
生长分化因子5与代谢性疾病*
2024-03-23 07:10王天幕任无竞田振军
生物化学与生物物理进展 2024年3期
王天幕 任无竞 田振军
(陕西师范大学体育学院暨运动生物学研究所,西安 710119)
生长分化因子5(growth/differentiation factor-5,GDF-5)是转化生长因子β(transforming growth factor-β,TGF-β)家族成员之一,在骨、软骨和肌腱生长发育中发挥关键作用[1]。GDF-5在心脏、大脑、肾脏、骨骼肌和肌腱、肝脏以及脂肪等诸多器官组织中表达,具有促进细胞增殖分化、减少氧化应激、细胞凋亡和组织纤维化等生物学作用[2]。代谢性疾病如心血管疾病、肌肉减少症、代谢性骨病、代谢相关脂肪性肝病(metabolic associated fatty liver disease,MAFLD)、糖尿病和肥胖症等,其发病机制主要为活性氧(reactive oxygen species,ROS)激增引发线粒体功能障碍、DNA 损伤和蛋白质结构紊乱等[3-4],最终导致氧化应激和炎症。神经退行性疾病虽未明确归类为代谢性疾病,但因发病机制相似,故将其归类为神经代谢性疾病。细胞信号转导分子Smad1/5/8、胞内磷脂酰肌醇-3-激酶(phosphoinositide-3-kinase,PⅠ3K)/蛋白激酶B(protein kinase B,Akt)[5]和p38 丝裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38-MAPK)[2]是GDF-5/骨形态发生蛋白受体(bone morphogenetic proteins recptor,BMPR)的下游信号通路。本文通过梳理近年来GDF-5 及其与代谢性疾病相关文献,总结GDF-5 在各器官中发挥作用的具体机制,为认识GDF-5 在改善代谢性疾病中发挥的作用提供新的见解。
1 生长分化因子5(GDF-5)概述
1.1 GDF-5的分子结构
1994 年Hotten 等[6]发现,GDF-5 编码基因全长488 kb,定位于20q11.2 染色体位点,有2 个外显子,编码501个氨基酸,由位于N端的信号肽和C 端的成熟肽3 部分构成。GDF-5 前体蛋白经过转录翻译裂解为成熟肽,修饰加工后的GDF-5 蛋白含有120个氨基酸残基,并以二聚体形式发挥生物活性[7]。GDF-5包含7个半胱氨酸氨基酸残基,由7 个链间二硫键(S—S)维持成熟蛋白的四级结构,其结构与骨形态发生蛋白7(bone morphogenetic protein 7,BMP-7)十分相似[8]。T201P 位点处于GDF-5前区和成熟区连接处,该位点突变可能会影响GDF-5 片段的完整性。通过测定GDF-5 前区发现,T201P和L263P位点突变会导致GDF-5前体蛋白功能丧失,还可能导致患者手或足趾短小或偏斜[10]。最新研究发现,成熟的GDF-5 可以和成熟的BMP-2和BMP-4结合形成异源二聚体[11]。鉴于GDF-5在不同组织中发挥多样的生物学作用,因此也被称软骨形态发生蛋白1 (cartilage-derived morphogenetic protein-1,CDMP-1)和BMP-14。
1.2 GDF-5的作用机制
GDF-5及其他TGF-β家族成员一般通过两种类型受体进入胞内。Ⅰ型受体包括激活素样受体激酶(activin receptor-like kinase, ALK) 1~7, 其中ALK-1/-2/-3/-6也称作BMPR-Ⅰ,其高度保守的丝氨酸、甘氨酸结构域是活化的起始点[12];ⅠⅠ型受体包括BMPR-ⅠⅠ、激活素受体ⅠⅠ(activin a receptor type ⅠⅠ,ACTR-ⅠⅠ)[13]。GDF-5 和大多BMP 成员与BMPR-Ⅰ和BMPR-ⅠⅠ具有较高亲和力[14-15]。Sieber等[16]构建出单体GDF-5,并证明与二聚体形式的GDF-5 有相同的生物学功能,且对BMPR-Ⅰ和BMPR-ⅠⅠ都具有高度的亲和性。在已知的7 种Ⅰ型受体中,BMP 受体BMPR-ⅠA(ALK-3)和BMPR-ⅠB(ALK-6)与骨骼结构形成高度相关[17-20]。有学者认为部分BMP 成员与BMPR-Ⅰ亲和力较差,需要BMPR-ⅠⅠ激活BMPR-Ⅰ甘氨酸结构与并使其磷酸化,受体复合物形成的异四聚体与BMPs进入胞内激活Smad 信号[21-22]。Smad 信号通路如Smad1/5/8和Smad2/3被认为是TGF-β下游最常见的非依赖性通路[22]。GDF-5 与BMPR 结合后,使R-Smads 通路激活并形成Smad蛋白异源复合物[23];与Smad4结合后,进入细胞核与特定DNA 位点结合,介导靶基因转录,调节软骨内成骨速度,上调软骨肥大细胞增殖水平[1]。相反,Smad7 则是BMP 下游的负调节因子,通过竞争Ⅰ型受体以阻断Smad2/3 活化[7]。但心肌Smad7 特异性丢失会增加梗死心脏心室不良重塑,心脏代谢功能恶化,而过表达Smad7则会降低Smad2/3表达,抑制胶原沉积,减少心肌组织纤维化[24]。此外,Smad 泛素调节因子1(smad ubiquitination regulatory factor 1,Smurf1)也可降解BMPR和R-Smads[22,25](图1)。
1.3 GDF-5与生物学功能
GDF-5与骨和软骨发育生长息息相关,在骨关节炎中发挥巨大的修复和保护作用[26]。GDF-5 表达水平随早期胚胎肢体形成而发生相应变化,在肢体软骨前体细胞聚集区、长骨发育的软骨核心和关节形成区表达依次升高,调节肢体骨骼发育和骨稳态平衡[1,14]。炎症是代谢性疾病的发病源之一,也是发病后的主要表征之一。目前认为,代谢紊乱导致的炎症是骨关节炎发生的主要因素[27]。文献表明,GDF-5可促进软骨发育,且在慢性低度炎症加剧的骨关节炎中发挥修复软骨的作用[28]。骨关节炎发生后,关节GDF-5 表达显著下降,核因子κB (nuclear factor kappa-B,NF-κB)、白介素-6(nterleuikn-6,ⅠL-6) 和肿瘤坏死因子α (tumor necrosis factor-α,TNF-α)等炎症因子表达显著上升[29]。由上可知,GDF-5 可通过降低骨关节炎症因子水平,改善骨关节炎病症。除此以外,GDF-5可通过p38-MAPK减少心梗心脏纤维化程度[2]。当BMPR-ⅠⅠ/p38-MAPK 被阻断时,主动脉瓣出现钙化,无法正常抑制血液回流[30]。BMPRII基因突变会导致遗传性肺动脉高压发病风险增加,且该基因高甲基化也会增加发病几率[31-32],其具体机制与Smad1/5/8 和DNA 结合抑制因子1/3(inhibitor of DNA binding,ⅠD1/3) 无法经过受体磷酸化有关[33]。因此,GDF-5/BMPR-ⅠⅠ是组织器官如心血管系统疾病关键的治疗靶点。GDF-5在白色脂肪中激活p38-MAPK信号,调节白色脂肪细胞生长,调控细胞生长和能量代谢平衡[34],或在棕色脂肪中激活PⅠ3K/Akt 平衡脂肪能量代谢[35]等。综上,GDF-5/BMPR是调节机体代谢的关键信号,除激活Smads 信号通路外,还可激活p38-MAPK、PⅠ3K/Akt等信号通路作用于相应器官组织。
2 GDF-5与代谢性疾病
2.1 代谢性疾病定义
代谢性疾病不是特指某一疾病,而是指代谢异常引发机体某一器官或多个器官功能紊乱甚至出现病征。常见的代谢性疾病包括心血管疾病、慢性肾病、高血压、神经退行性疾病、骨质疏松、肌肉减少症和非酒精性脂肪肝等[4]。代谢性疾病的具体发病机制尚未明确,目前认为遗传和表观遗传、机体氧化和慢性低度炎症等是主要诱因和发展助力[36-38]。代谢性疾病的诱因如慢性低度炎症等其机制与氧化应激联系紧密[37,39]。以心血管疾病为例,心脏发生病征后促使巨噬细胞和中性粒细胞分泌大量炎性细胞因子,如白介素家族,并诱导单核细胞吸引趋化因子在组织中引发炎症,严重影响血管和心肌功能,导致发展成为动脉粥样硬化或心肌梗死等心血管疾病[40-41]。此外,线粒体出现功能障碍如线粒体自噬,是炎症和代谢性疾病发生的主要诱因之一。当线粒体损伤时,代谢器官无法正常获得氧供给或氧运输,组织氧代谢平衡打破[42]。此时损伤的线粒体产出过量ROS且无法及时清除,引发脂质过氧化、蛋白质折叠错误和DNA损伤等,导致能量代谢失调和细胞功能障碍,诱发炎症[43-44],而炎症反过来加剧了氧化应激,二者互为因果,最终致使机体代谢紊乱并诱发代谢性疾病[45]。不仅如此,慢性低度炎症作为人类的第一大杀手,还会导致诸如代谢性疾病、心血管疾病和神经退行性疾病等[46-47]。代谢性疾病本身具有巨大的危害,且如肥胖症和糖尿病等会引发心血管系统代谢障碍,最终诱发心血管疾病[48]。因此,除探究如何遏制代谢性疾病发生外,寻求继发的代谢性相关疾病的治疗靶点至关重要。
2.2 GDF-5与心血管疾病
据《中国心血管健康与疾病报告2021》概要显示,心血管疾病高居中国城乡居民死亡原因首位[49],开展心血管疾病的防治工作迫在眉睫。代谢性心血管疾病的发病机制包括氧化应激、慢性炎症等,其他器官代谢障碍也会引发代谢性心脏病如糖尿病心脏病,以上与线粒体功能障碍和葡萄糖代谢异常有关[50]。早期研究发现,GDF-5 在大鼠心脏中显著表达[51]。小鼠心肌梗死后,心肌GDF-5表达水平显著升高,敲除gdf5后显著抑制心肌p38-MAPK 磷酸化,氧化应激水平升高,ⅠⅠⅠ型胶原表达上调,左心室管壁血管减少,小动脉密度降低,心肌梗死瘢痕区域扩张、纤维化水平显著上升,心肌出现过度细胞凋亡,心功能严重受损[2]。上述研究表明GDF-5 具有护心脏保作用。该实验小鼠心脏敲除gdf5后,Smad1/5/8磷酸化水平显著上升。然而有实验证明,内皮细胞中的富亮氨酸α2 糖蛋白1 (leucine-rich-alpha-2-glycoprotein-1, LRG1)可介导Smad1/5/8 促进心脏血管生成[52-53]。由此推测,心脏敲除gdf5后可能导致其他BMP 家族蛋白或抑制性Smad 如Smad7 表达失调。研究发现,肺动脉高压小鼠敲除BmprII后,虽然不会加重遗传性肺动脉高压,但心肌细胞Ca2+瞬态、肌浆网Ca2+负荷减少,导致心肌细胞收缩功能障碍,此外,Smad1/5/8的磷酸化水平也显著降低[54]。且BmprII的丢失或突变会导致遗传性肺动脉高压发病的概率增加[32-33]。推测BMPR-ⅠⅠ在心肌细胞中主要发挥调控正常收缩的功能,并通过磷酸化Smads促进血管生成,GDF-5 是BMPR-ⅠⅠ的重要配体,因此GDF-5在心肌细胞中的功能需要深入探讨。GDF-5可促使X 连锁凋亡抑制蛋白(X-linked inhibitor of apoptosis protein,XⅠAP)与BMPR-ⅠB 结合,阻止泛素化介导的XⅠAP降解,显著降低半胱天冬酶3(Caspase-3)/多聚腺苷酸二磷酸核糖聚合酶(poly ADP-ribose polymerase,PARP)表达水平,抑制小鼠胚胎成纤维细胞(mouse embryo fibroblast cells,MEFs)凋亡[55],但该过程无法通过BMPR-ⅠⅠ抑制人血管平滑肌细胞(human umbilical vein smooth muscle cell,HUVSMC)凋亡。以上研究提示BMPR-ⅠⅠ在不同细胞中发挥不同作用。GDF-5/BMPR/Samd1 信号可上调ⅠD1/3,抑制p38-MAPK磷酸化,促进细胞外调节蛋白激酶(extracellular regulated protein kinases,ERK)磷酸化,降低细胞炎症水平,缓解氧化应激,调节HUVSMC 分化和增殖,促进血管生成[56]。另有研究证明,BMP-4作为配体激活BMPR-ⅠⅠ/Smad1/5/ⅠD1/3 信号,减少肺动脉高压小鼠心肌细胞功能障碍[33]。在人肺动脉内皮细胞和人微血管内皮细胞中,GDF-5能够通过ALK-3和BMPR-ⅠⅠ介导Smad1/5发挥促进血管生成作用[57]。因此,Smad1/5/8信号在心血管系统中发挥促进血管生成的作用。综上,BMPR-ⅠⅠ是心血管系统中重要的受体,而GDF-5 作为重要配体,与BMPR-ⅠⅠ结合后减少心脏氧化应激、纤维化和细胞凋亡,增加血管生成,延缓代谢性心脏病进程。GDF-5在心血管系统中发挥保护作用的机制尚缺乏深入研究,如GDF-5 是否能调节心脏中炎症因子水平或在心肌梗死中阻止心肌细胞凋亡等。对于未来临床治疗心肌梗死等心血管疾病方面,GDF-5有望成为新的靶标分子(图2)。

Fig. 2 The functional mechanism of GDF-5 in cardiovascular disease图2 GDF-5在心血管疾病中的功能机制
2.3 GDF-5与神经退行性疾病
GDF-5 同源家族中BMP-2、BMP-5、BMP-6和BMP-7 等成员是多巴胺能神经元中重要的神经营养因子[58]。GDF-5 作为BMP 家族的成员之一,在皮层、海马、中脑和后脑等脑区表达,通过激活Smads1/5/8信号调节神经元和胶质细胞的生长、迁移和分化,延缓帕金森病(Parkinson’s disease,PD)和阿尔茨海默病(Alzheimer’s disease,AD)等神经退行性疾病的发展进程[59-60]。文献表明,氧化应激和炎症等因素是引发PD 和AD 等神经退行性疾病的主要原因,且炎症的发生和过量的ROS会导致产生大量小胶质细胞和星形胶质细胞,这两种细胞的过度活化是神经退行性疾病的病理学标志,并加剧疾病进程[61-62]。皮下注射GDF-5 后,小鼠海马神经元损伤显著减少,其减少细胞凋亡的作用可能是由BMPR-ⅠB介导[63]。在向小鼠海马区注射GDF-5 和GDF-5 重组蛋白培养的神经元细胞实验中发现,BMPR/ Samd1/5/8信号显著激活,上调转录因子HES5(hairy/enhancer of split 5),对抗ROS 爆发诱导的氧化应激和线粒体损伤,改善神经元细胞炎症,增加未成熟神经元数量并促进海马区锥体细胞树突显著生长,有效改善小鼠创伤性脑损伤,延缓AD 的进程[64-65]。另有研究发现,GDF-5/BMPR-Ⅰ/Smad1/5/8 激活可促进复侧中脑多巴胺能神经元突触生长[66-67]。gdf5过表达小鼠大脑的核苷二磷酸激酶A (nucleoside diphosphate kinase 1,NDKA)和丝氨酸-苏氨酸受体相关蛋白激酶 (serine-threonine kinase receptor-associated protein,STRAP)显著增加,并促进体外SH-SY5Y神经细胞系中神经元树突生长[68]。综上,GDF-5在神经退行性疾病中主要发挥阻止氧化应激和炎症,激活BMPR/Smads信号促进神经元细胞和神经树突生长等作用,有效延缓AD和PD等疾病进程。GDF-5被认为是重要的神经营养因子之一,其有效的抗炎作用能抵御或延缓神经退行性疾病的发生发展,但尚缺乏关于GDF-5 在神经系统中发挥作用过程的研究。因此探究GDF-5 在神经退行性疾病中减少炎症和氧化应激的生物学机制是未来治疗的重要靶点(图3)。
2.4 GDF-5与代谢性骨病
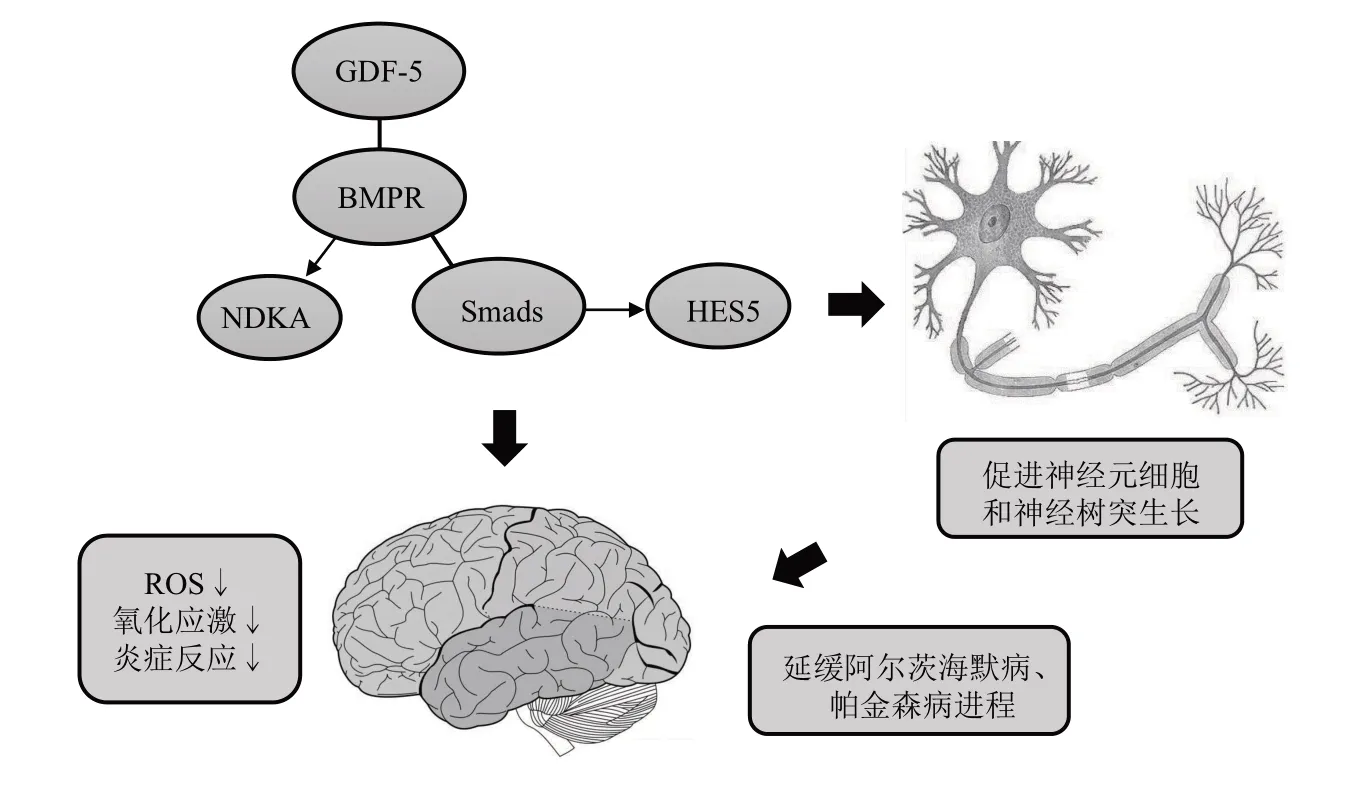
Fig. 3 The functional mechanism of GDF-5 in neurodegenerative disease图3 GDF-5在神经退行性疾病中的功能机制
常见代谢性骨病有骨关节炎、骨质疏松症和内分泌骨病等,其中骨关节炎是其典型代表,不仅严重影响中老年人生活质量,且通常伴有心血管疾病、糖尿病、高血压和肥胖症等至少一种共病[69]。慢性低度炎症是导致骨关节炎发生的主要原因[70]。GDF-5与关节软骨内环境稳态高度相关,在骨骼发育后期可促进软骨细胞增殖。GDF-5调节胚胎早期正常骨发育,软骨受损可诱导GDF-5 表达增加,促进自我修复。研究发现,骨关节炎相关位点gdf5-UQCC突变导致会关节软骨易感性增加,并显著阻碍人身高增长[71-72]。另一项全基因组研究也证明,gdf5是骨关节炎的易感基因[73]。临床证据显示,人gdf5突变会导致软骨发育不良[1],小鼠gdf5缺陷出现膝关节发育异常[74]。实验发现,由GDF-5和细胞外基质混合制成的功能性支架嵌入关节软骨缺陷的兔关节后,关节软骨出现再生且表现出更强的软骨细胞迁移和分化[75]。敲除小鼠BmprIb基因出现趾骨软骨细胞分化缺陷、骨细胞密度降低,短指发育畸形[20]。上述研究结果表明,GDF-5与BMPR是骨生长发育中的关键因子,通过与BMPR-ⅠB 结合调节软骨发育。骨关节炎发病机制可能与激活经典Wnt信号通路,上调基质金属蛋白酶13(matrix metalloprotease 13,MMP-13)表达有关。GDF-5 通过调节Dickkopf 相关蛋白1(Dickkopf-related protein 1,DKK-1)抑制经典Wnt信号通路,降低MMP-13表达,稳定软骨细胞外基质内环境并减少软骨细胞凋亡[76]。然而DKK-1 也具有诱导软骨细胞凋亡和软骨破坏的作用[77]。因此,DKK-1 具有两面性,GDF-5 调节经典Wnt 信号通路的作用仍需进一步探索。研究发现,GDF-5可上调miR-17的表达,通过降低MMP-2、MMP-3和MMP-13 等因子减轻骨关节炎[78],表明GDF-5通过降低MMPs 表达改善骨关节炎。阴阳蛋白(Yin Yang 1,YY-1)能够调节核仁小RNA 宿主基因5(small nucleolar RNA host gene 5,SNHG5),激活miR-212-3p 并靶向GDF-5/Smads,调节人骨髓间充质干细胞的成骨分化[79]。长期以来,GDF-5被认为是骨关节发育的重要因子,同时也是人生长发育的重要基因指标[80]。研究表明,给予抗炎药物地塞米松和GDF-5 有显著的抗炎作用并诱导骨髓间充质干细胞(bone mesenchymal stem cells,BMSC)向软骨细胞分化[81]。综上,GDF-5 通过激活下游通路,减少骨关节和软骨炎症因子表达,预防或延缓骨关节炎等代谢性骨病发展进程。目前GDF-5临床治疗的相关数据仍然缺乏,鉴于GDF-5在骨中发挥的重要作用,开展临床研究具有重要意义(图4)。
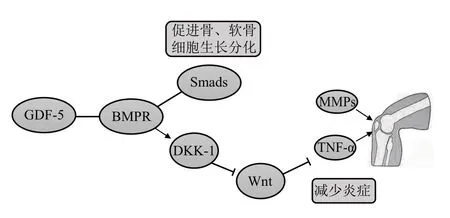
Fig. 4 The functional mechanism of GDF-5 in metabolic bone disease图4 GDF-5在代谢性骨病中的功能机制
2.5 GDF-5与肌肉减少症
骨骼肌作为人体最大器官之一,具有高度可塑性,在维持人体运动和身体平衡中发挥重要作用。GDF-5 是失神经支配肌肉的关键因子。研究发现,BMP及下游Smad1/5/8在肌肉维持、生长和运动神经元调控骨骼肌活动中发挥关键作用[82-84]。gdf5丢失小鼠骨骼肌失神经支配后,无法正常磷酸化Smad1/5/8 和ⅠD1,骨骼肌质量降低和横截面积减少,肌肉萎缩加剧[82]。Dachshund 同源基因2(Dachshund family transcription factor 2,DACH2)和组蛋白脱乙酰酶9 (histone deacetylase 9,HDAC9)在失神经支配肌肉中,通过调节gdf5表达,实现骨骼肌再神经支配[85]。肌细胞生成素(myogenin)基因上游区域的Myoparr敲除后,小鼠肌肉GDF-5 表达增加,Smad1/5/8 磷酸化水平上升,失神经支配肌肉的萎缩有所缓解[86]。综上,DACH2、HDAC9、Myoparr可调节GDF-5的表达,提高失神经支配肌肉疾病患者的运动能力。在DNA 甲基化转移酶3A (DNA methyltransferase 3A,DNMT3A) 缺失小鼠的肌肉卫星细胞中,GDF-5 启动子的DNA 甲基化水平下降,GDF-5 分泌增加,肌卫星细胞分化受到抑制,损伤肌肉的修复再生能力减弱[87]。敲除Dnmt3a显著增加GDF-5表达,抑制卫星细胞分化,老龄骨骼肌再生能力下降,骨骼肌再生受损[87]。上述结果表明,GDF-5在不同肌细胞中发挥的作用存在差异。电压门控钙通道CaVβ1E可促进下游GDF-5信号传导,改善小鼠失神经支配后的肌肉萎缩,而老龄小鼠肌肉CaVβ1E 量降低,过表达CaVβ1E 通过激活GDF5/Smad1/5/8信号显著改善老龄小鼠肌肉质量[88],表明gdf5受DNA 甲基化和电压门控钙通道的调节。因此认为,GDF-5是电压门钙通道离子流动的关键介质,在增龄性和失神经导致的肌肉减少症中通过激活Smad 信号,减少肌肉萎缩,增加肌肉质量(图5)。
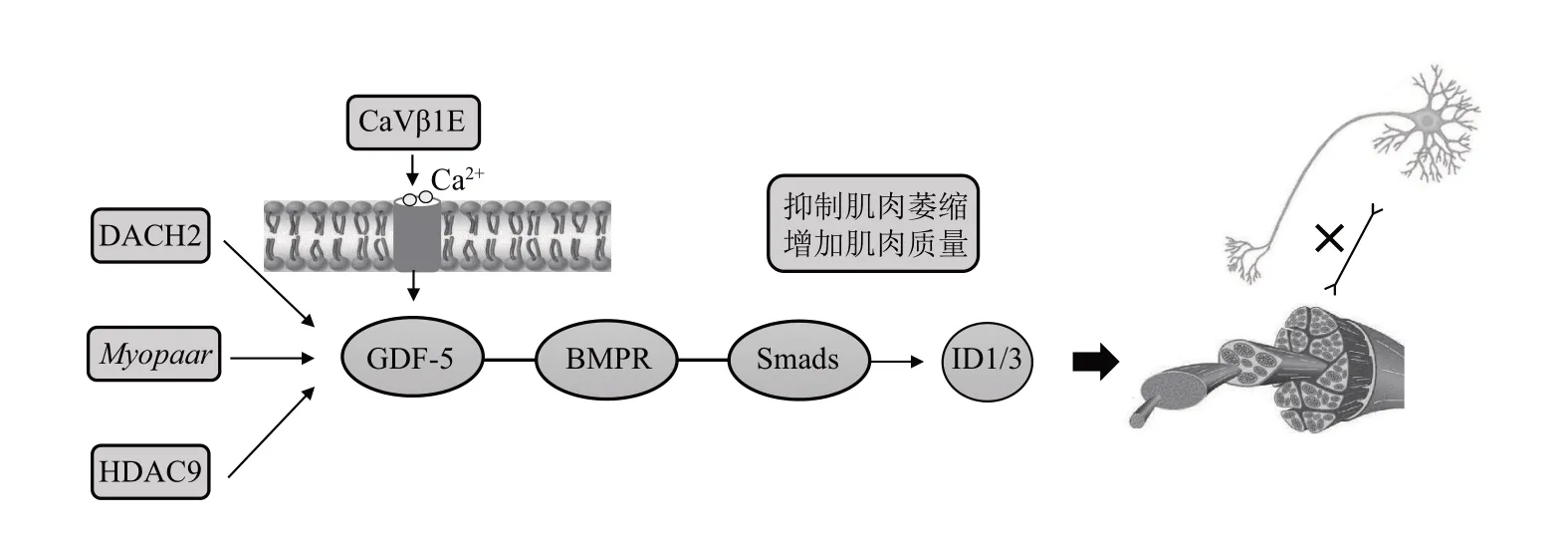
Fig. 5 The functional mechanism of GDF-5 in improving sarcopenia图5 GDF-5在肌肉减少症中的功能机制
2.6 GDF-5与代谢相关脂肪性肝病
MAFLD 发病机制复杂,主要是由肝脏细胞脂肪过度堆积引起,从肝脏脂肪变性进展为肝纤维化,具体发病机制与代谢应激和炎症的反复发生有关[89-90]。MAFLD 发生后,肝脏线粒体自噬严重,导致产生过量ROS,肝脏脂肪能量代谢失衡,并进一步促进炎症和纤维化发生发展[90]。研究表明,GDF-5可通过减少肝脏脂肪沉积和改善代谢,阻止MAFLD进程[91]。棕色脂肪组织在产热和调节能量平衡中起重要作用,肥胖症发病与棕色脂肪组织失衡有关。有文献报道,GDF-5/BMPR/Smad5/PGC-1α信号通路显著上调棕色脂肪组织的解偶联蛋白1(uncoupling protein 1,UCP-1)表达,增加机体能量消耗和产热能力,维持能量代谢稳态,降低饮食诱导的肥胖易感性[35]。另有研究发现,GDF-5 通过磷酸化PⅠ3K/Akt 信号,促进棕色脂肪生成[5],通过加速白色脂肪细胞周期进程并增加S期细胞数量,促进脂肪细胞分化[92]。GDF-5 还可激活p38-MAPK,提高胰岛素敏感性,增加白色脂肪产热,提高胰岛素敏感性[34]。最新研究发现,骨骼肌衍生外泌体通过miR-146a-5p靶向GDF-5/PPARγ调节脂肪组织脂质代谢[93]。以上研究结果表明,GDF-5可调节棕色脂肪代谢和白色脂肪细胞分化和能量代谢过程。综上,GDF-5 通过维持线粒体能量稳态,调节脂肪生成和能量消耗,降低肝脏发病风险,延缓MAFLD发展进程(图6)。
表1总结了GDF-5在不同组织器官及疾病中的变化和作用。
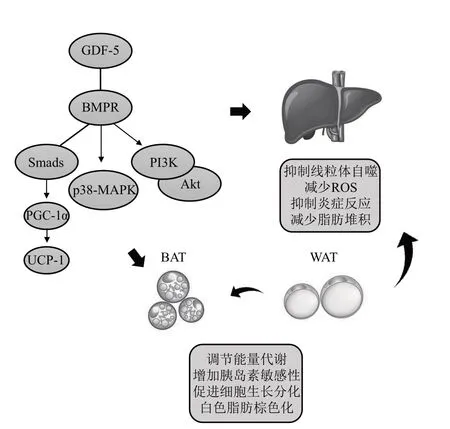
Fig. 6 The functional mechanism of GDF-5 in MAFLD图6 GDF-5在代谢相关脂肪性肝病中的功能机制

Table 1 Pathological phenotypes of different tissues and organs after GDF-5 interventing in different ways表1 GDF-5干预后不同组织器官的病理表型
3 总结与展望
GDF-5作为TGF-β家族成员之一,在骨生长发育中发挥关键作用,在心脏、大脑、肾脏、骨骼肌、肝脏和脂肪等多个器官组织中表达。炎症和氧化应激是代谢性疾病的核心环节,导致各个器官代谢紊乱并向代谢性疾病发展。GDF-5 通过Smads、p38-MAPK、PⅠ3K/Akt等信号,发挥促进细胞增殖分化、减少氧化应激和炎症以及细胞凋亡等生物学作用。近年来,除了对骨、关节、肌腱修复以及胚胎发育等研究外,GDF-5对其他器官组织的研究逐渐受到关注。临床和动物研究发现,通过注射GDF-5 或外源性刺激GDF-5 分泌增加等手段,对机体代谢产生积极作用。目前针对GDF-5 作用机制仅限基础研究,尚缺乏大量临床应用研究。本文通过梳理GDF-5 与代谢性疾病文献,总结GDF-5与下游信号,为GDF-5 在代谢性疾病治疗中的作用及其具体机制研究提供理论参考。GDF-5通过减少炎症和氧化应激改善代谢性疾病,有望成为相关疾病治疗的新靶点,其具体机制仍然需要深入研究。
猜你喜欢
中老年保健(2022年2期)2022-11-25
生命科学研究(2022年2期)2022-05-15
昆明医科大学学报(2022年1期)2022-02-28
中老年保健(2021年5期)2021-12-02
基层中医药(2021年9期)2021-06-05
中国临床医学影像杂志(2019年5期)2019-08-27
中国临床医学影像杂志(2019年1期)2019-04-25
老年医学与保健(2017年6期)2017-02-06
罕少疾病杂志(2016年4期)2016-03-11
中医研究(2014年11期)2014-03-11
