complementing activity channels, MCAs)、机械敏感压电通道(mechanosensitive piezoelectric channel, PIEZO)和高渗压诱导[Ca2+]cyt通道(hyperosmolality-induced [Ca2+]cytchannels, OSCAs),主要通过激活Ca2+内流感知触觉和光等机械的刺激[66]。此外, 还有一种能够与膜磷脂结合的Ca2+依赖性胞浆蛋白(annexins, ANN), 但ANN不具有跨膜结构域, 一些研究表明它们可能形成了氧化还原调节的Ca2+渗透孔, 但ANN通过什么机制形成通道还需继续深入研究[67-68]。另外在动物中, ANN通过Ca2+依赖的方式与膜脂结合, 参与胞吐和内吞[69]。
外排系统: Ca2+通过内流系统进入细胞质, 致使[Ca2+]cyt顿时增加数倍以响应各种胞外刺激, 但又会在短时间内恢复, 这种恢复便是由外排系统所介导的, 主要通过各种Ca2+转运蛋白逆浓度梯度将Ca2+排出胞质, 此过程消耗能量[70]。植物中Ca2+转运蛋白可分为2类: P型Ca2+-ATPase和Ca2+/H+反向转运蛋白(Ca2+/阳离子反向转运蛋白), 前者依赖于ATP的水解, 并利用这个过程中释放的能量[71], 后者则由H+-ATPase提供质子驱动力, 属于次级主动运输[72]。Ca2+-ATPase又称钙泵, 对Ca2+有较高的亲合性(m= 0.2~1.0 µmol L–1), 但运输能力较低[73]。钙泵主要包括内质网型Ca2+-ATPase (endoplasmic Ca2+-ATPase, ECA)和自抑制的质膜型Ca2+-ATPase (Ca2+-ATPase, ACA), 前者定位于内膜, 后者定位于内膜和质膜[65]。2种钙泵对于Ca2+有不同的亲和力, 分别为亚微摩尔和微摩尔级别[74]。另外ACA具有高度的选择性, 仅运输Ca2+, 而ECA除了Ca2+, 还可以介导Mn2+、Cd2+和Zn2+的运输[71,75]。与钙泵相反, Ca2+/H+反向转运蛋白是低亲合性(m约为10 µmol L–1),但运输能力强的载体, 在[Ca2+]cyt很高时发挥作用, 这些蛋白都由CAX (Ca2+/cationic exchanger)基因家族成员编码, 该家族还编码依赖K+的Na+/Ca2+反向转运蛋白[76]。
5.2.2 钙信号的解码 在[Ca2+]cyt升高后, 细胞质内一些钙感受器能够以高亲和力结合Ca2+, 对不同的钙信号进行解码并触发下游特定细胞反应[77]。有学者推测这种机制可能是早期生命形成的一种简单解毒机制, 后来演化为一种感知和解码钙信号的方式[62]。植物中钙感受器可分为3类: 钙调素/类钙调素(Ca2+-calmodulins/CaM-like proteins, CaMs/CMLs)、钙依赖蛋白激酶(Ca2+-dependent protein kinases, CDPKs)和钙调磷酸酶B类蛋白(calcineurin B-like proteins, CBLs), 这些感受器都具有结合Ca2+的EF-hands[14]。
在钙感受器感知特定钙信号后, 需要进一步调节下游靶蛋白触发特定反应以完成解码工作[78]。对于CaMs/CMLs而言, 其靶蛋白覆盖细胞功能相关的广泛领域, 如信号蛋白(激酶和磷酸酶)、CaM相互作用转录因子、细胞骨架成分、转运体和通道[66]。CDPKs作为一种钙依赖性的蛋白激酶, 既有结合Ca2+的EF-hands, 也有催化激酶结构域[9,79]。与CaMs/CMLs不同, CBLs的靶蛋白比较专一, 它能特异地与CBL互作蛋白激酶(CBL-interacting protein kinases, CIPKs)结合用于解码植物中的钙信号[80]。
5.2.3 非生物胁迫响应中的钙信号 钙信号存在于多种植物生长发育调控途径和各种生物胁迫、非生物胁迫以及机械刺激等响应过程中。植物受到的大部分非生物胁迫都能诱发细胞内的钙信号产生, 这些非生物胁迫主要包括干旱、盐碱、低温等逆境。
在植物中, 干旱胁迫通常与渗透胁迫密切相关, 一般认为干旱胁迫是由渗透胁迫引起的缺水反应, 会使一些生理指标受到影响, 包括气孔导度、蒸腾速率、光合速率以及细胞水势的降低, 为此植物也进化出一系列机制来启动干旱恢复(干旱恢复是指植物在严重干旱胁迫后重新开始生长的能力), 而钙信号是干旱恢复过程中非常重要的信号转导环节[70,81]。拟南芥的编码一种质膜钙渗透通道, 能感知渗透压的变化, 在高渗透压条件下诱导[Ca2+]cyt瞬时升高, 继而调节植株蒸腾速率和根系生长以响应干旱胁迫, 进行干旱恢复[82]。此外, 拟南芥机械敏感通道(AtMSL8)是通过在缺水诱导的渗透胁迫下调节低渗诱导的膜张力来维持花粉活力[83]。在棉花中, 过表达可增强其干旱胁迫的耐受性[84]。另外, 钙信号也能介导脱落酸(abscisic acid, ABA)信号的传导来增强水稻对渗透胁迫的耐受性[85]。
盐胁迫同样会造成渗透胁迫, 相当一部分水分胁迫的研究都聚焦在盐胁迫上, 这部分与干旱胁迫一定程度上存在交互作用, 但盐胁迫所带来的离子胁迫是有其独特性的, 离子胁迫主要是指高浓度的Na+/K+引起植物细胞离子稳态失衡, 从而导致植物生长被抑制[86]。科学家很早就发现拟南芥暴露于NaCl (0.333 mol L-1)胁迫下几秒钟内就表现出 [Ca2+]cyt的快速增加, 说明钙信号确实是参与了该胁迫反应。同样地, 如将100 mmol L-1NaCl溶液施用到拟南芥的部分根上, 几秒钟内就会触发Ca2+信号, 随后以波浪状方式扩展到整个根部和植株[87]。上述研究表明盐胁迫期间钙信号传导是相当复杂的过程, 其功能和影响并不局限于直接暴露在盐胁迫中的部位。另外, 有研究发现FERONIA (FER)受体激酶与果胶相互作用, 可以感知盐胁迫下的细胞壁损伤并诱导钙离子信号, 从而促进细胞壁完整性的恢复[88]。
低温是限制作物产量和品质的常见逆境之一[89]。研究证明低温胁迫会导致[Ca2+]cyt增加, 尽管其中机制还有待研究, 但也表明钙信号可能参与植物冷激响应过程[90]。Ca2+通道可能是潜在的冷传感器, 在拟南芥中, AtMCA1和AtMCA2被认为是感知冷胁迫的关键组分, 通过介导Ca2+内流调节细胞膜流动性和细胞骨架重组来应对寒冷[91]。环核苷酸门控通道OsCNGC9能触发[Ca2+]cyt升高增强水稻的耐冷性[92]。AtANN1能与OST1/SnRK2.6 (open stomata 1/Snf1- related kinase 2.6)互作并被磷酸化, 从而增强Ca2+转运活性, 继而与冷诱导的钙信号形成级联反应, 这激活了拟南芥冷激反应[93]。作为钙信号传导的中继站, 钙感受器同样参与了植物冷激响应过程,突变体植物表现为冷反应基因(cold-regulated,)转录水平升高, 并增强了对冷胁迫的耐受性, 说明AtCaM4负向调节拟南芥耐冷性[94]。突变体表现出更高的冷冻敏感性, 而的过度表达则导致耐冻表型并诱导了基因的表达[90]。甘蓝型油菜和白菜在冷处理6 h后,表达量显著增加[95]。冷激反应激活了充当冷传感器的Ca2+通道, 导致[Ca2+]cyt升高, 进而激活下游转录因子(transcription factors, TFs), 使作物产生特定的反应来对抗低温胁迫, 钙调蛋白结合转录激活因子(calmodulin-binding transcription activators, CAMTAs)就是其中一种TF, 它以Ca2+依赖性方式参与冷激反应过程, 并且受到CaM调节[96-97]。Kidokoro等[98]发现CAMTA3和CAMTA5能对温度的快速下降做出反应并诱导(一种冷响应基因)的表达。
6 钙在作物中的应用
6.1 钙对光合作用的影响
光合作用是绿色植物甚至是地球上绝大部分生物的基础能量来源, 对于作物的重要性不言而喻。作物光合器官的光合能力决定了作物光合同化物的积累能力, 也决定了作物最终的生物量和产量(图2)。光合作用主要发生在高等植物的叶绿体中, 而叶绿体就是植物细胞内的钙库之一, 也存在许多钙转运蛋白和钙结合蛋白, 这也表明光合作用过程很可能受到钙调节[99]。陈向明等[100]发现钙浓度为160 mg L–1的营养液能显著增加牡丹叶片的气孔导度从而提高光合作用。李敏等[101]也发现160 mg L–1的钙溶液能最有效地提升牡丹光合强度和光合有机物积累能力。叶面喷施外源原己二酮钙(Pro-Ca)提高大豆叶片叶绿素含量、光合速率(photosynthetic rate,n)、电子传递速率(electron transport rate, ETR)、PSII潜在活性(potential activity of PSII,v/o)和PSII的最大量子产率(maximum quantum yield of PSII,v/m), 并延缓类囊体的降解[102]。增施钙肥后, 春玉米的叶面积指数(leaf area index, LAI)、n、光能利用率、叶片叶绿素的含量得到显著提升[103]。Ca2+作为第二信使也参与调控植物光合作用[99]。位于叶绿体类囊体膜上的钙感受器(Ca2+-sensing receptor, CAS)通过响应[Ca2+]cyt升高参与调节气孔关闭, 且外源钙处理可以解除突变后对气孔关闭的限制作用[104]。卡尔文循环是碳同化的主要过程, 主要发生在叶绿体基质中, 碳同化过程中的景天庚酮糖-1,7-二磷酸酶(sedoheptanose-1,7-diphosphatase, SBPase)和果糖- 1,6-二磷酸酶(fructose-1,6-diphosphatase, FBPase)活性受到Ca2+调节, 但高浓度的Ca2+处理会抑制碳同化过程[105]。外源性Ca2+可以在热应激期间激活抗坏血酸过氧化物酶(ascorbic acid peroxidase, APX)、过氧化氢酶(catalase, CAT)和超氧化物歧化酶(superoxide dismutase, SOD)从而调节活性氧(reactive oxygen species, ROS)平衡和保护光系统[106]。Ca2+还通过调节微管运动系统和微丝运动系统介导蓝光下的叶绿体重组活动, 使得作物细胞的叶绿体排列方式能适应不同光照条件[107]。
6.2 钙对作物产量和品质的影响
研究表明, 大豆开花时叶面喷施钙肥可以提高叶片光合作用, 增加叶片中的可溶性糖和蛋白质浓度, 代谢活性的提高使植株获得更高的产量潜力, 同时钙的施用在后期可以促进花的保留和荚果的固定, 从而增加每株的荚果数量, 进而提高作物生产力[108]。钙镁磷肥能改善土壤pH值和养分有效性, 进而提高香蕉生物量和产量, 同时可以增加香蕉可溶性固体物和可溶性糖含量[109]。用不同物质在播种前对小麦种子进行预处理, 发现硝酸钙(1 mmol L–1)处理可以显著提高地上部干重、单株粒数、千粒重、单株穗数和叶绿素含量[110]。Kanu等[111]发现在土壤中施用5 mg kg–1钙能增加水稻的产量及构成因素(穗数、每穗粒数、结实率、千粒重)。外源Ca2+(5 mmol L–1)处理显著提升了酸雨胁迫下的水稻产量和籽粒品质(氨基酸和蛋白质含量)[112]。Metwally等[113]报道了CaCl2、Na2SiO3和水杨酸(salicylic acid, SA)的添加均能增强非酶抗氧化剂活性和水分含量从而改善油菜生长状况, 并提高单株角果数量、角果干重和千粒重。
6.3 钙能提高作物抗逆性
钙离子不仅在逆境胁迫响应中承担重要的信使作用, 还在调控作物抗逆性中被广泛应用(图2)。生理指标和转录组分析表明, 外源喷施钙能增加可溶性蛋白、可溶性糖和脯氨酸含量以及吲哚乙酸(indoleacetic acid, IAA)、油菜素内酯(brassinosteroid, BR)和乙烯(ethylene, Eth)等内源性激素含量, 并促进光系统-亚基PsbP同源物1 (PsbP-like protein 1,)、光系统-反应中心Psb28蛋白(photosystem- reaction center Psb28 protein,)、光系统II亚基S4 (non-photochemical quenching 4,)等光合作用相关基因的表达, 从而减轻弱光对马铃薯植株的伤害[114]。外源施加Ca2+能改变盐胁迫下玉米叶片气孔结构, 显著降低气孔导度, 提升叶片n, 增加幼苗生物量, 降低根冠比, 表明钙处理对盐胁迫有缓解作用[115]。在模拟酸雨胁迫下, 外源施加Ca2+可以维持大豆对硝态氮、铵态氮和磷的吸收, 增强了大豆的适应能力, 促进大豆产量和籽粒品质的恢复[116]。100 μmol L–1SA和5 mmol L–1CaCl2混合处理能有效改善鼠尾草应对热胁迫的能力[117]。大豆在移栽前10 d和移栽后14 d用10 mmol L–1Ca(HCOO)2-A (Suicaru)溶液处理能减少田间疫霉茎腐病发病率[118]。

图2 施钙对作物生长发育中的作用
6.4 钙调节作物重金属吸收
我国农业土壤重金属污染程度逐步加剧, 污染的面积也在扩大, 农作物重金属污染问题逐渐显露[119]。通过添加外源钙, 镉胁迫作物中被抑制的总酚、黄酮和矿物质元素含量得以缓解[120]。Ca2+以剂量依赖性方式调节作物对镉离子(Cd2+)的吸收和转运, 较低剂量的Ca2+促进作物对Cd2+的吸收和转运, 但较高剂量会降低根部对Cd2+的吸收[121]。随着水稻植株中钙浓度的增加, 镉在整个植株中的积累量显著减少, 但地上部的Cd2+含量却显著增加, 说明Ca2+减少了根系对Cd2+的吸收, 但刺激了Cd2+从根部到地上部的转运, 这种相反的结果表明Ca2+在调节水稻根系Cd2+吸收和根系到地上部Cd2+转运方面具有相反的作用[122]。氧化钙纳米颗粒(CaO NPs)的施用能够增强大麦活性氧清除能力, 减少根系砷吸收和向地上部的转运[123]。施用钙多肽能提高水稻根部的铅含量, 同时降低茎、叶和糙米中的铅含量[120]。另外, Ca2+还通过CDPKs和CBL-CIPKs信号转导途径介导有毒离子的吸收, 如Ji等[124]报道CPK31可调节非必需和有毒亚砷酸盐(As3+)的摄取。钙与其他金属离子存在潜在的协同或拮抗作用, 因此通过钙肥的合理施用来治理农作物重金属污染是可行的(图2)。
7 讨论与展望
钙是作物所必需的营养元素, 对作物生长发育具有重要意义。本文阐述了作物体内的钙分布和转运情况以及作物的钙需求特性和缺钙症状, 同时从结构成分和信使双重功能角度介绍了钙在作物中的角色和作用, 并综述了钙在作物生产的应用。
钙的施用已经被证明能增加作物产量、提升品质、增强抗逆性。由于土壤pH值和营养元素结构的影响, 特别是氮肥的大量施用, 导致N/Ca严重失调, 使得钙肥的吸收利用效率并不高, 造成了作物常出现缺钙情况[34,41-42,47]。研究表明, 均衡的养分比例更有利于钙素积累, 如尤召阳等[125]通过氮钙复配试验,发现600 kg hm–2Ca和225 kg hm–2N的组合显著增加花生氮素利用率和钙素积累。事实上, 如何保证作物根系吸收充足的钙元素是实现钙对作物生长发育发挥积极调控作用的首要环节。同时钙作为一种移动性较差的元素, 作物生长后期也经常因此出现生理性缺钙现象, 尤其是结实阶段, 茎、叶等营养器官向果实、籽粒的钙转运效率有限, 最终导致果实籽粒缺钙, 影响产量和品质。有研究证明IAA、萘乙酸(naphthalene acetic acid, NAA)、赤霉素(gibberellic acid, GA)等激素能够促进钙吸收和钙向果实的运输, 例如杜英俊等[126]利用20 mg L–1IAA和20 mg L–1GA3与钙肥混合喷施, 能显著提高补钙效果, 促进苹果的果重增加和品质提升, 但利用激素进行调节并不符合民众心理预期。合理的N/Ca施肥比也能增加钙素分配效率, 比如尤召阳等[125]发现在同一钙肥水平下, 225 kg hm–2的施氮量相较其他氮水平更能增加花生荚果的钙素分配率。因此如何提高作物钙素积累量和在作物体内的钙素转运效率是进一步发挥钙的作物生长发育调控作用的重点之一。
生产上通常利用根外喷施来进行针对性的钙补充, 目前喷施所用钙肥主要有无机钙盐和有机螯合钙。无机钙盐主要包括硝酸钙、氯化钙、过磷酸钙等, 硝酸钙的肥效相对更高, 但总体上无机钙盐的钙吸收效率要低于有机螯合钙; 有机螯合钙是钙与糖醇、氨基酸和有机酸等有机分子结合的螯合物, 是目前农业生产上的主流[127-130]。以糖醇钙为代表, 结合在糖醇上的钙元素会被作物的韧皮部识别成糖醇进行吸收和运输, 而不易被果胶和草酸等有机酸截获, 糖醇本身具有保湿功能, 也是一种表面活性剂, 可以延长钙在叶片上的停留时间并使液体均匀覆盖叶片或果实。但研究发现由于表面蜡质等原因, 叶面或果面喷施的钙吸收效率也很有限[11,14]。随着纳米技术的兴起, 人们开始研究纳米钙、天冬氨酸纳米钙(Ca (L-asp-NPs))、纳米磷酸钙(NCaP)等纳米肥料, 发现纳米肥料相比传统化肥的利用效率更高, 例如在菜豆的研究中发现, NCaP仅需过磷酸钙传统推荐用量(30 kg P2O5)的10%就能达到相同的补钙效果[131]。但纳米材料的使用多限于研究领域, 并未进一步应用推广到实际生产中。因此筛选出高效吸收、使用成本低的外源钙物质也是改善作物生理性缺钙和保证作物产量、品质的重要措施。
作为具有双重功能的特殊营养元素, 钙在调控作物生长发育方面发挥重要作用, 然而最近的研究更多集中在其信使功能上, 关于钙的营养功能和信使功能之间的联系却少有人探究。比如钙的外源处理能够加强果胶交联从而保证根系伸长区细胞在盐胁迫下的细胞壁完整性, 而FER能感知盐胁迫下的细胞壁损伤从而诱发钙信号的产生促使细胞壁恢复完整性[88], 是否钙外源处理也会诱发类似的钙信号产生, 而不仅仅是钙的结构效应, 具体机制还需后续深入研究。Weinl等[104]发现用5 mmol L–1的Ca2+溶液处理拟南芥后30 s内叶片保卫细胞中出现强烈的钙峰, 随即转变为持续的钙振荡。说明外源钙处理确实能够诱导钙振荡, 但这种“钙信号”是否能被正确解码, 是否能与最终钙处理的效应相关联值得进一步探究。
另外, 作物体内的钙多聚集在细胞壁间隙以及细胞壁中, 而细胞内的液泡以及叶绿体、内质网等细胞器也是重要的钙库[23](图1)。液泡中的钙主要形态是草酸钙和磷酸钙, 通常被认为是作物细胞内的钙储存库, 而液泡中游离的Ca2+浓度也能达到0.2~ 5.0 mmol L–1, 有研究表明液泡中的Ca2+也参与钙信号形成, 许多钙离子转运蛋白被定位在液泡膜上, 包括CAX2和TPC1, 其中CAX2被认为能够调节胞质Ca2+信号的持续时间和幅度从而参与拟南芥的渍水响应[132], 但并不能证明液泡中的Ca2+直接参与钙信号的形成, 这可能因为CAX2介导的Ca2+流方向是从胞质到液泡, 属于外排系统; TPC1已经被证明定位在液泡膜上, 是主要的液泡Ca2+释放通道, 属于内流系统, TPC1是否参与钙信号形成一直受到争议,突变体在各种逆境胁迫下, 与野生型的[Ca2+]cyt变化没有差异[133], 但Choi等[87]发现TPC1可能参与调节植物抵抗逆境形成的长距离钙信号传递速度。叶绿体中Ca2+浓度虽然可以达到15 mmol L–1,但大多是与蛋白结合, 而叶绿体基质中的游离Ca2+浓度却与[Ca2+]cyt相近, 只有约150 nmol L–1 [23]; CAS被定位在类蘘体膜上, 是一类钙感受器, 具有EF-hands, CAS可能通过接收外源钙信号, 将被结合的Ca2+释放出来, 然后被叶绿体膜上的钙离子通道转移至胞质中, 参与[Ca2+]cyt的升高从而调控气孔开闭[104], 但具体的调节机制尚未清楚。细胞器不仅是重要的钙储存库, 也在钙信号形成和传导方面具有重要作用, 但具体如何发挥功能还不清晰, 需要进一深入研究。
总而言之, 钙作为一种特殊的营养元素, 其在作物中承担的功能是非常重要的, 也是非常复杂的, 未来仍需要继续探究和明确钙在作物生长发育中的作用, 才能更好地利用钙和巧用钙。
[1] 白由路. 中国农业科学院植物营养与肥料研究60年. 植物营养与肥料学报, 2017, 23: 1409–1415. Bai Y L. History of plant nutrition and fertilizer research in China., 2017, 23: 1409–1415 (in Chinese with English abstract).
[2] 胡铁军, 张怀杰, 郑佩君, 周飞, 魏杰. 硅钙磷镁钾肥对小麦扬麦20经济性状和土壤理化性状的影响. 浙江农业科学, 2020, 61(1): 15–16. Hu T J, Zhang H J, Zheng P J, Zhou F, Wei J. Effects of silicon, calcium, magnesium, phosphorus and potassium fertilizer on economic characters of wheat Yangmai 20 and soil physical and chemical properties., 2020, 61(1): 15–16 (in Chinese with English abstract).
[3] 王建国, 张佳蕾, 郭峰, 唐朝辉, 杨莎, 彭振英, 孟静静, 崔利,李新国, 万书波. 钙与氮肥互作对花生干物质和氮素积累分配及产量的影响. 作物学报, 2021, 47: 1666–1679. Wang J G, Zhang J L, Guo F, Tang C H, Yang S, Peng Z Y, Meng J J, Cui L, Li X G, Wan S B. Effects of interaction between calcium and nitrogen fertilizers on dry matter, nitrogen accumulation and distribution and yield of peanut., 2021, 47: 1666–1679 (in Chinese with English abstract).
[4] Thor K. Calcium-nutrient and messenger., 2019, 10: 440.
[5] 陈德伟, 汤寓涵, 石文波, 张夏燕, 陶俊, 赵大球. 钙调控植物生长发育的进展分析. 分子植物育种, 2019, 17: 3593–3601. Chen D W, Tang Y H, Shi W B, Zhang X Y, Tao J, Zhao D Q. Progress in the regulation of calcium growth and development., 2019, 17: 3593–3601 (in Chinese with English abstract).
[6] Barberon M, Vermeer J E, De Bellis D, Wang P, Naseer S, Andersen T G, Humbel B M, Nawrath C, Takano J, Salt D E, Geldner N. Adaptation of root function by vutrient-induced plasticity of endodermal differentiation., 2016, 164: 447–459.
[7] White P J. The pathways of calcium movement to the xylem., 2001, 52: 891–899.
[8] McAinsh M R, Webb A, Taylor J E, Hetherington A M. Stimulus-induced oscillations in guard cell cytosolic free calcium., 1995, 7: 1207–1219.
[9] White P J, Broadley M R. Calcium in plants., 2003, 92: 487–511.
[10] 宋雯佩. 果实摄取钙的规律、途径及调控机理的研究. 华南农业大学博士学位论文, 广东广州, 2018. Song W P. The Study of Fruit Calcium Uptake Pattern, Pathways and Regulatory Mechanisms. PhD Dissertation of South China Agricultural University, Guangzhou, Guangdong, China, 2018 (in Chinese with English abstract).
[11] Hocking B, Tyerman S D, Burton R A, Gilliham M. Fruit calcium: transport and physiology., 2016, 7: 569.
[12] Hepler P K, Winship L J. Calcium at the cell wall-cytoplast interface., 2010, 52: 147–160.
[13] 徐平宜. 中国北方不同类群植物草酸钙特征研究. 南开大学硕士学位论文, 天津, 2021. Xu P Y. The Characteristics of Calcium Oxalate in Different Groups of Plants in Northern China. MS Thesis of Nankai University, Tianjin, China, 2021 (in Chinese with English abstract).
[14] 周卫, 汪洪. 植物钙吸收、转运及代谢的生理和分子机制. 植物学通报, 2007, 24: 762–778. Zhou W, Wang H. The physiological and molecular mechanisms of Calcium uptake, transport and metabolism in plants., 2007, 24: 762–778 (in Chinese with English abstract).
[15] Hetherington A M, Brownlee C. The generation of Ca2+signals in plants., 2004, 55: 401–427.
[16] 王守银, 张宁, 樊兆博, 宋涛, 刘志涛, 桑净净, 邢璐, 栾好安,彭云. 不同形态钙对设施黄瓜生长及钙吸收的影响. 安徽农业科学, 2015, 43(34): 199–201. Wang S Y, Zhang N, Fan Z B, Song T, Liu Z T, Sang J J, Xing L, Luan H A, Peng Y. Effects of different calcium forms on growth and assimilation of calcium of cucumber., 2015, 43(34): 199–201 (in Chinese with English abstract).
[17] 张鹏飞, 王立峰, 刘倩倩, 张小娟, 凌冬. 植物中Ca2+生理功能的研究进展. 中南农业科技, 2023, 44(5): 227–232. Zhang P F, Wang L F, Liu Q Q, Zhang X J, Ling D. Advances in physiological functions of Ca2+in plants., 2023, 44(5): 227–232 (in Chinese).
[18] 王学武. 水稻钙积累分布规律与调控研究. 湖南农业大学博士学位论文, 湖南长沙, 2007. Wang X W. Studies on the Characters of Calcium Accumulation and Distribution of Rice and Its Regulation. PhD Dissertation of Hunan Agricultural University, Changsha, Hunan, China, 2007 (in Chinese with English abstract).
[19] 徐静静, 慈华聪, 何兴东, 薛苹苹, 赵雪莱, 郭健潭, 高玉葆. 天津盐渍化生境54种植物钙晶体与钙组分特征. 应用生态学报, 2012, 23: 1247–1253. Xu J J, Ci H C, He X D, Xue P P, Zhao X L, Guo J T, Gao Y B. Features of calcium crystals and calcium components in 54 plant species in salinized habitats of Tianjin., 2012, 23: 1247–1253 (in Chinese with English abstract).
[20] 杨利玲, 张桂兰. 土壤中的钙化学与植物的钙营养. 甘肃农业, 2006, (10): 272–273. Yang L L, Zhang G L. Calcium chemistry in soil and calcium nutrition in plants., 2006, (10): 272–273 (in Chinese).
[21] 刘晓伟, 鲁剑巍, 李小坤, 卜容燕, 刘波. 直播冬油菜钙、镁、硫养分吸收规律. 中国油料作物学报, 2012, 34: 638–644. Liu X W, Lu J W, Li X K, Bu R Y, Liu B. Absorption of calcium, magnesium and sulfur by winter rapeseed () under direct-seeding cropping system., 2012, 34: 638–644 (in Chinese with English abstract).
[22] 龚明, 李英, 曹宗巽. 植物体内的钙信使系统. 植物学通报, 1990, (3): 19–29. Gong M, Li Y, Cao Z X. Calcium messenger system in plants., 1990, (3): 19–29 (in Chinese).
[23] 郑远, 陈兆进. 植物细胞器钙信号研究进展. 植物生理学报, 2015, 51: 1195–1203. Zheng Y, Chen Z J. Organellar calcium signaling in plants., 2015, 51: 1195–1203 (in Chinese with English abstract).
[24] 邹娟, 鲁剑巍, 吴江生, 李银水. 4个双低甘蓝型油菜品种钙、镁、硫吸收动态. 华中农业大学学报, 2009, 28: 295–299. Zou J, Lu J W, Wu J S, Li Y S. Uptake dynamics of calcium, magnesium and sulfur in four double lowvarieties., 2009, 28: 295–299 (in Chinese with English abstract).
[25] 刘珂珂, 于宏, 高华鑫, 郭峰, 张佳蕾, 王建国, 万书波. 施钙对酸性土花生钙素吸收与积累的影响. 中国油料作物学报, 2023 [2023-10-16]. https://kns.cnki.net/kcms/detail/42.1429.s. 20230327.1134.001.html. Liu K K, Yu H, Gao H X, Gao F, Zhang J L, Wang J G, Wan S B. Effects of Ca fertilizer application on Ca absorption and accumulation of peanut in acid soil., 2023 [2023-10-16] (in Chinese with English abstract). https://kns.cnki.net/kcms/ detail/42.1429.s.20230327.1134.001.html.
[26] 马文博. 高钙酵素水溶肥制备及对蔬菜钙含量和产量影响. 东北农业大学硕士学位论文, 黑龙江哈尔滨, 2022. Ma W B. Preparation of High Calcium Enzyme Water-soluble Fertilizer and Its Effect on Vegetable Calcium Content and Yield. MS Thesis of Northeast Agricultural University, Harbin, Heilongjiang, China, 2022 (in Chinese with English abstract).
[27] 林葆, 周卫. 棕壤中花生钙素营养的化学诊断与施钙量问题的探讨. 土壤通报, 1997, 28(3): 127–130. Lin B, Zhou W. Chemical diagnosis of peanut calcin nutrition in brown soil and discussion on the amount of calcium application., 1997, 28(3): 127–130 (in Chinese).
[28] 李兆林, 才卓伟. 钙素营养对大豆的影响研究概述. 农业系统科学与综合研究, 2009, 25: 487–489. Li Z L, Cai Z W. A review of the influence of calcium on soybean., 2009, 25: 487–489 (in Chinese with English abstract).
[29] 刘永菁, 周淑清. 番茄、秋白菜钙营养的研究. 辽宁农业科学, 1988, (5): 22–25.Liu Y J, Zhou S Q. Studies on calcium nutrition of tomato and Chinese cabbage., 1988, (5): 22–25 (in Chinese).
[30] 韩配配. 不同营养元素缺乏对甘蓝型油菜营养生长及根系生长相关基因表达的影响. 中国农业科学院硕士学位论文, 北京, 2016. Han P P. The Effects of Different Nutrient Deficiencies on Vegetative Growth and the Expressions of Root Growth-related Genes in Rapeseed Seedlings (L.). MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2016 (in Chinese with English abstract).
[31] 王一斐, 周江明, 黄俊峰. 硼镁钙肥对油菜生长及产量的影响. 中国农技推广, 2019, 35(10): 83–85. Wang Y F, Zhou J M, Huang J F. Effects of borax, magnesium and calcium fertilizer on growth and yield of rape., 2019, 35(10): 83–85 (in Chinese).
[32] 李树学. 水稻缺素防治技术浅析. 农民致富之友, 2017, (24): 139. Li S X. Analysis of prevention and control technology of rice deficiency., 2017, (24): 139 (in Chinese).
[33] 俞玉. 玉米补钙防止“牛尾巴”. 农家之友, 2016, (9): 56. Yu Y. Corn calcium supplements to prevent “cow tail”., 2016, (9): 56 (in Chinese).
[34] Wang Y, Martins L B, Sermons S, Balint-Kurti P. Genetic and physiological characterization of a calcium deficiency phenotype in maize., 2020, 10: 1963–1970.
[35] 李晓彤, 杨婉莹, 孙莎莎, 巩彪, 史庆华. 外源褪黑素对番茄缺钙胁迫的缓解效应. 植物生理学报, 2019, 55: 169–176. Li X T, Yang W Y, Sun S S, Gong B, Shi Q H. Effect of exogenous melatonin on alleviating calcium deficiency stress in tomato., 2019, 55: 169–176 (in Chinese with English abstract).
[36] 徐谦, 李登云, 李配, 修明霞, 姜兴盛. 果树巧施钙肥提质增效. 果树资源学报, 2021, 2(3): 81–83. Xu Q, Li D Y, Li P, Xiu M X, Jiang X S. Calcium fertilizer of fruit trees is used to improve fruit quality and efficiency., 2021, 2(3): 81–83 (in Chinese with English abstract).
[37] 刘银发, 戴熙燕. 合理施肥预防油菜生理性缺钙. 江西农业, 2015, (11): 64. Liu Y F, Dai X Y. Rational fertilization prevents physiological calcium deficiency in rapeseed., 2015, (11): 64 (in Chinese).
[38] 李新国, 万书波. 钙对花生生长发育调控的研究进展. 山东农业科学, 2011, 43(8): 65–67. Li X G, Wan S B. Research progress on regulation of calcium to growth and development of peanut (L.)., 2011, 43(8): 65–67 (in Chinese with English abstract).
[39] 贾立国, 李利, 秦永林, 樊明寿. 马铃薯钙素营养研究进展. 北方农业学报, 2018, 46(1): 72–75. Jia L G, Li L, Qin Y L, Fan M S. Research progress of calcium nutrition in potato., 2018, 46(1): 72–75 (in Chinese with English abstract).
[40] Wang X Q, Liu X M, Wang W. National-scale distribution and its influence factors of calcium concentrations in Chinese soils from the China Global Baselines project., 2022, 233: 106907.
[41] 刘芳. 钾镁互作对番茄生长、产量及钾、钙、镁养分吸收的影响. 宁夏大学硕士学位论文, 宁夏银川, 2022.Liu F. Effects of K-Mg Interaction on Growth, Yield and K, Ca and Mg Uptake in Tamato. MS Thesis of Ningxia University, Yinchuan, Ningxia, China, 2022 (in Chinese with English abstract).
[42] 路亚. 施钙对山东花生土壤特性及花生生长发育的调控机制. 湖南农业大学博士学位论文, 湖南长沙, 2022. Lu Y. Effects of Calcium Application on Soil Propertiesand Growth and Development of Peanut in Shandong Province. PhD Dissertation of Hunan Agricultural University, Changsha, Hunan, China, 2022 (in Chinese with English abstract).
[43] 林锋. 琯溪蜜柚果园钙、镁、硫营养状况及缺镁矫治措施研究. 福建农林大学硕士学位论文, 福建福州, 2013. Lin F. The Research on Calcium Magnesium Sulfur Nutritional Status and Correctional Measures about Lack of Magnesium about Guanxi Honey Pomelo. MS Thesis of Fujian Agriculture and Forestry University, Fuzhou, Fujian, China, 2013 (in Chinese with English abstract).
[44] 鲁剑巍. 湖北省柑橘园土壤: 植物养分状况与柑橘平衡施肥技术研究. 华中农业大学博士学位论文, 湖北武汉, 2003. Lu J W. Study on Soil and Plant Nutrition Status and Balanced Fertilization Techniques of the Citrus Orchards in Hubei. PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei, China, 2003 (in Chinese with English abstract).
[45] Maclin D, Donald T S, Allen L R, Matthew G. Calcium storage in plants and the implications for calcium biofortification., 2010, 247: 215–231.
[46] Liang M, Zhang S. Why are crops prone to show symptoms of calcium deficiency in abnormal weather., 2020, 1: 17–23.
[47] 马建梅. 施用钾肥对土壤中钙、镁有效性及其效应的影响. 宁夏大学硕士学位论文, 宁夏银川, 2021. Ma J M. Effects of Different Potassium Levels on Availability of Calcium and Magnesium in the Soil and Effection. MS Thesis of Ningxia University, Yinchuan, Ningxia, China, 2021 (in Chinese with English abstract).
[48] Li S, Bashline L, Lei L, Gu Y. Cellulose synthesis and its regulation., 2014, 12: e0169.
[49] Lampugnani E R, Khan G A, Somssich M, Persson S. Building a plant cell wall at a glance., 2018, 131: jcs207373.
[50] 王艳婷. 植物细胞壁果胶结构特性与木质纤维素高效酶解产糖分子机理的研究. 华中农业大学博士学位论文, 湖北武汉, 2018.Wang Y T. Characterization of Pectin Features that Distinctively Affect Lignocellulose Enzymatic Saccharification in Plants. PhD Dissertation of Huazhong Agricultural University, Wuhan, Hubei, China, 2018 (in Chinese with English abstract).
[51] Bischoff V, Nita S, Neumetzler L, Schindelasch D, Urbain A, Eshed R, Persson S, Delmer D, Scheible W R. TRICHOME BIREFRINGENCE and its homolog AT5G01360 encode plant-specific DUF231 proteins required for cellulose biosynthesis in., 2010, 153: 590–602.
[52] Hepler P K. Calcium: a central regulator of plant growth and development., 2005, 17: 2142–2155.
[53] Díaz-Corona D A, López-López M E, Ayón-Reyna L E, López-Velázquez J G, López-Zazueta B A, Vega-García M O. Impact of hot water-calcium on the activity of cell wall degrading and antioxidant system enzymes in mango stored at chilling temperature., 2020, 44: e13286.
[54] Tang Y H, Zhao D Q, Meng J S, Tao J. EGTA reduces the inflorescence stem mechanical strength of herbaceous peony by modifying secondary wall biosynthesis., 2019, 6: 36.
[55] Li C, Tao J, Zhao D Q, You C, Ge J. Effect of calcium sprays on mechanical strength and cell wall fractions of herbaceous peony (Pall.) inflorescence stems., 2012, 13: 4704–4713.
[56] Perik R R J, Razé D, Ferrante A, Doorn W G V. Stem bending in cutflowers: effects of a pulse treatment with sucrose and calcium ions., 2014, 98: 7–13.
[57] Bosch M, Cheung A Y, Hepler P K. Pectin methylesterase, a regulator of pollen tube growth., 2005, 138: 1334–1346.
[58] Benga G, Holmes R P. Interactions between components in biological membranes and their implications for membrane function., 1984, 43: 195–257.
[59] 王云堂. 钙在植物营养中的作用分析. 种子科技, 2019, 37(6): 32–33. Wang Y T. Analysis on the role of calcium in plant nutrition., 2019, 37(6): 32–33 (in Chinese).
[60] Zhang T, Yang J, Sun Y W, Kang Y, Yang J, Qi Z. Calcium deprivation enhances non-selective fluid-phase endocytosis and modifies membrane lipid profiles inroots., 2018, 226: 22–30.
[61] Varyukhina S, Lamazière A, Delaunay J L, De Wreede A, Ayala-Sanmartin J. The Ca2+and phospholipid-binding protein Annexin A2 is able to increase and decrease plasma membrane order., 2022, 1864: 183810.
[62] Luan S, Wang C. Calcium signaling mechanisms across kingdoms., 2021, 37: 311–340.
[63] Mochida S. Presynaptic calcium channels., 2019, 20: 2217.
[64] Peiter E, Maathuis F J, Mills L N, Knight H, Pelloux J, Hetherington A M, Sanders D. The vacuolar Ca2+-activated channel TPC1 regulates germination and stomatal movement., 2005, 434: 404–408.
[65] Demidchik V, Shabala S, Isayenkov S, Cuin T A, Pottosin I. Calcium transport across plant membranes: mechanisms and functions., 2018, 220: 49–69.
[66] Hamilton E S, Schlegel A M, Haswell E S. United in diversity: mechanosensitive ion channels in plants., 2015, 66: 113–137.
[67] Laohavisit A, Davies J M. Annexins., 2011, 189: 40–53.
[68] Laohavisit A, Richards S L, Shabala L, Chen C, Colaço R D, Swarbreck S M, Shaw E, Dark A, Shabala S, Shang Z, Davies J M. Salinity-induced calcium signaling and root adaptation inrequire the calcium regulatory protein annexin1., 2013, 163: 253–262.
[69] Gerke V, Creutz C E, Moss S E. Annexins: linking Ca2+signalling to membrane dynamics., 2005, 6: 449–461.
[70] Tong T, Li Q, Jiang W, Chen G, Xue D W, Deng F L, Zeng F R, Chen Z H. Molecular evolution of calcium signaling and transport in plant adaptation to abiotic stress., 2021, 22: 12308.
[71] Huda K M, Banu M S, Tuteja R, Tuteja N. Global calcium transducer P-type Ca²⁺-ATPases open new avenues for agriculture by regulating stress signalling., 2013, 64: 3099–3109.
[72] Pittman J K, Hirschi K D. CAX-ing a wide net: Cation/H+transporters in metal remediation and abiotic stress signalling.(Stuttg), 2016, 18: 741–749.
[73] Tidow H, Poulsen L R, Andreeva A, Knudsen M, Hein K L, Wiuf C, Palmgren M G, Nissen P. A bimodular mechanism of calcium control in eukaryotes., 2012, 491: 468–472.
[74] Meneghelli S, Luoni L, De Michelis M I. Heparin stimulates a plasma membrane Ca2+-ATPase of., 2008, 143: 253–259.
[75] Bonza M C, De Michelis M I. The plant Ca2+-ATPase repertoire: biochemical features and physiological functions.(Stuttg), 2011, 13: 421–430.
[76] Yadav A K, Shankar A, Jha S K, Kanwar P, Pandey A, Pandey G K. A rice tonoplastic calcium exchanger, OsCCX2 mediates Ca2+/cation transport in yeast., 2015, 5: 17117.
[77] Thor K, Peiter E. Cytosolic calcium signals elicited by the pathogen-associated molecular pattern flg22 in stomatal guard cells are of an oscillatory nature., 2014, 204: 873–881.
[78] 郑仲仲, 沈金秋, 潘伟槐, 潘建伟. 植物钙感受器及其介导的逆境信号途径. 遗传, 2013, 35: 875–884. Zheng Z Z, Shen J Q, Pan W H, Pan J W. Calcium sensors and their stress signaling pathways in plants., 2013, 35: 875–884 (in Chinese with English abstract).
[79] Harper J F, Harmon A. Plants, symbiosis and parasites: a calcium signaling connection., 2005, 6: 555–566.
[80] Kolukisaoglu U, Weinl S, Blazevic D, Batistic O, Kudla J. Calcium sensors and their interacting protein kinases: genomics of theand rice CBL-CIPK signaling networks., 2004, 134: 43–58.
[81] Upadhyaya H, Dutta B K, Sahoo L, Panda S K. Comparative effect of Ca, K, Mn and B on post-drought stress recovery in tea [(L.) O. Kuntze]., 2012, 3: 443–460.
[82] Yuan F, Yang H M, Xue Y, Kong D D, Ye R, Li C J, Zhang J Y, Theprungsirikul L, Shrift T, Krichilsky B, Johnson D M, Swift G B, He Y K, Siedow J N, Pei Z M. OSCA1 mediates osmotic-stress-evoked Ca2+increases vital for osmosensing in., 2014, 514: 367–371.
[83] Hamilton E S, Jensen G S, Maksaev G, Katims A, Sherp A M, Haswell E S. Mechanosensitive channel MSL8 regulates osmotic forces during pollen hydration and germination., 2015, 350: 438–441.
[84] He L R, Yang X Y, Wang L C, Zhu L F, Zhou T, Deng J W, Zhang X L. Molecular cloning and functional characterization of a novel cotton CBL-interacting protein kinase gene () reveals its involvement in multiple abiotic stress tolerance in transgenic plants., 2013, 435: 209–215.
[85] Jing P, Zou J Z, Kong L, Hu S Q, Wang B Y, Yang J, Xie G S. OsCCD1, a novel small calcium-binding protein with one EF-hand motif, positively regulates osmotic and salt tolerance in rice., 2016, 247: 104–114.
[86] 陈娇娆, 续旭, 胡章立, 杨爽. 植物感受盐胁迫及相关钙信号的研究进展. 植物研究, 2022, 42: 713–720. Chen J L, Xu X, Hu Z L, Yang S. Recent Advances on salt stress and related calcium signals in plants., 2022, 42: 713–720 (in Chinese with English abstract).
[87] Choi W G, Toyota M, Kim S H, Hilleary R, Gilroy S. Salt stress- induced Ca2+waves are associated with rapid, long-distance root-to-shoot signaling in plants., 2014, 111: 6497–6502.
[88] Feng W, Kita D, Peaucelle A, Cartwright H N, Doan V, Duan Q, Liu M C, Maman J, Steinhorst L, Schmitz-Thom I, Yvon R, Kudla J, Wu H M, Cheung A Y, Dinneny J R. The FERONIA receptor kinase maintains cell-wall integrity during salt stress through Ca2+signaling., 2018, 28: 666–675.
[89] 项洪涛, 郑殿峰, 何宁, 李琬, 王曼力, 王诗雅. 植物对低温胁迫的生理响应及外源脱落酸缓解胁迫效应的研究进展. 草业学报, 2021, 30(1): 208–219. Xiang H T, Zheng D F, He N, Li W, Wang M L, Wang S Y. Research progress on physiological response of plants to low temperature and the amelioration effcectiveness of exogenous ABA., 2021, 30(1): 208–219 (in Chinese with English abstract).
[90] Miura K, Furumoto T. Cold signaling and cold response in plants., 2013, 14: 5312–5337.
[91] Mori K, Renhu N, Naito M, Nakamura A, Shiba H, Yamamoto T, Suzaki T, Iida H, Miura K. Ca2+-permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca2+increase and cold tolerance in., 2018, 8: 550.
[92] Wang J C, Ren Y L, Liu X, Luo S, Zhang X, Liu X, Lin Q B, Zhu S S, Wan H, Yang Y, Zhang Y, Lei B, Zhou C L, Pan T, Wang Y F, Wu M M, Jing R N, Xu Y, Han M, Wu F Q, Lei C L, Guo X P, Cheng Z J, Zheng X M, Wang Y H, Zhao Z G, Jiang L, Zhang X, Wang Y F, Wang H Y, Wan J M. Transcriptional activation and phosphorylation of OsCNGC9 confer enhanced chilling tolerance in rice., 2021, 14: 315–329.
[93] Liu Q B, Ding Y L, Shi Y T, Ma L, Wang Y, Song C P, Wilkins K A, Davies J M, Knight H, Knight M R, Gong Z Z, Guo Y, Yang S H. The calcium transporter ANNEXIN1 mediates cold-induced calcium signaling and freezing tolerance in plants., 2021, 40: e104559.
[94] Chu M X, Li J J, Zhang J Y, Shen S F, Li C N, Gao Y J, Zhang S Q. AtCaM4 interacts with a Sec14-like protein, PATL1, to regulate freezing tolerance inin a CBF-independent manner., 2018, 69: 5241–5253.
[95] Zhang H F, Yang B, Liu W Z, Li H W, Wang L, Wang B Y, Deng M, Liang W W, Deyholos M K, Jiang Y Q. Identification and characterization of CBL and CIPK gene families in canola (L.)., 2014, 14: 8.
[96] Iqbal Z, Memon A G, Ahmad A, Iqbal M S. Calcium mediated cold acclimation in plants: underlying signaling and molecular mechanisms., 2022, 13: 855559.
[97] Iqbal Z, Shariq Iqbal M, Singh S P, Buaboocha T. Ca2+/Calmodulin complex triggers CAMTA transcriptional machinery under stress in plants: signaling cascade and molecular regulation., 2020, 11: 598327.
[98] Kidokoro S, Yoneda K, Takasaki H, Takahashi F, Shinozaki K, Yamaguchi-Shinozaki K. Different cold-signaling pathways function in the responses to rapid and gradual decreases in temperature., 2017, 29: 760–774.
[99] Wang Q, Yang S, Wan S B, Li X G. The significance of calcium in photosynthesis., 2019, 20: 1353.
[100] 陈向明, 郑国生, 张圣旺. 钙对保护地栽培牡丹光合特性的影响. 园艺学报, 2001, 28: 572–574.Chen X M, Zheng G S, Zhang S W. Effects of calcium on photosynthetic characteristics of peony cultivated in protected areas., 2001, 28: 572–574 (in Chinese with English abstract).
[101] 李敏, 吉文丽, 张恒, 李程程, 杨静萱, 张延龙. 外源Ca2+对油用牡丹凤丹白幼苗光合特性的影响. 西北林学院学报, 2017, 32(5): 39–45. Li M, Ji W L, Zhang H, Li C C, Yang J X, Zhang Y L. Effects of exogenous calcuim on photosynthetic characteristics and biomass of oil peony ostii ‘Fengdan White’., 2017, 32(5): 39–45 (in Chinese with English abstract).
[102] Feng N J, Yu M L, Li Y, Jin D, Zheng D F. Prohexadione-calcium alleviates saline-alkali stress in soybean seedlings by improving the photosynthesis and up-regulating antioxidant defense., 2021, 220: 112369.
[103] 秦喜彤. 钙调控对春玉米产量和氮肥利用效率的影响. 吉林大学硕士学位论文, 吉林长春, 2020. Qin X T. Effects of Calcium Regulation on Yield and Nitrogen Utilization Efficiency of Spring Maize. MS Thesis of Jilin University, Changchun, Jilin, China, 2020 (in Chinese with English abstract).
[104] Weinl S, Held K, Schlücking K, Steinhorst L, Kuhlgert S, Hippler M, Kudla J. A plastid protein crucial for Ca2+-regulated stomatal responses., 2008, 179: 675–686.
[105] Kreimer G, Melkonian M, Holtum J A, Latzko E. Stromal free calcium concentration and light-mediated activation of chloroplast fructose-1,6-bisphosphatase., 1988, 86: 423–428.
[106] Nishiyama Y, Yamamoto H, Allakhverdiev S I, Inaba M, Yokota A, Murata N. Oxidative stress inhibits the repair of photodamage to the photosynthetic machinery., 2001, 20: 5587–5594.
[107] Takamatsu H, Takagi S. Actin-dependent chloroplast anchoring is regulated by Ca2+-calmodulin in spinach mesophyll cells., 2011, 52: 1973–1982.
[108] Galeriani T M, Neves G O, Santos Ferreira J H, Oliveira R N, Oliveira S L, Calonego J C, Crusciol C A C. Calcium and boron fertilization improves soybean photosynthetic efficiency and grain yield.(Basel), 2022, 11: 2937.
[109] Zhang J Z, Li B S, Zhang J L, Christie P, Li X L. Organic fertilizer application and Mg fertilizer promote banana yield and quality in an Udic Ferralsol, 2020, 15: e0230593.
[110] Perveen S, Parveen A, Saeed M, Arshad R, Zafar S. Interactive effect of glycine, alanine, and calcium nitrate Ca(NO3)2on wheat (L.) under lead (Pb) stress., 2022, 29: 37954–37968.
[111] Kanu A S, Ashraf U, Mo Z, Sabir S U, Baggie I, Charley C S, Tang X. Calcium amendment improved the performance of fragrant rice and reduced metal uptake under cadmium toxicity., 2019, 26: 24748–24757.
[112] 张元棋. 外源钙对酸雨胁迫下水稻氮利用和稻米品质的影响. 江南大学硕士学术论文, 江苏无锡, 2022. Zhang Y Q. Effect of Exogenous Ca2+on Plasma Membrane Composition, Ca2+Distribution and Transport in Rice Roots under Acid Rain Stress. MS Thesis of Jiangnan University, Wuxi, Jiangsu, China, 2022 (in Chinese with English abstract).
[113] Metwally A M, Radi A A, El-Shazoly R M, Hamada A M. The role of calcium, silicon and salicylic acid treatment in protection of canola plants against boron toxicity stress., 2018, 131: 1015–1028.
[114] Hou J F, Li J, Yang Y, Wang Z X, Chang B W, Yu X W, Yuan L Y, Wang C G, Chen G H, Tang X Y, Zhu S D. Physiological and transcriptomic analyses elucidate that exogenous calcium can relieve injuries to potato plants (L.) under weak light., 2019, 20: 5133.
[115] 张浩, 郑云普, 叶嘉, 高伟, 乔雅君, 戴川景, 赵雨欣, 石少婕. 外源钙离子对盐胁迫玉米气孔特征、光合作用和生物量的影响. 应用生态学报, 2019, 30: 923–930. Zhang H, Zheng Y P, Ye J, Gao W, Qiao Y J, Dai C J, Zhao Y X, Shi S J. Effects of exogenous Ca2+on stomatal traits, photosynthesis, and biomass of maize seedings under salt stress., 2019, 30: 923–930 (in Chinese with English abstract).
[116] Li Y W, Liang C J. Exogenous application of Ca2+mitigates simulated acid rain stress on soybean productivity and quality by maintaining nutrient absorption., 2019, 26: 4975–4986.
[117] Lin K H, Lin T Y, Wu C W, Chang Y S. Protective effects of salicylic acid and calcium chloride on sage plants (L. andVahl) under high-temperature stress.(Basel), 2021, 10: 2110.
[118] Sugimoto T, Watanabe K, Yoshida S, Aino M, Furiki M, Shiono M, Matoh T, Biggs A R. Field application of calcium to reduce phytophthora stem rot of soybean, and calcium distribution in plants., 2010, 94: 812–819.
[119] 陈卫平, 杨阳, 谢天, 王美娥, 彭驰, 王若丹. 中国农田土壤重金属污染防治挑战与对策. 土壤学报, 2018, 55: 261–272. Chen W P, Yang Y, Xie T, Wang M E, Peng C, Wang R D. Challenges and countermeasures for heavy metal pollution control in farmalands of China., 2018, 55: 261–272 (in Chinese with English abstract).
[120] Ahmad P, Abdel Latef A A, Abd Allah E F, Hashem A, Sarwat M, Anjum N A, Gucel S. Calcium and potassium supplementation enhanced growth, osmolyte secondary metabolite production, and enzymatic antioxidant machinery in cadmium-exposed chickpea (L.)., 2016, 7: 513.
[121] Gong X M, Liu Y G, Huang D L, Zeng G M, Liu S B, Tang H, Zhou L, Hu X, Zhou Y Y, Tan X F. Effects of exogenous calcium and spermidine on cadmium stress moderation and metal accumulation in(L.) Gaudich., 2016, 23: 8699–8708.
[122] Zhang S, Li Q, Nazir M M, Ali S, Ou-Yang Y, Ye S, Zeng F. Calcium plays a double-edged role in modulating cadmium uptake and translocation in rice., 2020, 21: 8058.
[123] Nazir M M. 钙和氧化钙纳米颗粒减缓大麦砷和镉毒害及其机理. 浙江大学博士学位论文, 浙江杭州, 2022.Nazir M M. Alleviation of Arsenic and Cadmium Toxicity in Barley by Calcium and Calcium Oxide Nanoparticles (CaO NPs) and their Mechanisms. PhD Dissertation of Zhejiang University, Hangzhou, Zhejiang, China, 2022 (in Chinese with English abstract).
[124] Ji R J, Zhou L M, Liu J L, Wang Y, Yang L, Zheng Q S, Zhang C, Zhang B, Ge H M, Yang Y H, Zhao F G, Luan S, Lan W Z. Calcium-dependent protein kinase CPK31 interacts with arsenic transporter AtNIP1;1 and regulates arsenite uptake in., 2017, 12: e0173681.
[125] 尤召阳, 王建国, 刘颖, 闫振辉, 张佳蕾, 万书波. 氮钙互作对花生氮素利用及钙素积累的影响. 中国油料作物学报, 2023 [2023-10-16]. https://kns.cnki.net/kcms/detail/42.1429.s.20230614. 1821.001.html.You Z Y, Wang J G, Liu Y, Yan Z H, Zhang J L, Wan S B. Interactive effect of N and Ca on the nitrogen metabolism enzyme activity, nitrogen utilization and calcium accumulation of peanut., 2023 [2023-10-16] (in Chinese with English abstract). https://kns.cnki.net/kcms/detail/42.1429.s. 20230614.1821.001.html.
[126] 杜英俊, 杨洁, 张琪, 陈泽浩, 毛柯, 马锋旺, 李超. 喷施不同钙肥以及钙肥混施激素对‘蜜脆’苹果果实的影响. 陕西农业科学, 2022, 68(6): 54–63. Du Y J, Yang J, Zhang Q, Chen Z H, Mao K, Ma F W, Li C. Effects of spraying different calcium fertilizers and mixed application of calcium fertilizers with hormones on ‘Honey Crisp’ apple fruit., 2022, 68(6): 54–63 (in Chinese with English abstract).
[127] 张贤聪. 喷施钙肥对蓝莓品质及果胶降解的影响. 四川农业大学硕士学位论文, 四川雅安, 2019. Zhang X C. Effects of Calcium Fertilizer Spraying on Blueberry Quality and Pectin Degradation. MS Thesis of Sichuan Agricultural University, Ya’an, Sichuan, China, 2019 (in Chinese with English abstract).
[128] Amarante C V T, Katsurayama J M, Pereira A J, Steffens C A. Apple orchard spraying with commercial sources of calcium to improve fruit quality., 2020, 1275: 201–206.
[129] El-Hady E S, Merwad M A, Shahin M F M, Hagagg L F. Influence of foliar spray with some calcium sources on flowering, fruit set, yield and fruit quality of olive Kalmata and Manzanillo cultivars under salt stress., 2020, 44: 197.
[130] Wahab M, Ullah Z, Sajid M, Usman M, Sohail K, Nayab S, Ullah M. Effect of calcium sources as foliar applications on fruit quality of peach cultivars., 2016, 48: 167–172.
[131] El-Ghany M F A, El-Kherbawy M I, Abdel-Aal Y A, El-Dek S I, Abd El-Baky T. Comparative study between traditional and nano calcium phosphate fertilizers on growth and production of snap bean (L.) plants.(Basel), 2021, 11: 2913.
[132] Bakshi A, Choi W G, Kim S H, Gilroy S. The vacuolar Ca2+transporter CATION EXCHANGER 2 regulates cytosolic calcium homeostasis, hypoxic signaling, and response to flooding in., 2023, 240: 1830–1847.
[133] Islam M M, Munemasa S, Hossain M A, Nakamura Y, Mori I C, Murata Y. Roles of AtTPC1, vacuolar two pore channel 1, instomatal closure., 2010, 51: 302–311.
Function and application of calcium in plant growth and development
WANG Yu, GAO Geng-Dong, GE Meng-Meng, CHANG Ying, TAN Jing, GE Xian-Hong, WANG Jing, WANG Bo, ZHOU Guang-Sheng*, and FU Ting-Dong
College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, Hubei, China
Calcium is one of the essential elements in crops. It is widely present in roots, stems, leaves, flowers, fruits, and seeds, and is of great significance for the growth and development of crops. Calcium is a difficult element to redistribute, and its absorption and transport are subject to transpiration. Therefore, crops often experience physiological calcium deficiency, which weakens their stress resistance and reduces both yield and quality. Calcium in crops has dual functions. It not only participates in the formation of cell walls and membranes but also plays a role in responding to various environmental stimuli and internal growth and development signals as an intracellular second messenger. The absorption and transportation of calcium in cells are essential for maintaining intracellular calcium homeostasis and ensuring calcium signal transduction. In recent years, the function and application of calcium in crops have been extensively studied. In this study, we describe the distribution, absorption, transportation, and demand of calcium in crops, introduce the symptoms and causes of calcium deficiency in crops, review the nutritional structure functions of calcium, the second messenger function and the mechanism of calcium signal generation, transmission, and decoding, and summarize the role of calcium in crop growth and development, including its effects on yield, quality, and stress resistance. Meanwhile, the future research direction is proposed.
crop; calcium deficiency; calcium absorption; calcium transporter proteins; the second messenger; stress resistance
10.3724/SP.J.1006.2024.34145
本研究由国家重点研发计划项目“绿肥及增效产品创制与产业化” (2021YFD1700201)资助。
This study was supported by the National Key Research and Development Program “Creation and Industrialization of Green Fertilizer and Efficiency Enhancing Products” (2021YFD1700201).
周广生, E-mail: zhougs@mail.hzau.edu.cn
E-mail: wangyu4630@webmail.hzau.edu.cn
2023-07-31;
2023-10-29;
2023-11-17.
URL: https://link.cnki.net/urlid/11.1809.S.20231116.1641.002
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜欢
今日农业(2020年20期)2020-12-15 15:53:19
今日农业(2020年17期)2020-12-15 12:34:28
世界农药(2019年4期)2019-12-30 06:25:10
中国生殖健康(2019年12期)2019-01-07 01:54:36
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
广东农业科学(2017年5期)2017-08-29 10:37:54
上海农业学报(2016年2期)2016-10-27 00:49:58
故事作文·高年级(2015年12期)2016-01-22 21:18:11
爆笑show(2015年4期)2015-06-24 22:07:31
西南军医(2015年6期)2015-01-23 01:25:59
