HBx蛋白抑制乙型肝炎病毒诱导肝细胞表达Ⅰ型干扰素的研究
2024-03-20 09:41潘颖杨凯陈谨孙蓓蓓田平平张发苏安徽医学高等专科学校医学技术学院合肥3060安徽医科大学第二附属医院检验科合肥3060
中国免疫学杂志 2024年2期
潘颖 杨凯 陈谨 孙蓓蓓 田平平 张发苏 (.安徽医学高等专科学校医学技术学院,合肥 3060;.安徽医科大学第二附属医院检验科,合肥 3060)
Ⅰ型干扰素主要包括IFN-α和IFN-β,在病毒感染期间,宿主对病毒入侵的初始反应是诱导Ⅰ型干扰素产生并与其受体(interferon-α/β receptor,IFNAR)结合,激活酪氨酸蛋白激酶和信号转录与转录激活因子(Janus kinase-signal transducers and activators of transcription,JAK/STAT)信号通路,合成多种抗病毒蛋白,如:核糖核酸依赖性蛋白激酶(protein kinase RNA dependent,PKR)、2'-5'寡聚腺苷酸合成酶(2'-5'oligoadenylate synthetase,OAS)、黏病毒抗性蛋白A(myxovirus resistance protein A,MxA),直接抑制病毒复制[1-2]。除了直接的抗病毒作用外,IFN-α还可以强烈调节宿主的先天性和适应性免疫反应[3-4],如:增强NK细胞的增殖、CD8+T细胞和巨噬细胞的细胞毒性以及IFN-γ分泌等,以增强宿主对病原体的清除能力,因此,Ⅰ型干扰素信号通路激活是宿主早期重要的自然防御机制,在抑制病毒入侵方面发挥重要作用。然而,WIELAND等[5]对实验室感染HBV的黑猩猩进行体内研究表明,HBV不能诱导干扰素刺激基因(interferon-stimulated gene,ISG)的表达,所以可以在肝脏中持续复制;SUSLOV等[6]通肝脏活检发现,HBV感染患者肝组织中IFN和ISG的表达被抑制,使得病毒不能被有效清除;本课题组亦在前期研究中发现,由HBV编码的HBx蛋白可以抑制外源性Ⅰ型干扰素诱导抗病毒蛋白MxA的合成[7]。据此,本课题组推测HBV很有可能通过损伤Ⅰ型干扰素信号通路,影响机体的先天免疫应答,从而引起病毒感染的慢性化。为此,本课题组拟研究HBV对肝细胞中Ⅰ型干扰素表达的影响,并进一步探讨可能存在的机制,为揭示HBV慢性感染与宿主免疫功能紊乱的关系提供新的思路。
1 材料与方法
1.1 材料 人肝癌HepG2和HepG2.2.15细胞由安徽医学高等专科学校生化实验室保存;细胞培养基DMEM和胎牛血清购自美国Gibco公司;Lipofectamine 3000TM转染试剂盒、ECL超敏发光试剂盒购自美国Thermo公司;TRIzol购自美国Life technogies公司;氯仿、异丙醇及乙醇均为分析纯;反转录试剂盒购自大连TaKaRa公司;荧光染料购自中国Novoprotein公司;BCA试剂盒、SDS-PAGE凝胶配制试剂盒均购自碧云天试剂公司;mock-siRNA、HBxsiRNA及引物由上海生工生物合成;HBx、p-IRF3及GAPDH抗体购自美国Santa Cruz公司;pEGFP-N1和pEGFP-HBx质粒由南京大学附属鼓楼医院刘巧玉博士惠赠;荧光显微镜购自德国Leica公司;荧光定量PCR仪购自美国Thermo Scientific公司;电泳仪、电泳槽购自上海天能科技有限公司;自动曝光仪购自培清科技有限公司。
1.2 方法
1.2.1 细胞培养及转染 HepG2和HepG2.2.15细胞用含10%胎牛血清的DMEM于37 ℃、5%CO2培养箱中常规培养,取对数生长期的细胞以2×105个/ml密度接种于6孔板用于转染,待细胞融合度达到80%~90%时候,改用无胎牛血清DMEM培养,以Lipofectamine 3000TM转染试剂将mock-siRNA和HBxsiRNA转染至HepG2.2.15细胞,48 h后收集细胞沉淀检测HBx蛋白表达水平;将带绿色荧光蛋白的pEGFP-N1和pEGFP-HBx质粒转染至HepG2细胞,在荧光显微镜下观察HBx蛋白的表达。
1.2.2 荧光定量PCR检测Ⅰ型干扰素IFN-α和IFN-β mRNA含量 收集转染前后的细胞沉淀,加入1 ml TRIzol完全裂解,再加入0.2 ml氯仿振荡离心取上清,加入0.5 ml预冷的异丙醇,-20 ℃孵育30 min后离心去上清,最后加入1 ml预冷的75%乙醇,12 000 r/min离心去上清,室温干燥RNA沉淀,按照TaKaRa反转录试剂盒说明书反转录为cDNA。以cDNA为荧光定量的模板,运用SYBR Green PCR试剂盒进行扩增反应,PCR仪设置扩增程序为:95 ℃ 1 min,95 ℃ 20 s,60 ℃ 1 min,40个循环,72 ℃20 s。目的mRNA相对表达量的计算方法为2-ΔΔCt。所用引物序列见表1。

表1 引物序列Tab.1 Primer sequences
1.2.3 Western blot检测HBx及p-IRF3蛋白含量使用RIPA细胞裂解液细胞,并以12 000 r/min离心10 min收集上清液,即含有细胞总蛋白,BCA法检测蛋白浓度。在收集的蛋白样品中按照1∶4加入蛋白上样缓冲液,经沸水浴加热10 min以充分变性蛋白质,待样品冷却至室温后,上样至加样孔内行SDSPAGE蛋白电泳1.5 h、转膜、封闭,一抗4 ℃孵育过夜,二抗室温孵育2 h。采用ECL超敏发光试剂检测目的蛋白。
1.3 统计学分析 采用MedCalc10.4软件进行数据统计分析,两组数据比较采用独立样本t检验,P<0.05表示差异具有统计学意义。
2 结果
2.1 HBV对肝细胞中Ⅰ型干扰素表达的影响 荧光定量PCR分析显示,与HepG2细胞相比,转染HBV全基因组的HepG2.2.15细胞中IFN-α和IFN-β的mRNA水平升高,但差异不具有统计学意义(P>0.05,图1),提示HBV在体外细胞中并未充分诱导Ⅰ型干扰素的表达。

图1 Ⅰ型干扰素在HepG2和HepG2.2.15细胞中的表达Fig.1 Expression of typeⅠinterferon in HepG2 and HepG2.2.15 cells
2.2 HBx-siRNA上调细胞中IFN-α和IFN-β mRNA含量 将mock-siRNA和HBx-siRNA分别转染至HepG2.2.15细胞,结果显示,与未经处理的HepG2.2.15细胞相比,转染HBx-siRNA的细胞中HBx蛋白表达显著降低(P<0.01,图2A),而IFN-α和IFN-β mRNA含量显著升高(P<0.01或P<0.001,图2B),提示HBV可通过HBx抑制Ⅰ型干扰素信号通路激活。
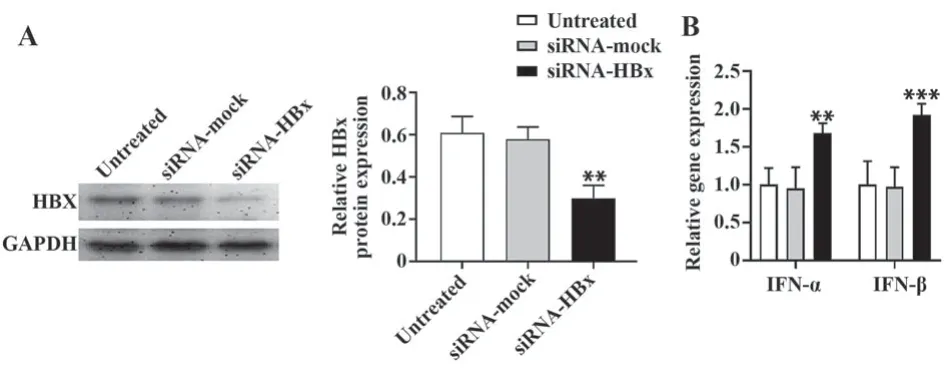
图2 HBx-siRNA对HepG2.2.15细胞内IFN-α和IFN-β表达的影响Fig.2 Effects of HBx-siRNA on expressions of IFN-α and IFN-β in HepG2.2.15 cells
2.3 pEGFP-HBx通过抑制IRF3的磷酸化影响Ⅰ型干扰素信号通路 将HepG2细胞分别转染空载体质粒pEGFP-N1和pEGFP-HBx,48 h后运用荧光显微镜观察HBx蛋白在细胞中的表达(图3A)。进一步分析各转染细胞中Ⅰ型干扰素mRNA及p-IRF3蛋白的表达,结果显示,与空载体转染的HepG2细胞相比,转染pEGFP-HBx质粒的细胞中p-IRF3蛋白含量降低,IFN-α和IFN-β mRNA水平降低,差异具有统计学意义(P<0.01,图3B、C)。

图3 HBx蛋白影响HepG2细胞p-IRF3和Ⅰ型干扰素表达Fig.3 HBx protein affects expressions of p-IRF3 and typeⅠ interferon in HepG2 cells
3 讨论
HBV是一种非细胞病变的嗜肝DNA病毒,可编码多种病毒蛋白,如:DNA多聚酶、表面抗原(HBsAg)、核心抗原(HBcAg)和HBx蛋白[8-9]。肝脏急性感染HBV后,90%~95%的患者能够清除感染并完全康复,其余5%~10%的患者发展为慢性HBV感染,并且部分逐步发展为肝硬化和肝细胞癌[10]。WHO估计全世界约有2.57亿人呈慢性HBV感染,每年有近100万人死于HBV相关并发症[11]。一般认为,HBV感染的结局取决于病毒与宿主免疫系统的相互作用,研究表明,HBV会抑制宿主的免疫系统,表现为先天和适应性免疫细胞功能障碍,而Ⅰ型干扰素在体内具有调节先天免疫反应和激活后天免疫反应的作用[12-13]。此外,尽管常规和聚乙二醇化的IFN-α蛋白已成为慢性乙型肝炎(chronic hepatitis B,CHB)患者的一线治疗药物,且Ⅰ型干扰素的抗病毒作用已明确,但在CHB患者中IFN-α治疗的病毒清除率相当低,具体机制尚未明确[14-15],因此推测HBV可能影响了Ⅰ型干扰素信号通路。为此,本实验通过对比HepG2细胞和转染HBV全基因组的HepG2.2.15细胞中IFN-α和IFN-β的基因表达量,证实HBV并不能充分诱导肝细胞表达Ⅰ型干扰素,进一步将HBx-siRNA转染至HepG2.2.15细胞发现,抑制肝细胞在病毒转录中维持共价闭合环状DNA(covalently closed circular DNA,cccDNA)微型染色体的重要调节蛋白—HBx蛋白的表达,可上调IFN-α和IFN-β的mRNA水平,提示HBx可能抑制了HBV诱导肝细胞表达Ⅰ型干扰素。
Ⅰ型干扰素的产生受到干扰素调节因子(interferon regulatory factors,IRFs)家族的调控,其中,IRF3是Ⅰ型干扰素信号通路的主要调节因子,机体感染病毒后,可通过模式识别受体对病毒病原体相关分子模式进行识别,导致下游转录因子IRF-3的激活和核转位,从而诱导IFN-α和IFN-β的表达[16-17]。而在HBV持续感染的肝细胞中,模式识别受体及IRF3呈低表达,LEE等[18]通过将IRF3小分子激活剂处理HBV感染的细胞,发现激活IRF3可诱导先天免疫作用,抑制HBV cccDNA的形成。在本研究中,将HBV编码的HBx基因表达质粒转染至HepG2细胞,检测结果显示,过表达HBx蛋白的HepG2细胞中IFN-α和IFN-β mRNA表达水平均下降,且磷酸化的IRF3蛋白含量亦降低。
综上所述,本研究通过对培养的肝细胞模型进行HBx基因过表达和沉默,揭示了HBV通过其所编码表达的HBx蛋白抑制IRF3蛋白的活化,从而抑制Ⅰ型干扰素的表达,本实验为进一步探讨HBV感染的慢性化机制以及解决临床CHB患者Ⅰ型干扰素治疗应答不佳提供了新的思路。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
癌变·畸变·突变(2016年3期)2016-02-27
中国继续医学教育(2015年6期)2016-01-07
实用皮肤病学杂志(2015年4期)2015-12-22
哈尔滨医药(2015年6期)2015-12-01
哈尔滨医药(2015年4期)2015-12-01
中国医疗美容(2015年2期)2015-07-19
医学研究杂志(2015年12期)2015-06-10