
四川华鳊外周血液及血细胞发生的观察
2024-03-25 07:43:18石金凤刘小红汪浩宇刘小琴王志坚
水产学报 2024年3期
石金凤,刘小红,汪浩宇,刘小琴,王志坚
(西南大学生命科学学院,淡水鱼类资源与生殖发育教育部重点实验室,水产科学重庆市重点实验室,重庆 400715)
血液是动物体内十分重要的组织,除了物质循环和运输外,血细胞还承担了免疫应答、损伤修复等多种功能[1]。鱼类的血液对外界环境十分敏感,通过对鱼体血液的检测分析,可以了解到鱼类的健康状态及其生活环境状况[2-3]。鱼类是脊椎动物数量最大的类群,而目前对于鱼类血细胞的研究主要集中在鲤形目、鲈形目和鲇形目的部分物种,如斑马鱼(Danio rerio)、鲤(Cyprinus carpio)、淇河鲫(Carassius auratus)、军曹鱼(Rachycentron canadum)、点带石斑鱼(Epinephelus malabaricus)以及长吻鮠(Leiocassis longirostris)、南方鲇(Silurus meridionalis)等。然而由于不同鱼类生活环境的差异,不同鱼类血细胞在形态、功能、发生上可能存在较大差异。除了斑马鱼[4]、虹鳟(Oncorhynchus mykiss)[5]的血细胞和免疫系统方面有较深入的研究外,多数鱼类的有关研究还只能依赖于细胞学特征进行分类,然后利用这些基础研究开展深入研究。因此,为了开展鱼类资源的保护和开发利用,对于有潜在经济价值和科研价值的鱼类,有必要开展基础的血液学相关研究,为其将来的规模化、标准化、科学化人工养殖及健康评估提供参考。
四川华鳊(Sinibrama taeniatus)隶属于鲤形目(Cypriniformes)鲤科(Cyprinidae)鲌亚科(Cultrinae)华鳊属(Sinibrama),是长江上游特有的一种小型经济鱼类,具有重要的经济与生态价值。但由于其分布范围狭窄以及人为捕捞和水利设施修建等原因导致资源量持续下降[6]。有关四川华鳊的研究主要集中在年龄与生长[7]、胚胎发育[8-9]以及性腺发育[10]等方面。并且在本实验室研究中发现,达到性成熟后的四川华鳊在适宜的养殖条件下可14 d 繁殖一次,打破了其在野外环境下一年仅繁殖2 次的繁殖周期,具有重要的研究价值。鱼类的存活与其免疫系统息息相关,但目前对于四川华鳊血细胞以及免疫相关的研究还未见报道。本研究利用外周血涂片法和组织印片法,观察四川华鳊头肾、体肾、脾脏和肝脏4 种造血器官中血细胞的类型以及各类型间的数量比例,旨在了解四川华鳊血细胞的类型组成、各造血器官的造血功能以及血细胞发生的特征以及免疫系统的基本构成,促进四川华鳊的科学化人工养殖技术的建立,为推动长江特有鱼类资源的保护提供基础的生理资料。
1 材料与方法
1.1 实验材料
实验用四川华鳊来自西南大学淡水鱼类资源与生殖发育教育部重点实验室大棚循环水养殖缸。挑选体表无损伤,体色正常,健康活泼的12 尾四川华鳊[体长(8.6±0.8) cm,体重(9.5±0.4) g]转移至室内养殖缸中驯养7 d[11],驯养水为曝气24 h 的自来水,水温(25±0.5) °C,并利用充气泵增氧,每日定时2 次投喂饵料。
1.2 实验方法
实验用四川华鳊经MS-222(格林恒兴,北京)麻醉后,使用肝素钠处理过的1 mL 无菌注射器进行尾静脉采血[12],并制作外周血涂片[13]。采血后迅速解剖鱼体,分别取出肝脏、体肾、头肾和脾脏,0.65%生理盐水冲洗后,制作组织印片[13]。每个器官的每个切面印5 个印迹。待涂片与印片自然风干后,甲醇固定,Wright's-Giemsa 染色液(索莱宝,北京)染色,中性树脂封片,40 ℃烘箱干燥。
干燥后的装片于Eclipse 80i(Nikon,日本)显微成像系统下观察、计数、拍照。每尾鱼选取3张制作良好,染色均匀的外周血涂片,每张血涂片随机选取100 个白细胞进行分类计数。每尾鱼的头肾、体肾、脾脏、肝脏的组织印片于100 倍镜下各拍30 个视野,参照《野生动物血液细胞学图谱》[14]将各种细胞划分为原始型、幼稚型、成熟型3 个发育阶段,分类计数并计算出各类细胞在总细胞中的占比。Image J 测量各类血细胞及不同发育阶段各类型血细胞的胞体长短径以及胞核长短径,每种细胞测量100 个,使用SPSS 20.0 进行数据统计分析。
2 结果
2.1 四川华鳊外周血细胞组成
成熟红细胞是四川华鳊外周血主要细胞类型,淋巴细胞、血栓细胞、单核细胞以及嗜中性粒细胞等较少(图版Ⅰ),可偶见浆细胞(图版Ⅰ-15),未观察到嗜酸性粒细胞和嗜碱性粒细胞。四川华鳊外周血中还发现有少量处于晚幼阶段的红细胞(图版Ⅰ-9);并可观察到红细胞分裂现象(图版Ⅰ-2~4)。未观察到其他类型的未成熟血细胞。

图版Ⅰ 外周血涂片中各类型血细胞1.成熟的红细胞;2~3.正在分裂的红细胞(胞质分裂,胞核不分裂);4.正在分裂的红细胞(胞核胞质均分裂);5.衰老的红细胞;6.红细胞胞质小体;7.晚幼红细胞;8.大淋巴细胞;9.小淋巴细胞;10.血栓细胞(蝌蚪形和长椭圆形);11.单核细胞;12.分叶的单核细胞;13.嗜中性粒细胞;14.分叶的嗜中性粒细胞;15.浆细胞。Plate Ⅰ All kinds of blood cells in the peripheral blood smears1.erythrocyte;2-3.dividing of erythrocyte (the cytoplasm divides but the nucleus does not divide);4.dividing of erythrocyte (the cytoplasm and nucleus divide equally);5.old erythrocyte;6.erythroid plastid;7.orthochromatic erythroblast;8.large lymphocyte;9.small lymphocyte;10.thrombocyte (tadpole shaped and long oval shaped);11.monocyte;12.monocyte segmented cell;13.neutrophil;14.neutrophil segmented cell;15.plasmocyte.
测量各类血细胞的大小,并统计各类白细胞占比(表1)。从表1 可以看出,从细胞长径看,红细胞最大;从细胞大小、短径和核径看,都是单核细胞最大,小淋巴细胞最小。红细胞长短径比值在1.69~1.88。白细胞中淋巴细胞占比最高,为65.79%±0.10%;其次是血栓细胞,占16.76%±0.07%;再次为嗜中性粒细胞,占13.92%±0.09%;单核细胞含量少,仅有3.33%±0.01%。
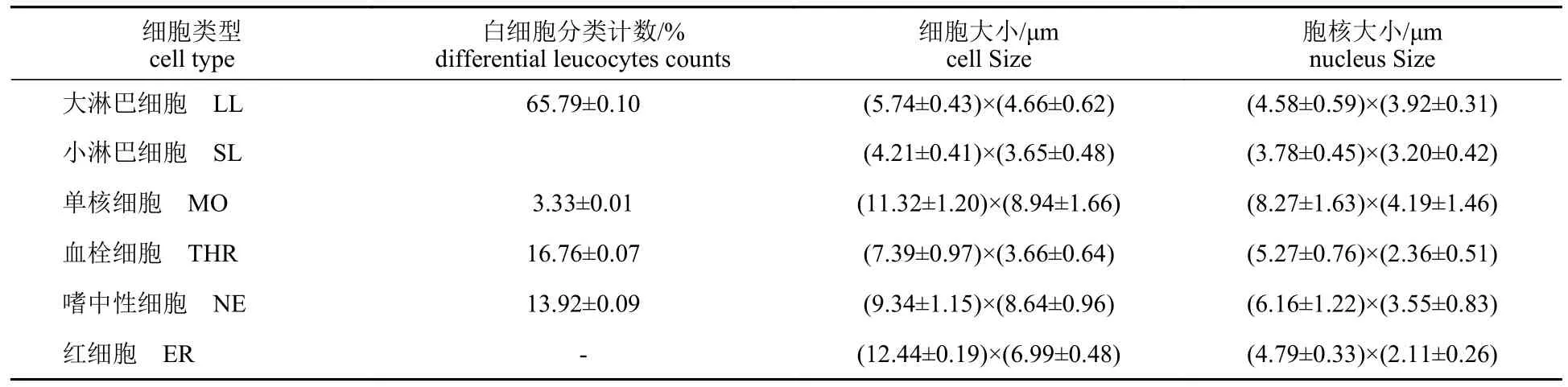
表1 四川华鳊外周血细胞数量百分比及细胞大小Tab.1 Differential leucocytes counts (DLC) and the sizes of different blood cells
2.2 各类血细胞不同发育阶段的形态特征
通过4 种器官的印片观察,可将血细胞分为红细胞、单核细胞、淋巴细胞、粒细胞4 个细胞系;每种血细胞又可分为原始型、幼稚型、成熟型3 个发育阶段;但具体阶段又有所差异,各细胞在形态和细胞大小上有较大差异(图版Ⅱ,表2)。

表2 四川华鳊各类血细胞大小Tab.2 The sizes of different blood cells of S.taeniatus
红细胞系 红细胞的发育可分为原红细胞(proerythroblast,PEB)、早幼红细胞(basophilic erythroblast,BE)、晚幼红细胞(orthochromatic erythroblast,OE)和成熟红细胞(erythrocyte,ER) 4个阶段。
原红细胞近圆形,边缘不规则,偶尔可见边缘有伪足状突起。胞体长径为(11.05±0.52) μm,短径为(9.48±0.72) μm。胞浆量少,染为深蓝色。胞核大,占胞体大部,多呈较规则的圆形,胞核长径(8.11±0.89) μm,胞核短径(6.15±0.68) μm。胞内核质呈颗粒状,排列疏松,染为紫红色(图版Ⅱ-1)。
早幼红细胞近圆形,部分有伪足状的突起,胞体长径(10.46±0.74) μm,胞体短径(9.07±0.97) μm,胞浆染为蓝色。胞核圆形,多偏位,胞核长径(7.11±0.99) μm,胞核短径(5.30±0.80) μm。相对原红细胞而言胞核凝缩变小,染色质粗糙密集成块,染为浅紫红色(图版Ⅱ-2)。
晚幼红细胞多为规则圆形,胞体缩小,是红细胞发育过程中胞体最小的发育阶段。胞体长径(8.28±1.02) μm,胞体短径(7.46±0.66) μm。胞浆中已有大量血红蛋白的合成,染成橘红偏灰色。胞核圆形居中,胞核凝缩,胞核长径(5.41±0.73) μm,胞核短径(4.54±0.81) μm,染为深紫色(图版Ⅱ-3)。
成熟红细胞呈长椭圆形或近卵圆形,胞体较大,其长径为(11.76±0.83) μm,短径为(6.40±0.57)μm。胞浆丰富,内部充满血红蛋白,染色为橘红色或浅红色,颜色均匀。胞核呈椭圆形,胞核长径为(4.82±0.53) μm,短径为(2.27±0.32) μm,核居中,染色质交织成网致密排布,染为蓝紫色(图版Ⅱ-4)。
淋巴细胞系 淋巴细胞的发育包括了原淋巴细胞(lymphoblast,LB)、幼淋巴细胞(prolymphocyte,PL)和成熟淋巴细胞(mature lymphocyte,ML)3 个阶段。
原淋巴细胞呈规则圆形或卵圆形,边缘平整。胞体长径(10.03±0.56) μm,胞体短径(8.97±0.67) μm,是血细胞系中原始细胞最小的一类。胞浆少,染为蓝色或亮蓝色,在胞核与胞浆交界处常有一圈狭窄的环核带。胞核圆形居中或稍偏位,核质比大,胞核长径(7.44±0.84) μm,胞核短径(6.20±0.88) μm,染为紫红色,核染质交织呈网状排布(图版Ⅱ-5)。
幼淋巴细胞类圆形,胞体长径(8.54±0.70) μm,胞体短径(7.21±0.67) μm。胞浆较少,染为淡蓝色。部分细胞胞浆内含有紫红色嗜天青颗粒。胞核圆形或不规则类圆形,居中或偏位与质膜相切,胞核长径(5.97±0.62) μm,胞核短径(4.78±0.70) μm,核染色质相对聚集浓缩,染为浅紫红色(图版Ⅱ-6)。
成熟淋巴细胞根据大小可分为大淋巴细胞(large lymphocyte,LL)和小淋巴细胞(small lymphocyte,SL)。大淋巴细胞多呈圆形,偶尔可见伪足。胞体长径(5.53±0.66) μm,胞体短径(4.48±0.67)μm,胞浆较少,染成蓝色。胞核呈圆形或椭圆形,长径(4.05±0.51) μm,短径(3.20±0.66) μm,核染色质浓缩致密,呈较深的紫红色(图版Ⅱ-7)。小淋巴细胞长径为(4.21±0.50) μm,短径为(3.43±0.50) μm。胞体呈类圆形,常有伪足伸出。胞浆量极少,胞浆为淡蓝色。胞核圆形居中,胞核长径(3.21±0.50) μm,胞核短径(2.60±0.47) μm,染为深紫红色,相对大淋巴细胞而言,核染质更加凝缩,染色更深(图版Ⅱ-8)。
粒细胞系 粒细胞的发育分为原粒细胞(myeloblast,MYB)、早幼粒细胞(promyelocyte,PML)、中幼粒细胞(myelocyte,MYC)、晚幼粒细胞(metamyelocyte,MML)和成熟粒细胞(mature granulocyte,MG)。嗜中性粒细胞经历原粒细胞、早幼粒细胞、中性中幼粒细胞(neutrophilic myelocyte,NM)、中性晚幼粒细胞(neutrophilic metamyelocyte,NMM)和成熟的嗜中性粒细胞(neutrophil,NE)5 个阶段。
原粒细胞为圆形或卵圆形,边缘整齐,较原红细胞大,胞体长径(11.60±0.61) μm,胞体短径(10.19±0.87) μm。胞浆较少,染色为着色均匀的深蓝色。胞核圆形或椭圆形,多居中也有偏位的现象,核质比较大,胞核长径(8.63±0.77) μm,胞核短径(6.97±1.08) μm。核染色质呈微细颗粒状排布,染为浅蓝紫色(图版Ⅱ-9)。
早幼粒细胞呈圆形或椭圆形,胞体长径(10.68±0.98) μm,胞体短径(9.56±0.92) μm,胞浆量较多,呈天蓝色,存在紫红色的嗜天青颗粒。胞核多为椭圆形,位于中央或稍偏位,胞核长径(7.30±0.80) μm,胞核短径(4.87±1.12) μm,胞核逐渐缩小,核染质有凝集趋势,胞核染为较深的粉紫色(图版Ⅱ-10)。
中性中幼粒细胞为圆形或类圆形,胞体长径(10.42±0.79) μm,胞体短径(9.70±0.71) μm。胞浆量多,呈蓝粉色且充满紫红色颗粒。核染色质进一步固缩,胞核缩小,胞核长径(7.23±0.63) μm,胞核短径(4.82±0.51) μm。胞核圆形或类圆形,居中或偏位,染为紫红色(图版Ⅱ-11)。
中性晚幼粒细胞为圆形或类圆形,胞体长径(11.19±0.99) μm,胞体短径(10.02±1.05) μm。胞浆量多,胞浆与中幼嗜中性粒细胞相似,染色为淡红色,含有丰富的紫红色颗粒。胞核呈肾形或卵圆形,胞核长径(7.27±1.21),胞核短径(4.45±1.25)。核染色质较粗糙交织成粗粒网状,染成深紫色(图版Ⅱ-12)。
成熟的嗜中性粒细胞圆形或长卵圆形,胞体长径(10.82±1.57) μm,胞体短径(9.61±1.25) μm,胞浆丰富,胞浆内有细小的紫红色中性特异性颗粒。胞核呈圆形、肾形,有时有分叶,偏位与质膜相切,胞核大小一般为胞体的三分之一,胞核长径(7.22±1.00) μm,胞核短径(4.49±0.22) μm,核染色质固缩粗糙形成颗粒网状(图版Ⅱ-13)。
单核细胞系 单核细胞的发育包括了原单核细胞(monoblast,MB)、幼单核细胞(promonocyte,PM)和成熟单核细胞(monocyte,MO) 3 个阶段。
原单核细胞呈圆形、椭圆形或不规则形,胞体相对其他的原始血细胞大,其胞体长径(14.41±1.20) μm,胞体短径(11.56±1.38) μm,染为较深的蓝色或灰蓝色。胞核形态多样,为长椭圆形、肾形或不规则形状,胞核长径(10.09±1.38) μm,胞核短径(7.54±1.24) μm。胞核染色质呈疏松纤细网状,染为淡紫红色(图版Ⅱ-14)。
幼单核细胞为长椭圆形或不规则形,胞体长径为(13.73±0.75) μm,胞体短径(11.26±0.94) μm,胞浆量多,偶见空泡,呈天蓝色或浅蓝色。胞核形状多为不规则形或长椭圆形,多偏位。胞核相较原单核细胞较小,胞核长径(8.80±1.15) μm,胞核短径(6.32±0.81) μm,核染色质逐渐凝缩成疏松网状,染为浅紫红色(图版Ⅱ-15)。
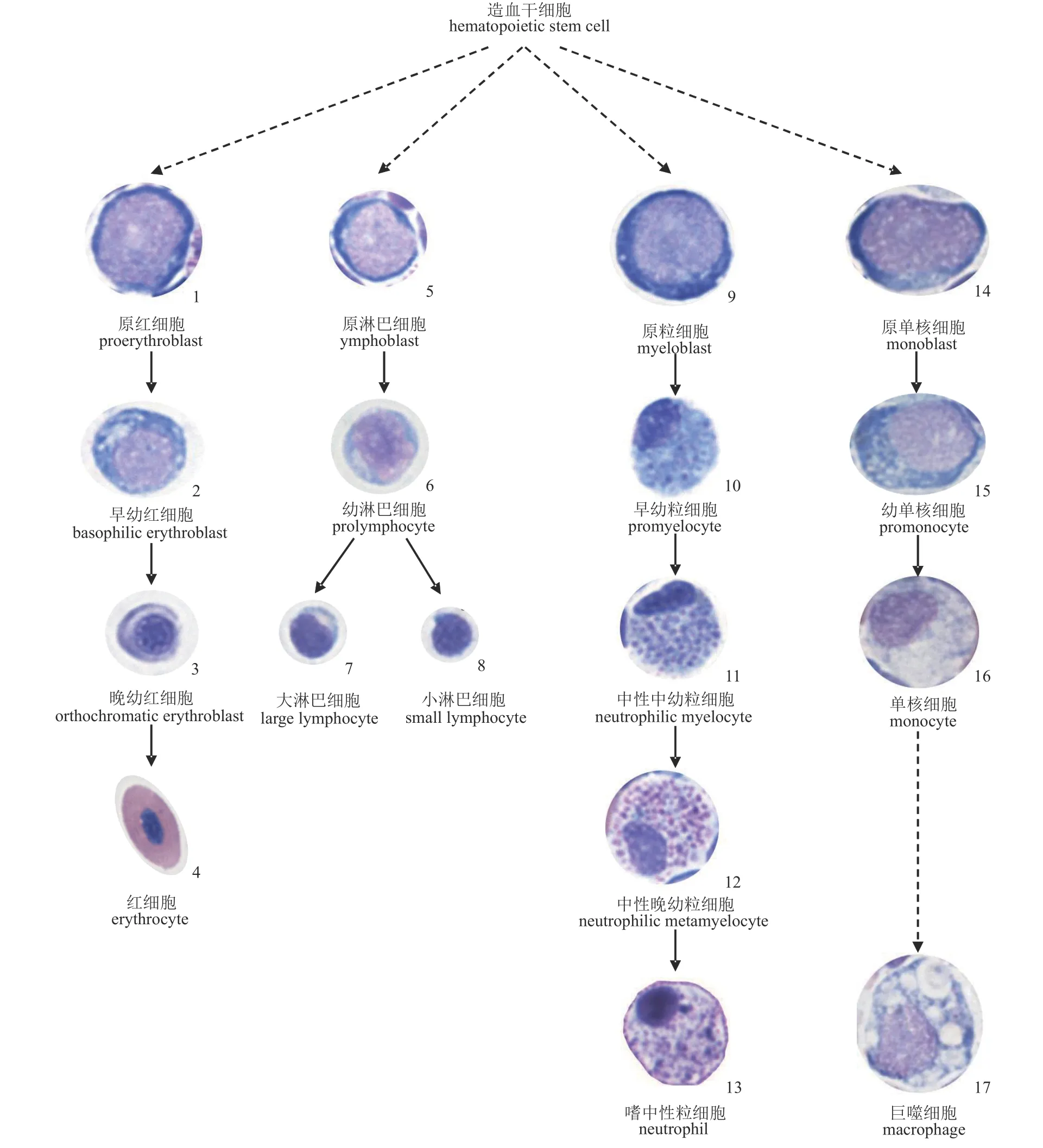
图版Ⅱ 四川华鳊血细胞发生过程Plate Ⅱ Hematocytes in different developmental stage in S.taeniatus
成熟单核细胞多为圆形或椭圆形,也有不规则形状,有时出现伪足。胞体是所有成熟血细胞中最大的一种。胞体长径为(12.43±0.87) μm,胞体短径为(10.42±1.07) μm,胞质淡蓝色或灰蓝色,少数有空泡。胞核有圆形、肾形、不规则形等,偶见分叶,多偏向一侧与质膜相切;胞核较大,可占到整个细胞的三分之一甚至二分之一,胞核长径为(8.36±1.27) μm,胞核短径为(5.86±0.92) μm。核染色质粗糙,分布浓密,呈粗糙疏网状,染成较深的红紫色(图版Ⅱ-16)。
其他细胞 血栓细胞(Thrombocyte,THR)一般呈长椭圆形或蝌蚪形,体积较小,胞体长径(7.39±0.97) μm,胞体短径(3.66±0.64) μm。胞浆少,染色均匀,呈淡蓝色,有时甚至无色。胞核也多为长棒状或长椭圆形,胞核长径(5.27±0.76) μm,胞核短径(2.36±0.51) μm,核染色质凝缩紧密,染成蓝紫色(图版Ⅰ-10)。
巨噬细胞(Macrophage,MP)由单核细胞发育而来的一类细胞[15]。胞体形状不规则,有伪足,胞体长径(17.16±1.81) μm,胞体短径(12.88±1.66)μm,胞浆丰富,空泡多且大。胞浆染为不均匀的淡蓝色且含有细小紫红色颗粒。胞核圆形或不规则形,多偏位。胞核长径(10.32±0.80) μm,胞核短径(9.13±0.84) μm,核染色质呈细网状排布,染为浅紫色(图版Ⅱ-17)。
浆细胞(Plasmocyte,PLA)呈椭圆形或卵圆形,边缘不整齐并有伪足突出。胞体长径(12.68±1.12),胞体短径(10.17±0.44),胞浆染为深蓝色。胞核椭圆形,胞核长径(8.87±0.39),胞核短径(5.02±0.42),核染色质粗糙,凝缩成深紫红色块状(图版Ⅰ-15)。
2.3 四川华鳊血细胞的发生场所
为清晰展示四川华鳊的血细胞发生场所的差异,统计了4 种器官印片中各类血细胞分类计数及百分比(表3),发现在头肾印片中,可见大量的处于不同发育阶段的血细胞,处于原始阶段以及幼稚阶段的红细胞、淋巴细胞、粒细胞和单核细胞都较为丰富(图版Ⅲ-1)。体肾印片也可以发现很多处于不同发育阶段的血细胞,但除原粒细胞外的各类原始细胞数量比头肾少(图版Ⅲ-2)。脾脏印片可观察到大量的成熟红细胞,在各类细胞中占比最高,而处于原始阶段和幼稚阶段的血细胞数量较少(图版Ⅲ-3)。肝脏印片中各类血细胞的数量少于其他3 个组织器官,主要为成熟的红细胞,除少数原淋巴细胞外,未发现其他原始血细胞(图版Ⅲ-4)。
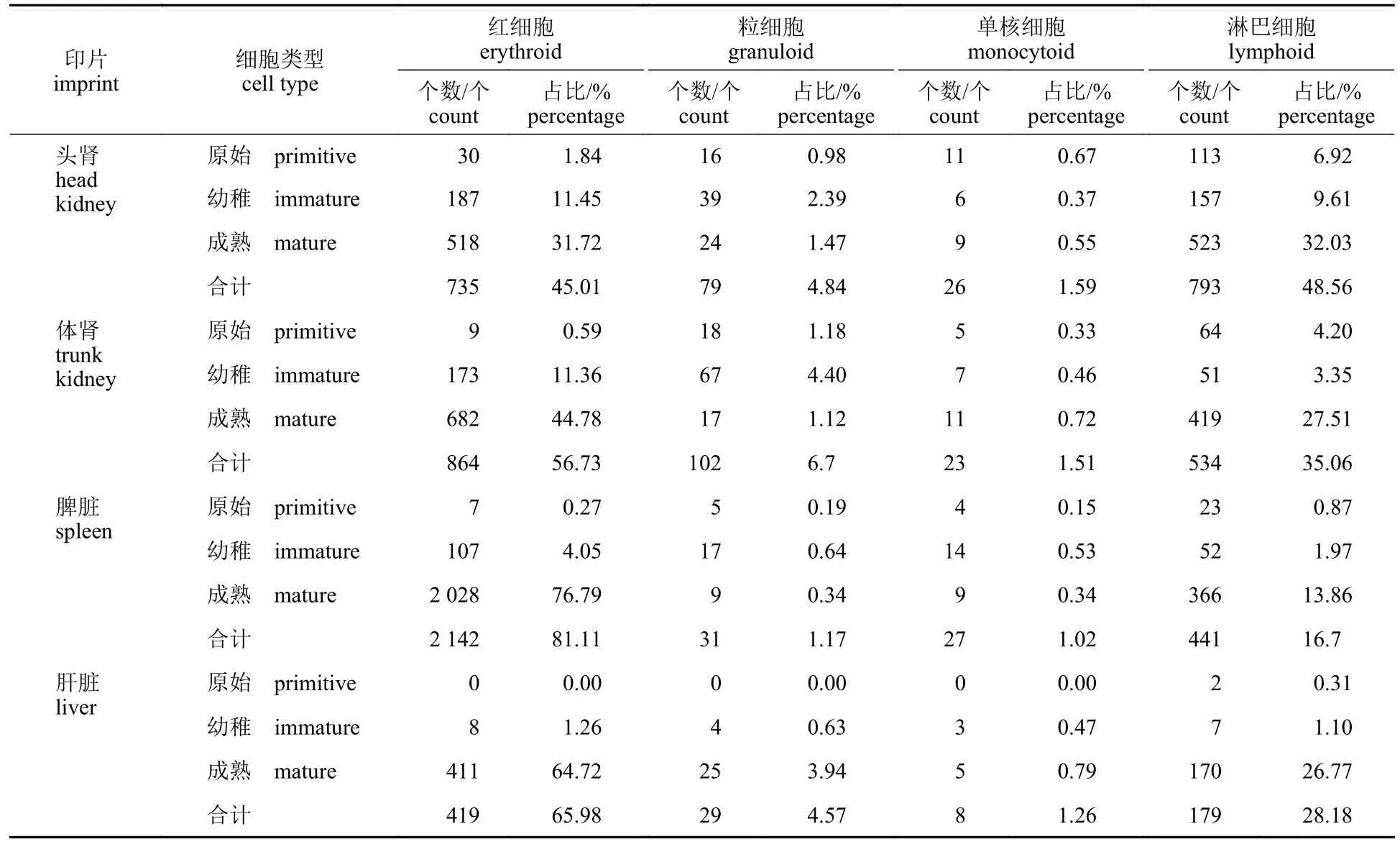
表3 四川华鳊四种器官印片中各类血细胞分类计数及百分比Tab.3 The count and percentage of different blood cells in four organs of S.taeniatus
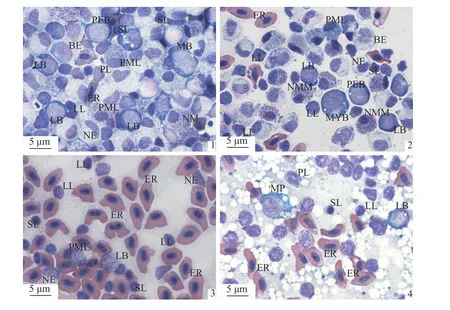
图版Ⅲ 四种器官的印片1.头肾印片;2.体肾印片;3.脾脏印片;4.肝脏印片。PEB.原红细胞;BE.早幼红细胞;ER.红细胞;LB.原淋巴细胞;PL.幼淋巴细胞;LL.大淋巴细胞;SL.小淋巴细胞;MB.原单核细胞;MYB.原粒细胞;PML.早幼粒细胞;NM.中性中幼粒细胞;NMM.中性晚幼粒细胞;NE.嗜中性粒细胞;MP.巨噬细胞。Plate Ⅲ Four kinds of organ imprints1.pronephros imprint;2.body kidney imprint;3.liver imprint;4.spleen imprint.PEB.proerythroblast;BE.basophilic erythroblast;ER.erythrocyte;LB.lymphoblast;PL.prolymphocyte;LL.large lymphocyte;SL.small lymphocyte;MB.monoblast;MYB.myeloblast;PML.promyelocyte;NM.neutrophilic myelocyte;NMM.neutrophilic metamyelocyte;NE.neutrophil;MP.macrophage.
3 讨论
3.1 四川华鳊外周血细胞特点
四川华鳊外周血细胞主要包括红细胞和白细胞,后者包括淋巴细胞、嗜中性粒细胞、单核细胞以及血栓细胞。在印片以及外周血中均未观察到嗜碱性粒细胞和嗜酸性粒细胞。大多数鱼类具有嗜酸性粒细胞,仅有少数鱼类如鲤[16]、军曹鱼[17]等观察到少量嗜碱性粒细胞。脊椎动物的血液中,嗜碱性粒细胞的嗜碱性颗粒易被酸中和,被水溶解,出现脱粒空白区,因此在制片过程中细胞易变形或破裂[14],这一现象在军曹鱼[17]、长吻鮠[18]中均有发现。在四川华鳊外周血与4 种器官的印片中均未观察到嗜碱性粒细胞,可能是由于其数量少,制片过程极易对其造成破坏导致。普遍认为,外周血是成熟红细胞的分布场所。但在四川华鳊的外周血中还含有正经历分裂的红细胞以及少量处于幼稚阶段的红细胞,这种现象在军曹鱼[17]、大弹涂鱼(Boleophthalmus pectinirostris)[19]、齐口裂腹鱼(Schizothorax prenanti)[20]等中有发现。在四川华鳊的外周血中,观察到两种红细胞的分裂方式,一种是核质均等分裂产生2 个细胞,这与大弹涂鱼[19]、淇河鲫[21]红细胞直接分裂相似;另一种是胞质分裂,胞核不分裂,产生有核的红细胞和无核的胞质小体,此种分裂方式与长吻鮠[18]红细胞的直接分裂相同。但鱼类红细胞的两种不同分裂方式在同一种鱼中发生还未见报道。鱼类的红细胞被认为是分化程度低,特化不完全的细胞[22],其本身具有较好的再生能力;也有认为出现直接分裂的红细胞是细胞衰老的一种表征,胞质分裂但胞核不分裂会产生衰老红细胞和胞质小体,最终被吞噬细胞吞噬[23]。但核质均等分裂所产生的红细胞是否为功能性细胞还需要进一步的研究。因此,成熟红细胞的直接分裂可能是四川华鳊的红细胞增殖方式之一,但这种直接分裂的现象被观察到的频率极少,不是红细胞主要的增殖方式。
四川华鳊的外周血中数量最多的是红细胞。红细胞的长短径可以反映红细胞的基本形态:比值越大,表明红细胞越呈长椭圆形;比值越接近1,表明红细胞越呈圆形。四川华鳊红细胞长短径比值在1.69~1.88,属于典型的长椭圆形红细胞。四川华鳊各类白细胞比例差异为淋巴细胞>血栓细胞>嗜中性粒细胞>单核细胞,以淋巴细胞为主要的白细胞类群,这一特性与鳙(Aristichthys nobilis)[24]、花斑裸鲤(Gymnocypris eckloni)[25]相同;而麦穗鱼(Pseudorasbora parva)[26]、齐口裂腹鱼[20]、瓦氏黄颡鱼(Pelteobagrus vachelli)[27]等其白细胞类群以血栓细胞为主。淋巴细胞主要的功能是产生抗体参与免疫应答,血栓细胞除了凝血功能外还具有吞噬和其他可能的免疫功能[28-29]。外周血中淋巴细胞和血栓细胞含量的差异与鱼体免疫能力的关系还需进一步研究。
3.2 鱼类血细胞发育过程
各类血细胞在发育的过程中都会经历细胞大小和细胞内容物的变化过程。四川华鳊从原红细胞发育到幼红细胞的过程为胞体体积逐渐变小,而在幼红细胞发育成熟过程中则又逐步变大,这与紫红笛鲷(Lutjanus argentimaculatus)[30]等的红细胞在发育中一直变小不同,但与军曹鱼[17]、美国红鱼(Sciaenops ocellatus)[31]、施氏魮(Barbonymus schwanenfeldii)[11]等的发育过程相一致。四川华鳊两类淋巴细胞的大小在发育过程中逐渐减小的现象与美国红鱼[31]、军曹鱼[17]等相同,而与淡水石斑鱼(Cichlasoma managuense)[32]的由小变大再变小不同。四川华鳊单核细胞胞体在发育过程中也是由大变小,这种变化以及细胞染色状态的变化与长吻鮠[23]、施氏魮[11]等相同。四川华鳊粒细胞的发育过程中,细胞大小则表现为先变小后变大再变小,这种现象与点带石斑鱼[33]等粒细胞发育过程相同,但与紫红笛鲷[30]、南方鲇[34]等表现出胞体一直变小不同。就目前为止,尚未发现不同种鱼的各类血细胞的发育均具有相同的变化特征的现象,这说明鱼类血细胞的发育具有物种特异性。
嗜天青颗粒被认为是初级溶酶体,含有多种水解酶,能消化分解异物,是免疫细胞发挥功能的关键细胞器之一[35]。与长吻鮠[18]相似,四川华鳊少数幼淋巴细胞和成熟淋巴细胞内含有紫红色的嗜天青颗粒,且成熟阶段更多。除了淋巴细胞,未成熟粒细胞也都有相似的现象。已有研究认为,小鼠(Mus musculus)体内白细胞随着发育阶段的推进,细胞中参与活性氧产生、吞噬和趋化的关键基因表达逐渐增强,细胞免疫功能逐步增强[36]。因此,鱼类未成熟白细胞也可能就已经具有免疫功能,并随着发育阶段的推进而增强。
3.3 鱼类血细胞发生的场所
除了血细胞的发育过程,鱼类各类血细胞的发生场所,也存在一定的种间差异性。南方鲇[34]的肝脏和脾脏是红细胞发生的主要场所,淋巴细胞、粒细胞、单核细胞主要发生在头肾和体肾;军曹鱼[17]的头肾和中肾是主要的血细胞发生场所,脾脏主要产生粒细胞,肝脏也能产生粒细胞和淋巴细胞。而在紫红笛鲷[30]体内,头肾、体肾和脾脏都是重要的血细胞发生场所,但肝脏无造血功能。各器官中各类原始血细胞含量的差异可反映其造血能力的强弱[37]。根据本实验中观察统计的结果(表3)可判断,头肾、体肾、脾脏都是四川华鳊的造血器官,但也具有一定分工。头肾是含有原红细胞、原淋巴细胞、原单核细胞数量最多的器官,是四川华鳊最重要的血细胞发生场所;体肾印片中,原粒细胞含量比头肾多,因此体肾是粒细胞发生的主要场所,但也在头肾大量发生,脾脏内少量产生。在脾脏印片中观察到的成熟红细胞数量最多,占各阶段红细胞总和的94.68%,但原始阶段数量极少,说明脾脏是四川华鳊极重要的储血器官。在肝脏印片中能观察到原淋巴细胞,说明肝脏也具有一定的造血功能,但局限于少数淋巴细胞的发生。肝脏中仅观察到一定幼稚阶段和较多成熟阶段的红细胞、粒细胞、单核细胞,提示幼稚阶段的血细胞形成后可迁移至其他造血器官并逐渐发育成熟。由此可见,头肾、肾脏和脾脏为鱼类普遍性的造血场所,但肝脏是否有造血功能以及其造血能力的强弱具有很大的物种差异性。另外,各类造血器官在造血的功能上既可以团结协作,又有较为明确的分工,这种分工具有物种差异性。
(作者声明本文无实际或潜在的利益冲突)
猜你喜欢
机械科学与技术(2023年1期)2023-02-16 12:25:40
大电机技术(2022年3期)2022-08-06 07:48:38
石家庄铁路职业技术学院学报(2021年1期)2021-06-09 06:07:00
现代塑料加工应用(2021年5期)2021-02-28 08:18:58
控制理论与应用(2020年6期)2020-07-15 02:24:26
中国现代神经疾病杂志(2020年3期)2020-04-01 07:28:04
图学学报(2020年1期)2020-03-19 04:39:24
中国现代神经疾病杂志(2018年2期)2018-05-09 07:47:02
中国现代神经疾病杂志(2018年1期)2018-05-07 02:49:44
物理化学学报(2015年7期)2015-12-30 12:13:16
