灰毡毛忍冬bZIP25基因的克隆及其表达模式分析
2024-03-14 03:48王珊曾娟谢瑜周日宝刘湘丹童巧珍龙雨青陈言刘小丽湖南中医药大学第二附属医院长沙4000湖南中医药大学药学院长沙40208湘产大宗道地药材种质资源及规范化种植重点研究室长沙40208湖南省普通高等学校中药现代化研究重点实验室长沙40208焦作市产品质量检验检测中心河南焦作44000
中南药学 2024年2期
王珊,曾娟,谢瑜,周日宝,刘湘丹,童巧珍,龙雨青,陈言,刘小丽*(.湖南中医药大学第二附属医院,长沙 4000;2.湖南中医药大学药学院,长沙 40208;3.湘产大宗道地药材种质资源及规范化种植重点研究室,长沙 40208;4.湖南省普通高等学校中药现代化研究重点实验室,长沙 40208;.焦作市产品质量检验检测中心,河南 焦作 44000)
灰毡毛忍冬LoniceramacranthoidesHand.-Mazz.是中药材山银花的主要来源,具有清热解毒,疏散风热的功效[1],是治疗内痈、外痈之要药。随着生物技术的快速发展,灰毡毛忍冬分子水平的相关研究也越来越多。其中对灰毡毛忍冬的基因研究多集中在参与其活性成分绿原酸生物合成[2-4]、黄酮类化合物合成[5-6]、植物激素乙烯合成[7-8]以及其花冠特殊表型等方面[9-11],而关于其转录因子的相关研究报道则很少。
bZIP(basic leucine zipper)转录因子是植物中非常重要且常见的一类生物蛋白,由高度保守的碱性区和相对保守的亮氨酸拉链区组成。bZIP 转录因子通过与靶基因的启动子区域结合,激活或抑制其转录,从而调控基因的表达,参与植物各种生物学过程,在植物的生长发育、次生代谢产物合成以及抵御逆境胁迫等方面都发挥着不可替代的作用[12-13],如 bZIP 转录因子可以加速马铃薯生殖块茎的生长和衰老[14]。苹果MdbZIP44通过增加MdMYB1与下游靶基因启动子的结合,促进了脱落酸诱导的花青素生物合成[15]。陆地棉GhVIP1过表达后,提高种子发芽率和改善根系发育,使其耐旱性增强[16]。有研究对金银花五个花期的转录因子表达量进行差异分析,发现 49 个 bZIP 转录因子的表达量存在明显差异,提示其可能参与金银花的发育[17]。
本研究利用课题组前期研究获得的灰毡毛忍冬转录组测序数据,筛选出在花不同发育阶段具有显著表达差异的bZIP25(log2Fold Change:2.12,P<0.05),对其进行克隆及生信分析,并对其在不同器官、不同开花阶段的表达量进行研究分析,为后续深入探索灰毡毛忍冬bZIP25在花发育方面的功能研究奠定基础。
1 材料
1.1 药材
灰毡毛忍冬花、叶及茎在湖南隆回采摘,经湖南中医药大学周日宝教授鉴定为灰毡毛忍冬的花、叶及茎,将采集的花样品按不同花期分成七个阶段(f1~f7)[5]。将样品于封口袋中密封,装入液氮罐中,带回实验室于-80℃冰箱保存。
1.2 试药
多糖多酚植物总RNA提取试剂盒(杭州博日科技有限公司);RevertAid First Strand cDNA synthesis试剂盒(美国Thermo Fisher公司);快速琼脂糖凝胶DNA回收试剂盒、TS-GelRed Ver.2 1000 x in Water、Trelief 5α感受态细胞(北京擎科新业生物技术有限公司);SMARTer RACE 5'/3'试剂盒(日本Takara公司);pEASY-Blunt Cloning试剂盒、pEASY-T1 Cloning试剂盒、TranStart Green qPCR SuperMix UDG(北京全式金生物科技有限公司);2×Taq Master Mix(Dye)(康为世纪科技有限公司)。所用引物均由生工生物工程(上海)股份有限公司合成,引物序列如表1所示,浓度为10 μmol·L-1。

表1 引物序列Tab 1 Primer sequence
2 方法与结果
2.1 bZIP25基因克隆
2.1.1bZIP25基因核心片段扩增 取适量灰毡毛忍冬花蕾,根据多糖多酚植物总 RNA 提取试剂盒的方法提取灰毡毛忍冬总 RNA,琼脂糖凝胶电泳和核酸蛋白分析仪检测合格后,参照 RevertAid First Strand cDNA synthesis试剂盒的步骤,用提取得到的RNA 逆转录合成灰毡毛忍冬 cDNA。根据前期的转录组测序结果,筛选出标注为 bZIP 的序列,设计引物bZIP25-S 和bZIP25-A(见表1),以上述步骤得到的cDNA为模板进行PCR扩增。PCR 体系共25 μL,其中cDNA 1 μL,2×Taq Master Mix(Dye)12.5 μL,bZIP25-S 和bZIP25-A 各 1 μL,ddH2O 9.5 μL。PCR 扩增条件为 94℃ 5 min;94℃ 30 s,55℃30 s,72℃,90 s,40 个循环;72℃ 10 min。PCR产物用琼脂糖凝胶进行电泳分离,凝胶回收试剂盒回收目的条带,与 pEASY-T1 克隆载体连接,转化至感受态细胞后,涂抹于 LB 固体培养基进行蓝白斑筛选,挑取出白斑进行菌落 PCR,得到约 300 bp的目的条带(见图 1A),送至生工生物工程(上海)股份有限公司测序,明确核心片段序列长为 286 bp。
2.1.2bZIP25基因RACE扩增 根据上述测序结果,设计 RACE 引物bZIP25-5'和bZIP25-3'(见表1)。参照 RACE试剂盒的步骤分别获得bZIP25的 5'和 3'RACE-Ready cDNA。PCR体系包括5' 或 3' RACEReady cDNA 2.5 μL,Seq Amp DNA Polymerase 1 μL,bZIP25-5'或bZIP25-3'引物 1 μL,10×UPM 5 μL,PCR-Grade H2O补足 50 μL,以及 2×Seq Amp Buffer 25 μL。PCR 扩增条件为 94℃ 30 s,72℃ 2 min,5 个循环;94℃ 30 s,60℃ 30 s,72℃ 2 min,5 个循环;94℃ 30 s,58℃ 30 s,72℃ 2 min,40 个循环。回收、连接、克隆与测序等步骤同“2.1.1”项下,其中所用载体为 pEASY-Blunt 克隆载体。5'-端和3'-端RACE扩增结果显示,分别在400 bp 和600 bp 左右有一明亮条带(见图 1B 和 1C)。
2.1.3bZIP25基因cDNA全长获得及验证 应用序列拼接软件对 5'-端和 3'-端进行拼接,获得灰毡毛忍冬bZIP25基因cDNA序列全长。根据全长序列设计验证引物Y-bZIP25-F和Y-bZIP25-R(见表1),以“2.1.1”项下所得cDNA为模板进行全长验证。PCR体系共包括cDNA 1 μL,Y-bZIP25-F和Y-bZIP25-R各 1 μL,2×Pfu Master Mix(Dye)12.5 μL,以及ddH2O 补足 25 μL。PCR 扩增条件为 94℃ 5 min;94℃ 30 s,60℃ 30 s,72℃ 90 s,40 个循环;72℃10 min。回收、连接、克隆与测序等步骤同“2.1.1”项下,其中所用载体为 pEASY-Blunt克隆载体。将5'-端、3'-端及核心片段序列的测序结果用 Conting Express 软件拼接后,得到bZIP25基因cDNA全长序列共 892 bp。验证 cDNA全长,发现在 900 bp 左右一明显亮带(见图1D),经过回收、连接、克隆及测序,证实测序结果与拼接全长序列一致。
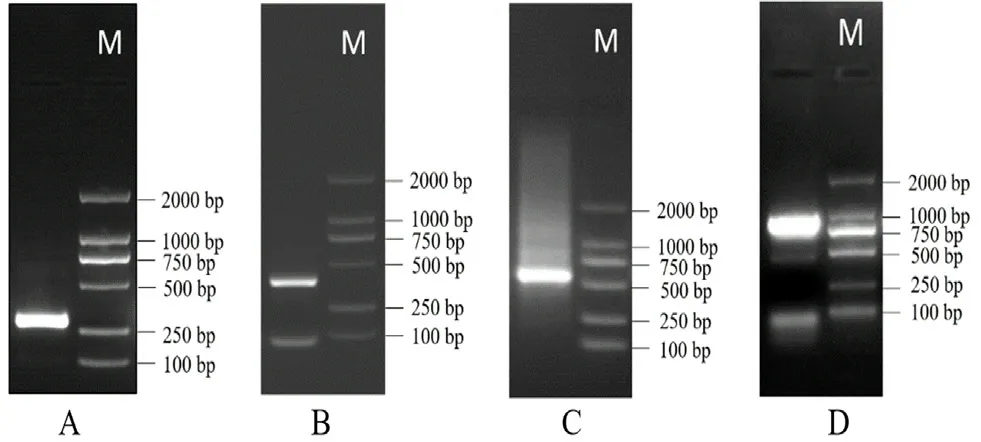
图1 bZIP25 基因PCR产物Fig 1 PCR product of bZIP25 gene
2.2 bZIP25 基因生物信息学分析
对灰毡毛忍冬bZIP25进行生物信息学分析所用的在线软件及具体网址见表2。

表2 生物信息学分析方法Tab 2 Bioinformatics analysis method
2.2.1 LmbZIP25 蛋白理化性质分析 灰毡毛忍冬bZIP25基因全长为892 bp,包含的ORF 区长度为576 bp,编码191个氨基酸,命名为LmbZIP25,其GenBank 登录号为:OR551766。
LmbZIP25蛋白分子式为C966H1525N305O295S7,相对分子质量为 22.35 kDa,理论等电点为 9.99。LmbZIP25蛋白的 191个氨基酸残基中,精氨酸(Arg)和丝氨酸(Ser)含量最高,为11.5%;络氨酸(Tyr)含量最低,仅 0.5%。LmbZIP25 蛋白不稳定系数为 72.09,大于阈值 40,属于不稳定蛋白。LmbZIP25 蛋白的脂溶系数为 63.25,总平均疏水指数(GRAVY)为 -1.007,为亲水性蛋白。通过 ProtScale 软件分析,该蛋白亲水/疏水氨基酸分布负值明显大于正值,提示亲水区占比远大于疏水区,与ProtParam 预测的结果一致。
经 WOLF PSORT 在线分析,该蛋白定位于细胞核中。利用 TMHMM 2.0 在线分析,其编码的氨基酸均在膜外,不具跨膜区域。通过 Signal P 4.1 Server 软件可知该蛋白无信号肽序列,为非分泌蛋白。NetPhos 3.1 在线软件分析提示,LmbZIP25中共有 28 个潜在磷酸化位点,其中有 22 个 Ser磷酸化位点、5 个 Thr 磷酸化位点及 1 个 Tyr 磷酸化位点。
2.2.2 bZIP25 蛋白结构分析 通过 CD-Search 在线分析,发现灰毡毛忍冬 bZIP25 蛋白在第 81~132 位氨基酸残基上含有 bZIP_plant_GBF1 保守结构域(见图2),提示其属于 bZIP 家族,与家族中其他成员具有相同或相似的功能。

图2 LmbZIP25 蛋白结构域分析Fig 2 Domain prediction of LmbZIP25 proteins
SOPMA在线预测LmbZIP25 蛋白二级结构。α螺旋为其主要结构组成元件,占46.07%;其次为无规则卷曲,占41.88%。通过SWISS-MODEL对LmbZIP25 蛋白进行同源建模,发现其与阿拉伯咖啡的bZIP蛋白在三级结构水平的相似度为53%,三级结构的覆盖率达 99%,且以α螺旋和无规则卷曲为主,与二级结构预测结果相符,结果见图 3。

图3 LmbZIP25 蛋白三级结构预测Fig 3 Prediction of tertiary structure of LmbZIP25 proteins
2.2.3 氨基酸序列同源性比对、进化分析和基序分析 将LmbZIP25 编码的氨基酸序列与 NCBI-Blast中的植物蛋白数据库进行搜索比对,发现与可可Theobromacacao、哥伦比亚锦葵Herraniaumbratica、小蓬草Erigeroncanadensis、莴苣Lactucasativa、狭叶油茶Camellialanceoleosa和甜橙Citrussinensis等的同源性较高,下载上述植物的氨基酸序列,使用 DNAMAN 软件进行多序列比对,结果见图4。这些植物中均包含有符合 bZIP 转录因子的“N-X7-R-X9-L-X6-L-X6-L”特征结构,且碱性区的同源性非常高,亮氨酸拉链区相对稍低。

图4 LmbZIP25 与其他植物bZIP 蛋白的多序列比对Fig 4 Multiple sequence alignment of LmbZIP25 with bZIP proteins of other plants
将在 NCBI-Blast 上下载的部分植物bZIP基因家族氨基酸序列,利用 MEGA 11.0 软件构建系统进化树。由图 5A 可知,进化树分为三支,灰毡毛忍冬LmbZIP25与芝麻SibZIP4、木犀榄OebZIP4、旋蒴苣苔DhbZIP5、小蓬草EcbZIP4、刺苞菜蓟CcbZIP4、莴苣LsbZIP4和除虫菊TcbZIP4聚为一支,说明与这些植物的亲缘关系较近。茄科的3种植物,渐狭叶烟草NabZIP11、马铃薯StbZIP63和辣椒CabZIP11聚为一支,其余植物为另一支。
对参与分析植物的蛋白序列进行保守基序分析(见图 5B),结果发现所有序列均具有 Motif 1、Motif 3、Motif 4 及 Motif 6 等4个保守基序,而不同进化分支 bZIP 的基序组成有较明显的种属特性,如第一大类均不含 Motif 8,第二大类的3个茄科植物NabZIP11、CabZIP11和StbZIP63均无 Motif 7,但具有 Motif 9。另外,同一种属的植物保守基序也呈现出个体差异,如第三大类中的菊科植物EcbZIP4无Motif 8,同为菊科的CcbZIP4则无 Motif 5 和 Motif 7。

图5 LmbZIP25 蛋白的系统进化树(A)和基序(B)Fig 5 Phylogenetic tree(A)and motif(B)of LmbZIP25 proteins
2.3 bZIP25 基因的表达分析
基于bZIP25基因的全长序列,设计实时荧光定量PCR的引物Q-bZIP25-F和Q-bZIP25-R(见表2)。提取灰毡毛忍冬七个花期的花及茎、叶的总 RNA,定量后逆转录成 cDNA。将18SrRNA基因作为内参,设计引物18S-F、18S-R(见表 2)进行 qRT-PCR。反应体系包括 2×TransStart Green qPCR SuperMix UDG 10 μL,上、下游引物各 0.4 μL,cDNA 1 μL,ddH2O补足 20 μL。扩增条件为 50℃ 2 min,94℃ 10 min;94℃ 5 s,55℃ 15 s,72℃ 10s,40 个循环。重复 3次,进行熔解曲线分析,运用2-ΔΔCt法计算相对表达量,结果见图6。不同花期中,LmbZIP25在黄色花蕾期的相对表达量最高,显著高于其他花期;不同器官中,以花期黄色花蕾期为参照,LmbZIP25相对表达量在茎中最高,其次为花期黄色花蕾期,而在叶中最低。其中,在茎中的相对表达量为叶的20.67倍,为黄色花蕾期的2.07倍。

图6 LmbZIP25 基因相对表达量Fig 6 Relative expression level of LmbZIP25 gene
3 讨论
bZIP 家族作为最大的转录因子家族之一,具有多种生物学功能,广泛地参与到植物的生命活动中。本研究为探索其在灰毡毛忍冬花器官发育中的功能克隆了bZIP25基因序列全长,开放阅读框(ORF)为 576 bp,编码 191个氨基酸,预测其为不稳定亲水蛋白,无跨膜结构,为非分泌蛋白。研究显示,bZIP 主要定位于细胞核中[18],LmbZIP25 蛋白分析结果与之相符。结构域分析其含有 bZIP_plant_GBF1 保守结构域,推测其属于 GBF1 类 bZIP 转录因子,与蓖麻[19]、橡胶草[20]和野菊[21]等 bZIP 编码的结构域一致。同源性比对结果表明,同源性较高的区域主要位于保守结构域上。与灰毡毛忍冬LmbZIP25进化关系较近的芝麻、木犀榄、旋蒴苣苔、莴苣和除虫菊等的 bZIP 蛋白在基序组成上高度保守,均含有Motif 1~Motif 8。
灰毡毛忍冬的入药部位为花蕾或带初开的花,为研究bZIP25在灰毡毛忍冬开花进程中的表达量差异,课题组将整个花期细分为七个阶段。qRTPCR 结果显示,LmbZIP25在灰毡毛忍冬七个花期、茎和叶中均有表达,表达量为茎>黄色花蕾期>叶,但表达量存在时空差异与组织特异性。bZIP 家族在不同的物种中具有不同的组织表达模式。如金银花LjbZIP1表达量在叶中高于成熟花中[22]。牡丹PobZIP1在茎和叶中的表达量基本持平,在花芽中表达量最少[23]。红花CtbZIP47在茎中的表达量高于花中,在叶中则最低[24]。
LmbZIP25在灰毡毛忍冬不同花期的表达水平整体呈先升后降的趋势,其中黄色花蕾期的LmbZIP25表达量显著高于其他花期。黄色花蕾期是灰毡毛忍冬开花前的最后阶段,此时的LmbZIP25表达量突增,提示其很可能促进灰毡毛忍冬花的开放。在其他植物中,也发现不少此类现象。如大豆GmbZIP33在拟南芥中过表达后,转基因拟南芥出现早花,并使FT等开花基因的表达量升高[25]。bZIP 转录因子GmFDL19能与开花基因GmFT2a和GmFT5a相互作用,GmFDL19在大豆中过表达时,能上调开花基因的转录水平,导致大豆出现早花[26]。水稻OsbZIP62不仅能够调控水稻的成花转变,还参与调控分枝、幼穗、花分生组织以及花器官的发育[27]。后续可通过获得LmbZIP25基因的过表达植株或功能缺失突变株,为深入研究其对灰毡毛忍冬开花的调控机制及验证其生物学功能提供参考。
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2021年15期)2021-10-14
今日农业(2020年24期)2020-12-15
兽医导刊(2016年12期)2016-05-17
法医学杂志(2015年4期)2016-01-06
法医学杂志(2015年4期)2016-01-06
传奇故事(破茧成蝶)(2015年8期)2015-02-28
火花(2015年7期)2015-02-27
现代检验医学杂志(2015年4期)2015-02-06