
Development and identification of two novel wheat-rye 6R derivative lines with adult-plant resistance to powdery mildew and high-yielding potential
2024-03-07 01:51GuohoHnJingWngHnwenYnTintinGuLijunCoShiyuLiuXiuqunLiYilinZhouJieruFnZhipengShiHongLiuLihuiLiDioguoAn
The Crop Journal 2024年1期
Guoho Hn, Jing Wng, Hnwen Yn, Tintin Gu, Lijun Co, Shiyu Liu, Xiuqun Li, Yilin Zhou,Jieru Fn, Zhipeng Shi, Hong Liu, Lihui Li,*, Dioguo An,*
a Center for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Shijiazhuang 050021, Hebei, China
b The National Key Facility for Crop Gene Resources and Genetic Improvement, Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081, China
c The State Key Laboratory for Biology of Plant Disease and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193, China
Keywords:Powdery mildew Secale cereale Triticum aestivum Wheat-rye 6R derivatives Agronomic performance
ABSTRACT
1.Introduction
Powdery mildew, caused by Blumeria graminis f.sp.tritici (Bgt),is a major threat to wheat (Triticum aestivum L.) production that can lead to a yield reduction of 5%–40%[1].Breeding resistant cultivars is crucial for controlling powdery mildew epidemics.To date,more than 80 formally designated powdery mildew(Pm)genes/alleles (i.e., Pm1-Pm69, Pm8 = Pm17, Pm18 = Pm1c, Pm22 = Pm1e,
Pm23 = Pm4c, and Pm31 = Pm21) have been identified in common wheat and its relatives [2–4].However, two challenges have been identified concerning the use of powdery mildew resistance which include: 1) most Pm genes confer all-stage resistance (ASR) which can be easily overcome by the highly variable Bgt isolates after extended periods in production.In this regard, adult-plant resistance(APR)conferred by quantitative resistance loci could be better than ASR, which is mostly conferred by qualitative resistance locus [5–7]; 2) most of the effective Pm genes have linkage drags and/or pleiotropic effects,which limits their large-scale utilization in wheat breeding [8,9].Therefore, it is critical to develop highyielding germplasm lines with stable and durable resistance.
Wheat relatives provide a precious resource for genetic improvement.Among the various wheat relatives, rye (Secale cereale L., 2n = 2x = 14, RR) has been an important gene donor for improving disease resistance in wheat [10].One breakthrough germplasm was the wheat-rye T1RS∙1BL translocation line.The chromosome 1RS carried multiple disease resistance genes including Pm8,Yr9,Lr26,and Sr31,as well as genes associated with superior yielding ability [11–15].From the 1980s to 1990s, the T1RS∙1BL translocation has been used in approximately 70% of wheat cultivars released in southwestern China and 55% in northern China [15].
Octoploid triticale (2n = 8x = 56, AABBDDRR), which was produced from a cross of hexaploid wheat and rye, possesses abundant genes associated with disease resistance, biotic or abiotic stress tolerance, and superior yield.It is more convenient to cross common wheat with triticale than with rye [16].To develop elite germplasm resources, in this study, we aim to adequately use the breeding potential of octoploid triticale: 1) to develop wheat-rye derivatives with powdery mildew resistance and high-yielding potential using triticale and common wheat; 2) to analyze the chromosome composition of the resulting derivatives using comprehensive molecular cytogenetic methods; 3) to systematically evaluate powdery mildew resistance and agronomic traits of those elite derivative lines.
2.Materials and methods
2.1.Plant materials
09R2-95 is an octoploid triticale line(2n=8x=56,AABBDDRR)which was previously cytogenetically identified and maintained in our laboratory.09R2-95 exhibits multiple elite traits for wheat improvement, particularly yielding ability due to its large spikes,more spikelets,and a high grain yield,and has shown outstanding breeding combining ability in hybridization with common wheat cultivars during long terms of our transfer practice.For many years,09R2-95 has displayed a high level of resistance to powdery mildew in the field.Henong 827 and Heng 4338 are two susceptible wheat cultivars to powdery mildew at all growth stages.They both carried T1RS∙1BL translocation lines.09R2-95 was successively crossed with Henong 827 and Heng 4338, and then selfpollinated for nine generations.Based on field performance observations,three stable derivative lines YT4-1,YT4-2,and YT4-3 were obtained from out of tens of thousands of plants.Wheat cultivar Holdfast, rye cultivar KingII, a complete set of disomic addition lines (DA1R to 7R) of Holdfast × KingII, 6RS and 6RL ditelosomic addition lines(DTA6RS and DTA6RL),and 1RS and 1RL ditelosomic addition lines (DTA1RS and DTA1RL) of Holdfast × KingII, were used as controls in the molecular marker analysis of the wheatrye derivatives.The susceptible wheat cultivar Shixin 733 was crossed with YT4-2 to generate F2populations to determine the inheritance of powdery mildew resistance.
2.2.Molecular cytogenetic analyses
Genomic in situ hybridization (GISH), multi-color fluorescence in situ hybridization (mc-FISH), and multicolor-GISH (mc-GISH)analyses were conducted to determine the chromosomal composition of the wheat-rye derivative lines.Root tip cells were prepared for mitotic chromosome observation and hybridization [17].For GISH analysis, the genomic DNA of rye cultivar German White labeled with fluorescein-12-dUTP (green) was used as a probe,while the genomic DNA of wheat cultivar Chinese Spring was used as a blocker using a ratio of 1:50 of the probe and blocker.After GISH analysis, two highly repeated sequence probes pAs1 (red)labeled with Texas-red-5-dCTP and pSc119.2 (green) labeled with fluorescein-12-dUTP were used in combination for mc-FISH to determine the identity of specific chromosomes [18–20].Mc-GISH analysis also was conducted to determine the genomic composition of the three wheat-rye lines.Genomic DNA samples of German White and T.urartu were labeled with fluorescein-12-dUTP, that of Aegilops tauschii was labeled with Texas-red-5-dCTP, and that of Ae.Speltoides was used as a blocker [21].
2.3.Molecular marker analysis
To confirm the chromosomal composition of the wheat-rye lines YT4-1, YT4-2, and YT4-3, the 6RS-specific marker SW22057 and 6RL-specific marker SW22063 [22] were used to detect chromosomes 6RS and 6RL, respectively.The 1RS-specific marker SW5284 [22] and 1BS-specific O11B3/O11B5 [14] were used to determine the chromosome composition of T1RS∙1BL translocation line.
2.4.Phenotypic assessment of powdery mildew resistance
To evaluate the powdery mildew resistance,YT4-1,YT4-2,YT4-3, and their parents 09R2-95, Henong 827, and Heng 4338 were tested for seedling resistance by separate inoculation with a total of 21 single-pustule-derived Bgt isolates under greenhouse conditions as described in Qie et al.[23].At the one-leaf stage, all the seedlings were inoculated.After 10–14 d, when the colonies were fully developed on the first leaves of the susceptible control Mingxian 169,infection types(ITs)on each plant were assessed based on a 0–4 scale [24,25].
At the adult-plant stage,YT4-1,YT4-2,and YT4-3 and their parents 09R2-95,Henong 827,and Heng 4338 were assessed for their reaction patterns against a mixture of Bgt isolates collected at Luancheng Agro-Ecological Experimental Station, Chinese Academy of Sciences, Shijiazhuang, China (37°53′15′′N, 114°40′47′′E)from 2018 to 2021.Three replicates were tested in each year with Mingxian 169 planted beside and around as the susceptible control and inoculum spreader.For each replicate, each genotype was planted in four rows with 20 seeds per row.When Mingxian 169 showed severe disease symptoms, the powdery reactions were assessed on a 0–9 scale, in which 0–4 were considered resistant and 5–9 were susceptible [26].
2.5.Evaluation of agronomic traits
To evaluate agronomic performance, YT4-1, YT4-2, and YT4-3 and their parents 09R2-95, Henong 827, and Heng 4338 were planted in the field from 2018 to 2020 with three replicates for each genotype in a randomized complete block design.For each replicate, 20 seeds of each genotype were planted in four rows(1.5 m in length and inter-row distance of 25 cm).At physiological maturity, 20 randomly selected plants in the middle of the two internal rows were harvested, and their spike number per plant(SNPP), plant height (PH), spike length (SL), total spikelet number per spike(TSS),fertile spikelet number per spike(FSS),kernel number per spike (KNS), thousand-kernel weight (TKW), and grain yield per plant (GYPP) were determined.Analysis of variance(ANOVA) and least significant difference (LSD) tests were performed using software SPSS 22.0 (SPSS Inc., Chicago, IL, USA) to compare the yield traits of the genotypes.
3.Results
3.1.Cytogenetic analyses
GISH results demonstrated that YT4-1, YT4-2, and YT4-3 had 44, 44, and 42 chromosomes in each a cell, respectively (Fig.1A–C).All three lines possessed a pair of intact rye chromosome arms that had translocated onto a pair of wheat chromosomes.The two rye chromosome arms contained obvious secondary constriction.In addition, YT4-1 had a pair of intact rye chromosomes, whereas YT4-2 had a pair of rye ditelosomic chromosomes.
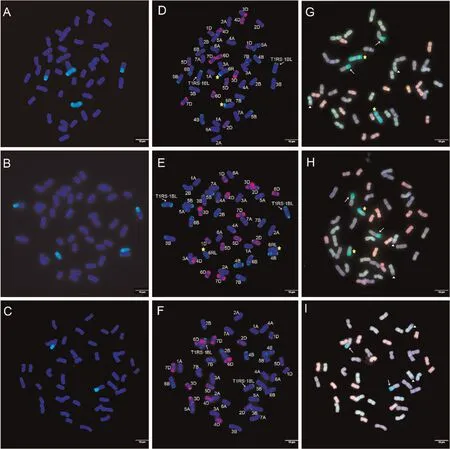
Fig.1.Genomic in situ hybridization(GISH),multicolor fluorescence in situ hybridization(mc-FISH),and multicolor-GISH(mc-GISH)identification of YT4-1,YT4-2,and YT4-3.GISH analyses of YT4-1(A),YT4-2(B)and YT4-3(C)used total genomic DNA of rye cultivar German White(green)as a probe.Sequential mc-FISH analyses(D),(E),and(F)were performed on the same metaphase after GISH analyses(A),(B),and(C),respectively,using probes pSc119.2(green)and pAs1(red).Mc-GISH analyses of YT4-1(G),YT4-2 (H), and YT4-3 (I), and R-genome, A-genome, B-genome, and D-genome chromosomes displayed bright-green, yellow-green, brown or gray, and pink or red fluorescence signals,respectively.The yellow asterisks indicate the rye chromosomes 6R(D,G)or 6RL(E,H),the white arrows indicate the translocation chromosomes T1RS∙1BL(D–I),the white triangles indicate the chromosome arrangement event in which two small B-genome chromosome fragments translocated to the two long arms of A-genome chromosomes (G–I), and the bar represents 10 μm.
After GISH analysis, mc-FISH using a combination of probes pAs1 and pSc119.2 determined the identity of the individual chromosomes.In YT4-1,pSc119.2 produced strong green hybridization signals in the telomeric region of the short arm and several weak green signals in the long arm of the intact rye chromosomes.Meanwhile, strong hybridization signals of pSc119.2 were observed in the telomeric regions and weak hybridization signals in the subtelomeric regions on both the long and short arms of the translocated rye and wheat chromosomes, respectively(Fig.1D).Thus, the additional rye chromosomes were identified as 6R and the rye chromosome arm 1RS was translocated onto the wheat chromosome arm 1BL forming a wheat-rye T1RS∙1BL translocation.YT4-2 carried the T1RS∙1BL translocation, it had two rye ditelosomic chromosomes that showed similar FISH patterns as the long arm of the rye chromosomes in YT4-1,thus were identified as 6RL ditelosomic chromosomes (Fig.1E).In YT4-3, no other rye chromatin was observed except for the wheat-rye T1RS∙1BL translocation (Fig.1F).
To further determine the genomic characteristics of the three wheat-rye lines, mc-GISH analysis was performed.Crosshybridization made the rye chromosomes display a green hybridization signal, whereas the wheat A-, B-, and D-genome chromosomes displayed yellow-green, brown or gray, and pink or red fluorescent signals, respectively.The mc-GISH results demonstrated that the three lines all had seven pairs of chromosomes from the A- and D-genome, six pairs of B genome chromosomes, and a pair of rye chromosome arms translocated onto a pair of B-genome chromosomes of wheat(Fig.1G,H, I).Compared with YT4-3 (Fig.1I white arrows), YT4-1 had a pair of additional rye chromosomes (Fig.1G yellow asterisks) and YT4-2 had a pair of additional rye ditelosomic chromosomes (Fig.1H yellow asterisks).Meanwhile, a chromosome arrangement event in which two small B-genome chromosome fragments were translocated to the two long arms of A-genome chromosomes was also observed in each of the three lines (Fig.1G, H, I white triangles).
3.2.Molecular marker analysis
Using the 6RS-specific marker SW22057, targeted fragments of approximately 400 bp in size were detected in 09R2-95, YT4-1,KingII, DA6R, and DTA6RS, but not in the genotypes Henong 827,Heng 4338, YT4-2, YT4-3, Holdfast, DA1R, DA2R, DA3R, DA4R,DA5R,DA7R,and DTA6RL(Fig.S1A).Using the 6RL-specific marker SW22063, the targeted fragments of approximately 400 bp in size were detected in 09R2-95,YT4-1,YT4-2,KingII,DA6R,and DTA6RL,but not in the other genotypes (Fig.S1B).Using the 1RS-specific marker SW5284 with approximately 400 bp diagnostic fragments,the targeted fragments were detected in 09R2-95, Henong 827,Heng 4338, YT4-1, YT4-2, YT4-3, KingII, DA1R and DTA1RS(Fig.S1C).Meanwhile, the targeted approximately 630 bp fragments genotyped by the 1BS-diagnostic marker O11B3/O11B5 were not detected in Henong 827, Heng 4338, YT4-1, YT4-2, YT4-3, and KingII, which indicated the absence of chromosome arm 1BS in those genotypes (Fig.S1D).
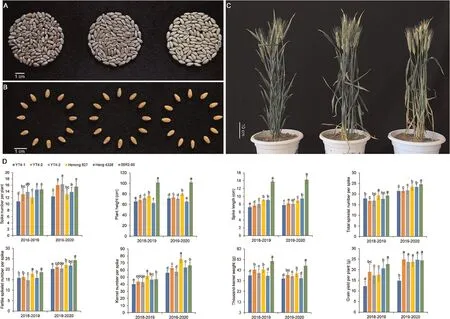
Fig.2.Agronomic performance of YT4-1, YT4-2 and YT4-3.Seed characteristics (A and B) and plants (C) of wheat-rye lines YT4-1, YT4-2, and YT4-3 (from left to right).Agronomic evaluation(D)of YT4-1,YT4-2 and YT4-3 and their wheat parents Henong 827 and Heng 4338 and octoploid triticale parent 09R2-95 from 2018 to 2020.Values followed by the same letters in the same year were not significantly different at the 0.05 probability level according to the LSD test.Error bars represent the standard deviation of the mean.
Taking the results from the molecular markers, GISH, and FISH analyses together, YT4-1, YT4-2, and YT4-3 were confirmed to be wheat-rye 6R disomic addition and T1RS∙1BL translocation line,6RL ditelosomic addition and T1RS∙1BL translocation line, and T1RS∙1BL translocation line, respectively.
3.3.Assessment and analyses of powdery mildew resistance
At the seedling stage,09R2-95 was immune to all the 21 Bgt isolates tested with IT 0, whereas Henong 827, Heng 4338, YT4-1,YT4-2, and YT4-3 were highly susceptible to all of them with IT 4, suggesting that all of the alien and wheat chromosomes in YT4-1, YT4-2, and YT4-3 had no seedling resistance gene(s) to powdery mildew.
At the adult-plant stage, 09R2-95, YT4-1, and YT4-2 were all highly resistant to the Bgt mixture with IT 0–1 for three consecutive years, and Henong 827,Heng 4338,and YT4-3 were all highly susceptible with IT 7–8 (Fig.S2A).To assess the inheritance of adult-plant powdery mildew resistance of YT4-2, it was crossed with the powdery mildew susceptible cultivar Shixin 733 to obtain an F2population.The 6RL-specific marker SW22063 was used to detect the presence or absence of 6RL in each F2plant.After the evaluation at the adult-plant stage, the population segregated in 78 resistant and 64 susceptible plants.Using the 6RL-specific marker SW22063, we found that all the resistant plants had chromosome arm 6RL, whereas all the susceptible plants did not(Fig.S2B).This result suggested that the chromosome 6RL in YT4-2 possessed APR loci.
3.4.Assessment of agronomic performance
After bagging for self-pollinating for more than nine consecutive generations, YT4-1, YT4-2, and YT4-3 were stable in either their morphological or genetic performance.YT4-1 and YT4-2 had red seeds while YT4-3 had white seeds, and their seed sizes were comparable(Fig.2A,B).All of the three wheat-rye derivative lines exhibited satisfactory agronomic performance (Fig.2C, D).Even compared with the wheat parents Henong 827 or Heng 4338, the three lines, especially YT4-2, had significantly higher or comparable SNPP, FSS, KNS, TKW and GYPP, which were crucial yield components (Fig.2D).YT4-3 is susceptible to powdery mildew at the whole growth period,and hence may have less breeding value compared with YT4-1 and YT4-2.
4.Discussion
In recent years, wheat breeding has had to deal with a genetic bottleneck that has resulted from domestication and breeding.In this regard, distant hybridization between wheat and wild relatives is an effective strategy for addressing this issue.Rye, a naturally cross-pollinating relative of common wheat, has been an important gene donor for wheat improvement [27–29].However,transferred rye chromosomes are often present in the form of addition, substitution, and translocation lines, which usually contain undesirable chromatin that can affect the yield and quality of wheat [27].Therefore, it is of prime importance to develop and/or identify alien germplasm resources with enhanced disease resistance and with good agronomic performance.
Several wheat-rye derivatives that carrying chromosome 6R have been shown to confer disease resistance in wheat, however,most of them showed poor agronomic performance,indicating that rye chromosome 6R most likely has undesirable linkage drags[16,30,31].To widen the utilization of chromosome 6R, in this study, we developed two novel wheat-rye 6R derivative lines YT4-1 and YT4-2 through an extensive screening of the progenies derived from the crossing of a triticale line with common wheat cultivars.Compared with most of the reported rye-derived 6R stocks with ASR, both YT4-1 and YT4-2 exhibited APR to powdery mildew.Generally, APR is regarded to provide more durable disease resistance in production [5,32,33].In addition, disease resistance is often at the expense of some agronomic traits and reduced plant adaptation [34].Thus, APR is more economical and favored by breeders[35].As expected,YT4-1 and YT4-2 have combined improved powdery mildew resistance with agronomic traits.In particular, YT4-2 demonstrated comparable or even superior comprehensive agronomic performance than the commercial cultivars Henong 827 and Heng 4338 that carry a 1RS translocation.It appears that the chromosome arm 6RL from octoploid triticale 09R2-95 provides a high level of powdery mildew resistance and without any negative effects on the yield traits.Although YT4-2 is a ditelosomic addition line, it has breeding potential, so it is expected to be a promising bridge resource for wheat breeding.
To accelerate the use of the disease resistance in breeding practice, we have transferred the chromosome arm 6RL of YT4-2 into several wheat cultivars to produce a number of diverse progenies.As expected,the chromosome arm 6RL in YT4-2 has demonstrated wide adaptability in all the wheat backgrounds into which it was transferred, which may have the potential to produce elite and stable small-segment translocation lines.In the future, more research will be conducted on chromosome arm 6RL including its genetic effects on agronomic and quality traits,transmission characteristics in different wheat backgrounds, and the evaluation of agronomic performance of the stable lines in different ecological regions.
CRediT authorship contribution statement
Guohao Han:Writing–original draft,Resources,Formal analysis,Methodology.Jing Wang:Methodology,Formal analysis,Visualization.Hanwen Yan:Methodology, Formal analysis,Investigation.Tiantian Gu:Methodology, Formal analysis, Investigation.Lijun Cao:Methodology, Formal analysis, Visualization.Shiyu Liu:Methodology, Formal analysis.Xiuquan Li:Resources,Formal analysis.Yilin Zhou:Methodology, Formal analysis.Jieru Fan:Formal analysis, Investigation.Zhipeng Shi:Formal analysis.Hong Liu:Formal analysis.Lihui Li:Conceptualization, Supervision.Diaoguo An:Conceptualization, Review & Editing, Supervision, Funding acquisition.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This research was supported by the National Key Research and Development Program of China (2021YFD1200600) and the National Natural Science Foundation of China (32272105).The authors are grateful to Dr.Robert L.Conner for critically reviewing and professionally editing this manuscript.The authors are grateful to the following researchers for providing valuable plant materials:Dr.Zongxiang Tang (Sichuan Agricultural University, Chengdu,Sichuan, China) for wheat cultivar Holdfast and a set of wheatrye disomic addition lines of Holdfast×King II;Dr.Yiwen Li(Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing, China) for rye cultivar King II; Dr.Adrian Turner(John Innes Centre, Norwich, UK) for wheat-rye 1RS, 1RL, 6RS and 6RL ditelosomic addition lines of Holdfast × King II.
Appendix A.Supplementary data
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2023.09.003.

- The Crop Journal的其它文章
- Flowering-time regulation by the circadian clock: From Arabidopsis to crops
- Global characterization of OsPIP aquaporins reveals that the H2O2 transporter OsPIP2;6 increases resistance to rice blast
- Drought-triggered repression of miR166 promotes drought tolerance in soybean
- The OsBSK1-2-MAPK module regulates blast resistance in rice
- Natural variation of an autophagy-family gene among rice subspecies affects grain size and weight
- Rice gene OsUGT75A regulates seedling emergence under deep-sowing conditions