
ZmCYP90D1 regulates maize internode development by modulating brassinosteroid-mediated cell division and growth
2024-03-07 01:51CnrnSunYngLiuGuofngLiYnleChenMengyunLiRuihuYngYongtinQinYongqingChenJinpengChengJihuTngZhiyunFu
The Crop Journal 2024年1期
Cnrn Sun, Yng Liu, Guofng Li, Ynle Chen, Mengyun Li, Ruihu Yng, Yongtin Qin,Yongqing Chen, Jinpeng Cheng, Jihu Tng,,*, Zhiyun Fu,*
a Key Laboratory of Wheat and Maize Crops Science/Collaborative Innovation Center of Henan Grain Crops/College of Agronomy, Henan Agricultural University, Zhengzhou 450046, Henan, China
b The Shennong Laboratory, Zhengzhou 450002, Henan, China
c Hebi Academy of Agricultural Sciences, Hebi 458030, Henan, China
d Yunnan Tang Jihua Expert Workstation/Zhaotong Academy of Agricultural Sciences, Zhaotong 657000, Yunnan, China
Keywords:Maize ZmCYP90D1 BR biosynthesis Dwarf plant
ABSTRACT Plant height(PH)is associated with lodging resistance and planting density,which is regulated by a complicated gene network.In this study,we identified a spontaneous dwarfing mutation in maize,m30,with decreased internode number and length but increased internode diameter.A candidate gene,ZmCYP90D1,which encodes a member of the cytochrome P450 family, was isolated by map-based cloning.ZmCYP90D1 was constitutively expressed and showed highest expression in basal internodes,and its protein was targeted to the nucleus.A G-to-A substitution was identified to be the causal mutation, which resulted in a truncated protein in m30.Loss of function of ZmCYP90D1 changed expression of hormoneresponsive genes, in particular brassinosteroid (BR)-responsive genes which is mainly involved in cell cycle regulation and cell wall extension and modification in plants.The concentration of typhasterol(TY),a downstream intermediate of ZmCYP90D1 in the BR pathway,was reduced.A haplotype conferring dwarfing without reducing yield was identified.ZmCYP90D1 was inferred to influence plant height and stalk diameter via hormone-mediated cell division and cell growth via the BR pathway.
1.Introduction
PH is an agronomic trait strongly associated with lodging resistance and planting density, which in turn increasing crop yield[1,2].The Green Revolution genes, such as Ossd1 and TaRht,involved in gibberellin (GA) biosynthesis or signaling, have been widely used in rice and wheat breeding by adopting semi-dwarf varieties that were resistant to lodging[3,4].But their maize orthologs usually confer unfavorable pleiotropic phenotypes, including reduced crop yields [5–8].
The MaizeGDB database(https://www.maizegdb.org) describes more than 50 PH-associated loci, most of them involved in plant hormone biosynthesis, transport, and signaling.Mutations in GA-biosynthesis genes result in short internodes and dwarf plants.Examples are DWARF1,encoding a GA 3-oxidase that catalyzes the final step in GA biosynthesis, DWARF3, encoding an ent-kaurenoic acid oxidase that catalyzes three steps in GA biosynthesis, and DWARF5, encoding an ent-kaurenoic acid oxidase that catalyzes ent-kaurenoic acid to GA12 [7–10].Mutations in DWARF8 and DWARF9, two negative regulators of GA signaling, also produce dwarf plants[6].BR is another important plant hormone that influences PH [11,12].Several cytochrome P450-catalyzed oxidative reactions play an important role in BR biosynthesis and their disruption reduced PH.Many mutants of CYP724B1, which involve in the generation of 6-deoxoTY and TY, such as dwarf11, clustered primary branch1, grain number and size on chromosome4 in rice,and zmd11 in maize, exhibit reduced PH [13–16].Mutation of CYP90B1, encoding a 22 alpha-hydroxylase which converts campestanol to 6-deoxocathasterone in Arabidopsis, inhibits cell elongation and produces a dwarf phenotype [17].Mutants of
Arabidopsis CYP90A1, CYP90C1 and CYP90D1 all exhibited reduced stature,although they are involved in different steps of BR biosynthesis [18–21].In maize and rice, mutations of CYP90D1 ortholog,including d1,EBISU DWARF(d2),CHROMOSOME SEGMENT DELETED DWARF 1 (csdd1), SMALL GRAIN 11 (smg11), and qLTG1, have also been characterized with shorter PH [22–26].In addition, genes involved in BR signal transduction cascade also influence plant growth and development, such as BR-INSENSITIVE 1-ASSOCIATED RECEPTOR KINASE 1 (BAK1).Loss of function of BAK1 in rice and Arabidopsis both reduced sensitivity to BR and PH [27,28].Thus,BR biosynthesis and signaling are both required for plant development.
Here,we identified a dwarf mutant,m30,with shorter and fewer internodes in a maize breeding population.To isolate the candidate gene and elucidate its molecular mechanism on PH, we characterized the phenotype and cytological features of m30, positionally cloned the causal gene ZmCYP90D1 of m30, and explored the expression level changes influenced by loss function of ZmCYP90D1.We further performed candidate-based association analysis for ZmCYP90D1 to identify favorable haplotype in natural population for its exploit in practices.Our results show that ZmCYP90D1 influences PH through mediating BR-related genes which involved in cell division,cell wall biosynthesis,and vascular bundle formation.Importantly, there is a favorable haplotype in ZmCYP90D1 for a shorter PH without reducing yield.
2.Materials and methods
2.1.Plant materials
The spontaneous PH mutant m30 from a maize breeding population was crossed with the inbred line YU87-1 to generate an F1,which was backcrossed to m30 to produce a BC1mapping population.All plant materials were planted at Henan Agricultural University experimental sites in Henan and Hainan, China.The PH phenotypes of BC1plants were recorded from 10 to 60 days after sowing.The number, length, and diameter of internodes and ear leaves, as well as yield traits, were recorded at maturity.For RNA-sequencing (RNA-seq), the first to third internodes of the wild type (WT, YU87-1) and m30 mutant were sampled at the R1 stage with two replications.
2.2.Cytological analysis
The third internodes above the ground of five plants were sampled and sectioned using a vibratome (VT1000 S, Leica, Germany)[29].Internode tissues were fixed in formalin:acetic acid:alcohol fixative and evacuated three times for 5 min each time, followed by washing with 1×phosphate buffered saline for 10 min at room temperature.Tissues were then embedded in 3% agarose and processed into 30 μm-thick sections, which were stained with a Saffron-O and Fast Green Stain kit (for plants) (G1375, Solarbio,Beijing, China) and observed under an emicroscope (Axio Scope A1, Zeiss, Germany).
2.3.Map-based cloning of m30 locus
The m30 locus was assayed using 276 InDel markers distributed evenly across the 10 maize chromosomes by bulk-segregant analysis.New InDel or single-nucleotide polymorphism(SNP) markers were developed,and a BC1mapping population of 1152 plants was used to narrow the map interval containing the m30 locus.All primers used are listed in Table S3.Candidate genes were annotated according to the B73 reference genome (RefGen_V5).
2.4.Subcellular localization of ZmCYP90D1
The coding sequence of ZmCYP90D1 was fused with the enhanced GFP (eGFP) reporter gene and cloned into the pRTL vector(35SPro::ZmCYP90D1-eGFP).The resulting construct and the control nuclear marker Ghd7-RFP were then co-transformed into maize protoplasts.The transfected protoplasts were cultured overnight in the dark at 28 °C and observed under a confocal microscope(LSM710, Zeiss, Germany).All primers used are listed in Table S3.
2.5.RNA-seq and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted from internodes, roots, ears, tassels,leaves, and husks of the WT and m30 at the R1 stage using the TransZol Plant Kit (ET121, Transgen, Beijing, China).
For RNA-seq,an RNA library for RNA-seq was prepared,and the concentration of the library was measured with a Qubit fluorometer (ThermoFisher).Samples with two biological replicates were subjected to Illumina sequencing (Illumina NovaSeq 6000), and clean reads were mapped to the B73 reference genome(RefGen_V5).For each transcript model, gene expression levels were converted to fragments per kilobase million.Differentially expressed genes(DEGs)were defined by the following criteria:fold change>2 and adjusted P value<0.05.Gene Ontology(GO)enrichment analysis of the DEGs was performed with agriGO(https://systemsbiology.cau.edu.cn/agriGOv2/) [30].Functional annotation of the DEGs was performed using BLAST analyses against the UniProt database (https://www.uniprot.org/blast/) and the NCBI database(https://www.ncbi.nlm.nih.gov/datasets/taxonomy/4577/).Biological processes of the DEGs were identified by the Kyoto Encyclopedia of Genes and Genomes (KEGG) (KOBAS, https://kobas.cbi.pku.edu.cn/) [31].
For reverse transcription, cDNA was synthesized from 1 μg of total RNA using an All-in-One First-Strand Synthesis Master Mix(with dsDNase) (KR0501, Kemix, Zhengzhou, Henan, China), and quantitative polymerase chain reaction (qPCR) was performed on a Bio-Rad CFX96 system using a 2× SYBR Green qPCR Premix(Universal) (KS0601, Kemix, Zhengzhou, Henan, China).Relative expression levels were calculated using the 2-ΔΔCT(threshold cycle)method with actin as the internal standard gene[32].All primers used are listed in Table S3.
2.6.Liquid chromatography tandem mass spectrometry (LC-MS/MS)analysis of endogenous brassinosteroid
Metabolites of BR biosynthesis were measured in internodes of the ear,above the ear,and below the ear of m30 and WT at the R1 stage using MetWare(https://www.metware.cn/)with an AB Sciex QTRAP 6500 liquid chromatography tandem mass spectrometry(LC-MS/MS) instrument.Each sample had three biological replicates, and each replicate represented more than three plants.Student’s t-test was used to compare metabolite abundances between WT and m30.
2.7.Candidate gene-based association analysis and haplotype identification
An association mapping population containing 509 maize inbred lines from temperate, tropical/subtropical, and mixed groups(https://www.maizego.org/Resources.html/) was used for candidate gene-based association analysis.The population was planted in three environments (Yongcheng in Henan province, Yuanyang in Henan province,and Xingtai in Heibei province in 2022, China),and kernel weight per ear was measured using five ears for each genotype in each environment.Data for other agronomic traits,and a set of 299 SNPs surrounding or within ZmCYP90D1 were retrieved from MaizeGo (https://www.maizego.org/Resources.html/).SNPs were filtered to ensure a minor allele frequency>0.05 and a missing rate<20%.Analysis was performed with TASSEL v.5.0(https://www.maizegenetics.net/copy-of-tassel/) with a mixed linear model considering kinship and population structure [33].Linkage disequilibrium and Manhattan plots were generated with LDBlockShow (https://github.com/BGI-shenzhen/LDBlockShow/).The expression data of ZmCYP90D1 in 368 inbred lines were retrieved from the reported transcriptome[34].Phenotypic values,expression levels, and haplotypes were compared by one-way ANOVA or Student’s t-test.
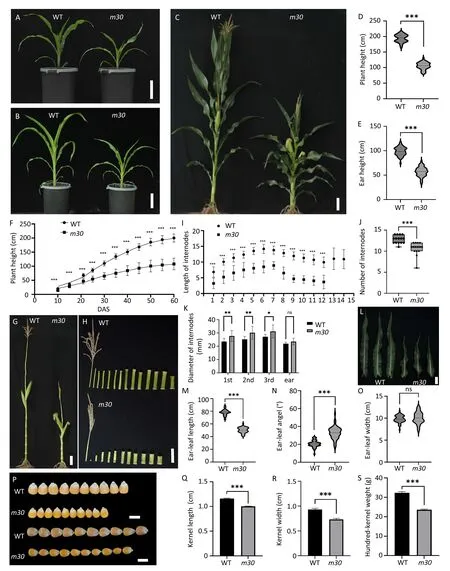
Fig.1.Plantand kernel characterizationofm30.(A–C)Plantphenotypicfeatures of wildtype(WT)andm30attheV4(A),V6(B),andR1(C)stages.Scalebars,10cm.(D,E)Comparisonof PH andEHbetweenWTandm30.n=50(***,P<0.001,t-test).(F)VariationofPHbetweenWTandm30onmultipledaysaftersowing(DAS).Valuesaremeans±standarddeviation(SD);n=25(***,P<0.001,t-test).(G)Plant phenotype without leaves between WT and m30 at maturity.Scale bar,10 cm.(H)Internode phenotype at maturity between WT and m30.Scale bar,10 cm.(I)ComparisonofinternodenumberbetweenWTandm30atmaturity.Valuesaremeans±SD;n=15(***,P<0.001,t-test).(J)ComparisonofinternodelengthbetweenWTandm30atmaturity.Data are shown in scatter plot;n=15(***,P<0.001,t-test).(K)Comparison of internode diameter between WTand m30 at maturity.Values are means±SD;n=12(*,P<0.05;**,P<0.01;ns,not significant,t-test).(L)Ear-leaf phenotype at maturity between WT and m30.Scale bar,10 cm.(M-O)Comparison of length(M),angle(N),and width(O)of the ear-leaf between WT and m30.n=50(***,P<0.001;ns,notsignificant,t-test).(P)MatureWTandm30kernels.Scalebar,1cm.(Q–S)Comparisonofkernellength(Q),width(R),andhundred-kernelweight(S)between WT and m30.Values are means±SD;n=30 for length and width;n=3 for hundred-kernel weight(***,P<0.001,t-test).
3.Results
3.1.Phenotypic characterization of m30
Compared with the WT, the m30 mutant showed increasingly delayed growth following plant development (Fig.1A, B) and displayed shorter PH and ear height(EH)with 57.6%and 57.7%reductions at the pollen shedding stage, respectively (Fig.1C–E).PH measurement for whole developmental stages indicated that the difference in PH and EH between m30 and WT lasted from 10 days after seedling (DAS) to 60 DAS (Fig.1F).At the mature stage, both the number and the length of internodes, especially the fourth to the eighth, decreased in m30 compared with the WT (Fig.1G–J).In contrast,the width of internodes,especially the first to the third,was significantly increased in m30 (Fig.1K).Additionally, the earleaf showed a significant reduction in length (by 35.7%) (Fig.1L,M) and an increase in leaf angle (by 59.9%) (Fig.1N) but not in width (Fig.1O) in m30 compared with the WT.The reduction in PH and EH consequently resulted in a decrease in kernel length(18.8%), kernel width (21.5%), and hundred-kernel weight (26.8%)(Fig.1P–S).The results indicate that a shorter PH of m30 is derived from a reduction in both the number and length of the internode,which in turn affects kernel yield.
3.2.Cell division and growth are altered in m30
Transverse sections showed light safranin staining in the stem epidermis and vasclar bundles in m30 compared to WT (Fig.2A,B), indicating reduced lignin in m30.m30 showed an increase in stalk cell area and a decrease in cell number per mm2(Fig.2C,D).By contrast, the length of epidermal and parenchyma cells of the internode was significantly decreased (Fig.2E, F), especially in some extreme m30 individuals with twisted vascular bundles(Fig.2G), but no significant changes in width were observed between WT and m30 in longitudinal sections (Fig.2H).These results indicate that cell division and growth are affected in m30.
3.3.ZmCYP90D1 is the causal gene of m30
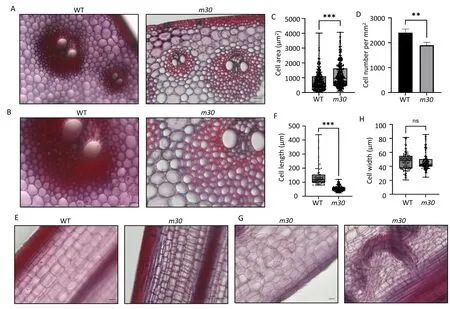
Fig.2.Cytological differences in stem cells between WT and m30.(A–D)Cross sections(A,B),cell area beyond the epidermis and vascular bundles(C),and cell number per mm2(D)of the third internodes at the R1 stage between WT and m30.Scale bars,50 μm for(A)and 20 μm for(B).Data are shown in scatter plot(C).Values are means±SD(D);for cell area,n=385 for WT and n=389 for m30;for cell number per mm2,n=3 for WT and n=4 for m30(**,P<0.01;***,P<0.001,t-test).(E–H)Longitudinal sections(E,G),cell width(F),and cell length(H)of the third internodes at the R1 stage between WT and m30.Scale bars,50 μm for(E)and 20 μm for(G).Data are shown in scatter plot(F and H); for width, n = 113 for WT and n = 142 for m30; for length, n = 84 for WT and n = 163 for m30 (***, P < 0.001; ns, not significant, t-test).
The PH of F1was similar to that of YU87-1,and the BC1population was segregated for PH with a ratio of 1:1 between WT(m30/+)and dwarf plants (m30/m30) (Table S1), indicating that the m30 phenotype was controlled by a single recessive nuclear gene.The candidate gene was mapped on chromosome 3 between two markers,S05065 and S05297.The region was further narrowed to a 271-kb interval between newly developed InDel markers S05141 and S05150 using a population of 518 WT (m30/+) and 634 dwarf mutants (m30/m30) (Fig.3A).Nine genes were in this interval(Fig.3A), and cDNA sequencing of these genes revealed a G-to-A mutation in the eighth exon of Zm00001eb120890, which caused a premature termination of the predicted protein translation(Fig.3B,C).The dCAPS marker developed based on the G-to-A conversion was found to co-segregate with the PH phenotype.As shown in Fig.3D,all dwarf plants with homozygous m30 genotype were represented by a large uncleaved fragment, whereas all normal individuals with homozygous wild-type or heterozygous genotype showed a small fragment or two fragments.Thus, the G-to-A conversion co-segregated with the dwarf mutation.(Fig.3D).
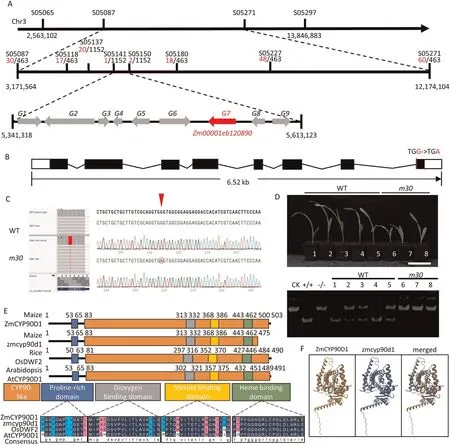
Fig.3.Positional cloning of the m30 locus and co-segregation test.(A) The m30 locus was mapped to a 271-kb interval on chromosome 3 according to the B73 reference genome.Red and black numbers represent recombinants and population size, respectively.Zm00001eb120810 (G1), Zm00001eb120820 (G2), Zm00001eb120840 (G3),Zm00001eb120850(G4),Zm00001eb120860(G5),Zm00001eb120880(G6),Zm00001eb120890(G7),Zm00001eb120900(G8),and Zm00001eb120910(G9).(B)Schematic structure and polymorphic sites of Zm00001eb120890 between WT and m30.Lines represent introns,black boxes represent exons,and white boxes represent the 5ʹand 3ʹuntranslated regions.(C) Sequence polymorphisms of Zm00001ed0120890 between WT and m30 RNA-seq cDNA sequence (left) and genome (right).Red arrows indicate the causal mutation.(D)Co-segregation test of m30,showing seedlings with WT and m30 phenotypes(top).Scale bars,1 cm.The genotype of seedlings was identified using the dCAPs markers 1st-2nd-L/2nd-RP-cut-with-KpnI-R,which were designed based on the mutation site of m30.CK, PCR product without enzyme digestion by KpnI;+/+,homozygous M30, -/-, homozygous m30.(E) Alignment of amino acid residues among ZmCYP90D1, zmcyp90d1 in m30, OsDWF2, and AtCYP90D1.Zm00001eb120890 was predicted to contain one small domain (blue square, proline-rich domain) and one large domain (orange square, CYP90-like domain, containing three small domains: gray square,dioxygen binding domain;yellow square,steroid binding domain;green square,heme binding domain).Conserved amino acids are highlighted in blue.(F)Three-dimensional structure of Zm00001ed0120890 protein between WT(left)and m30(middle).WT is represented in khaki,m30 is represented in blue,and the missing 28 aa residues in m30 are represented in gold.
The full-length cDNA of Zm00001eb120890 comprises an open reading frame of 1512 bp, a 5′-untranslated region (UTR) of 248 bp, and a 3′-UTR of 490 bp (Fig.3B).Zm00001eb120890 is the orthologue of Arabidopsis CYP90D1 and rice OsD2,which share several conserved domains,including a proline-rich domain,dioxygen binding domain, steroid binding domain, heme binding domain,and CYP90-like active site (Fig.3E).The truncated protein in m30 lost 28 amino acids in the C-terminus, resulting in a shortened CYP90-like active site and a conformational change in the CYP90D1 three-dimensional structure (Fig.3E, F).Thus, the disruption of Zm00001eb120890 was the cause of the reduction in PH and EH of m30.This gene was named ZmCYP90D1.
3.4.ZmCYP90D1 is constitutively expressed and its protein is targeted to the nucleus
To characterize the temporal and spatial expression patterns of ZmCYP90D1, the transcript abundance of ZmCYP90D1 in a wide range of tissues and organs was measured by qRT-PCR.ZmCYP90D1 was constitutively expressed in all examined tissues, with high expression in roots and internodes, especially the first to the third internodes (Fig.4A), in agreement with online RNA-seq data(https://www.maizegdb.org).A 35SPro::ZmCYP90D1-eGFP vector was transiently expressed in maize leaf protoplasts to ascertain the subcellular location of ZmCYP90D1.As shown in Fig.4B, eGFP fluorescence signals were concentrated in the nucleus and overlapped with RFP fluorescence signals of the nuclear marker GHD7(Fig.4B).These results indicate that ZmCYP90D1 constitutively expresses and its protein localizes to the nucleus.
3.5.BR pathway was changed in m30
To investigate changes at gene expression levels in response to the ZmCYP90D1 mutation,the transcriptome profiles of the first to three internodes above ground of WT and m30 at the rapid growth stage were characterized by RNA-seq.Of 3686 differentially expressed genes (DEGs), 1631 were up-regulated and 2055 were down-regulated in m30(Fig.5A;Table S2).Of the DEGs,2977 were functionally annotated and were enriched in GO: 0,009,408 (response to heat; P = 4.8E-15),GO: 0,009,266(response to temperature stimulus; P = 4.4E-08), and GO: 0,006,355 (regulation of transcription, DNA-templated; P = 4.8E-10) (Fig.5B).Among the 290 DEGs in GO: 0006355, 29 were transcription factors (TFs)involved in plant hormone signaling as determined by KEGG, and could be grouped into BR,auxin,ethylene,and abscisic acid biosynthesis pathways(Fig.5C;Table S2).Other non-TF DEGs involved in BR, GA, and cytokinin biosynthesis pathways were also identified,most of them up-regulated in m30(Table S2).In the BR biosynthesis pathway, the four up-regulated DEGs encoded cytochrome P450 family proteins: one CYP90D1 (Zm00001eb284090); two
CYP90A1, two orthologs of Arabidopsis CPD (Zm00001eb196530,Zm00001eb093140); and one CYP734A1 (Zm00001eb122680)(Fig.5D).Many DEGs responded to BR levels.ZmBZR1 (Zm00001eb120750),the direct target of the CYP90 gene family,which is a BRresponsive TF negatively regulating cell and fruit size [35], was down-regulated in m30 (Fig.5C).The expression trend of ZmBZR1 was complementary to that of ZmCYP90D1 in the basal three internodes, suggesting that the BR pathway modulates internode development in maize.A set of 905 DEGs were classified as downstream target genes of BZR1, including 343 up-regulated and 562 down-regulated(Fig.5E;Table S2)[36].These DEGs were functionally annotated as involved in the cell cycle and cell wall extension and modification(Fig.5F,G;Table S2).Surprisingly,only TY among the 11 detected metabolites showed significant a decrease in m30(Table 1).
3.6.Favorable haplotype of ZmCYP90D1 conferring dwarf PH without reducing yield
Among the 299 SNPs surrounding or within ZmCYP90D1,15 and 13 SNPs were significantly associated with PH and EH,respectively(Fig.6A, B).Among them, eight were associated with both PH and EH.SNPs chr3.s_4747984, chr3.s_4748007, and chr3.s_4748739 were found in introns, whereas chr3.s_4752835, chr3.s_4752992,chr3.s_4753166, chr3.s_4753349, and chr3.s_4753446 were found in the promoter region(Fig.6A,B).Two of the most strongly associated SNPs, chr3.s_4752992 (8.11E-05) and chr3.s_4753166(4.23E-05), were used to define haplotypes.The 507 maize inbred lines were classified into three haplotypes based on these two SNPs(Fig.6C).Hap 1 was the largest group and was present in 335 lines,Hap 2 was present in 126 lines and Hap 3 in only 31 lines.In comparison with Hap 1 and Hap 3, inbred lines that carried Hap 2 showed reduced PH and EL (ear length) (Fig.6D, E), but without a significant difference in ear weight or kernel weight per ear(Fig.6F, G).Hap 2 should be a favorable haplotype for shorter PH without a significant reduction in yield in natural population.Because Hap 3 was carried by only a few lines and the PH of these lines was similar to that of Hap 2, they were combined into one group.Hap 1 inbred lines showed significantly lower ZmCYP90D1 transcriptions than Hap 3 and Hap 2 lines (Fig.6H).

Fig.4.Expression pattern and subcellular localization of CYP90D1.(A) Expression levels of ZmCYP90D1 in WT and m30 in the first to fifth internodes as well as roots, ears,tassels, leaves, and husks.Values are means ± standard error of mean; n = 3 (*, P < 0.05; **, P < 0.01; ***, P < 0.001; ns, not significant, t-test).(B) Subcellular location of CYP90D1-eGFP in maize protoplasts.The full-length coding sequence of ZmCYP90D1 was fused to the C-terminus of eGFP.The fusion construct and GHD7-RFP (nuclear marker) were transiently expressed in maize protoplasts.Scale bars, 10 μm.
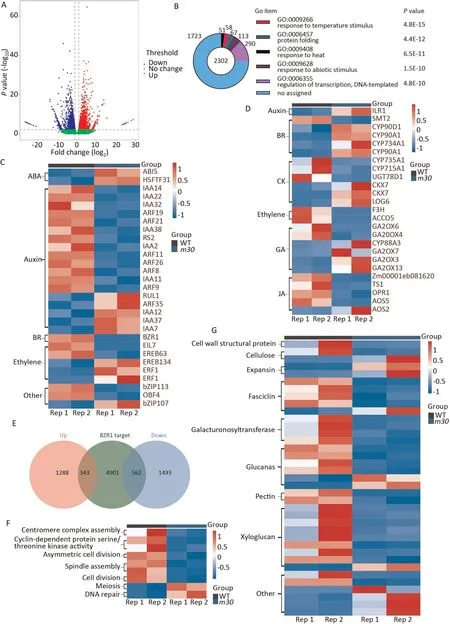
Fig.5.DEGs and expression analysis of BR response genes between WT and m30.(A)Gene expression classification of DEGs.Blue and red circles indicate respectively downand up-regulated genes(significance level:FDR value<0.05,|log2 fold change|>1).(B)Most significantly enriched Gene Ontology terms in biological process categories are shown.(C–D)Heat map of expression of transcription factors involved in plant hormone signaling(C)and plant hormone biosynthesis(D).(E)Venn diagram of ZmBZR1 target genes and PH-associated DEGs in WT and m30.(F–G) Heat map of expression of DEGs in (E) involved in the cell cycle (F) and cell wall extension and modification (G).
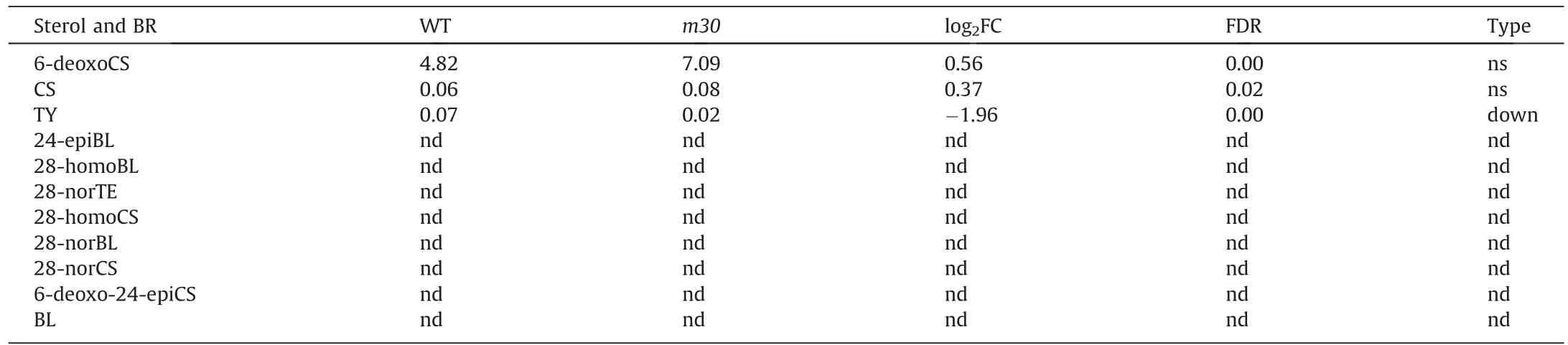
Table 1 Contents of endogenous BRs in WT and m30 measured by LC-MS/MS.

Fig.6.Natural variations and haplotypes of ZmCYP90D1.(A,B)Candidate gene-based association mapping and pairwise linkage disequilibrium analysis of ZmCYP90D1.Circles represent SNPs and triangles represent lead SNPs.The SNPs most strongly associated with PH(A)and EH(B)are denoted with squares.(C)Haplotypes(Hap) of ZmCYP90D1 associated with the mapping population.(D–H)Comparison of PH(D),EL(E),ear weight(F),and kernel weight per ear(G),as well as expression(H)of the three Haps.Values are means ± standard error of mean (SE); n = 335 for Hap 1; n = 126 for Hap 2; n = 31 for Hap 3; n = 157 for Hap 2, 3 (**, P < 0.01; ***, P < 0.001; ns, not significant, t-test).
4.Discussion
TY has been reported as the downstream intermediate of CYP90D1 in Arabidopsis [20,21].In this study, we found that the TY content was significantly reduced in m30, which showed down-regulation of many BR-response genes, indicating involvement of ZmCYP90D1 in the hydroxylation of C-23 (Fig.5C–G;Table 1).CYP90D1 is a direct effector of BZR1 in loquat [35], however, we found that BZR1 gene was significantly down-regulated but ZmCYP90D1 was significantly up-regulated in m30, a complementary expression trend between ZmCYP90D1 and BZR1.A similar complementary expression trend was also identified in previous research in maize [26].However, the reason for this phenomenon was unknown.
Both stalk length and width are regulated by stem elongation,which is driven by cell division and cell expansion of the internode[37–39].In Arabidopsis, BR has been found in several studies to be associated with cell division.Balanced BR signaling is required for a normal cell cycle in the root meristem, and mitotic activity decreased in a BR-sensitive mutant bri1-116, which showed a defective meristem [40].BR-deficient dwarf mutant cpd exhibited a prolonged cell division phase and delayed differentiation in leaves[41].BIN2 influences cell division by interacting with tubulin protein [42].BRs also function at the cell wall to influence cell expansion or elongation.In Arabidopsis,BES1 regulated expression levels of cellulose synthase genes, which in turn modulate cellulose synthesis.Cellulose was reduced in BR-deficient mutant det2-1 or BR-perceptional mutant bri1-301 compared with the wild-type, which restricted cell elongation [43].In Utricularia gibba,defects of BR biosynthesis reduced wall relaxation properties in the epidermis, resulting in twisted internal tissue and shorter and smaller epidermal cells [44].In contrast, BR stimulated hypocotyl elongation by expanding cell wall in soybean(Glycine max L.)[45].In the maize BR biosynthesis pathway, several genes have been identified as associated with PH development through the regulation of cell wall biogenesis and cell division.A mutation in ZmDWF1,which encodes 24-sterol reductase,affects the transcript levels of cell wall biogenesis genes.A mutation in Brassinosteroiddeficient dwarf1,which encodes C-6 oxidase,increased cell density per unit area, and reduced PH [46].In the BR signaling pathway,BRI1 encodes a member of leucine-rich-repeat receptor-like kinases,and RNA interference of BRI1 results in decreased cell division and cell elongation, producing shorter internodes and dwarf plants [47].In the present study, the mutation in ZmCYP90D1 caused changes in the transcript levels of cell wall biosynthesis(41) and cell division-associated (44) genes (Table S2), similar to those of the cell size-associated genes in ZmD1 mutant[26],a finding consistent with that of other BR-responsive genes, resulting in increased cell area (Fig.2C), decreased cell length (Fig.2F), and reduced cell number (Fig.2D).By contrast, very few common cell size DEGs were identified in these two studies(Table S2),possibly due to differences in sampling stages and internodes.We infer that ZmCYP90D1 likely modulates maize PH via BR-mediated cell wall biogenesis and cell division.
Short and thick stature is crucial for increasing planting density and lodging resistance in maize [48].Dwarf and semi-dwarf mutants caused by deficiency of GA biosynthesis and signaling genes in rice(e.g.,Sd1)and wheat(e.g.,Rht)have been widely used in breeding.However, these mutations in maize reduce yield[4,6,8].PH and 100-kernel weight were reduced in the zmxyl mutant [49], PH was increased and kernel weight per ear was reduced with ZmLBD5 overexpression, and the opposite changes were observed in the loss-of-function mutant[50],PH,EL,and seed weight were reduced in the terminal ear 1 mutant[51],and PH and yield were reduced in the sucrose transporter2 mutant [52].Although mutation of ZmCYP90D1 reduced PH and yield, we identified a favorable naturally occurring haplotype that reduced PH without affecting yields (Fig.6D, F, G).Thus, the ZmCYP90D1 mutant allele identified in this study has great potential to be utilized in the breeding of new dwarf varieties suitable for highdensity planting without yield loss.
5.Conclusions
The candidate gene ZmCYP90D1 of the maize dwarf mutant m30 may regulate BR biosynthesis in internodes, influencing internode development.This regulatory function may involve cell division and cell wall synthesis.Natural variation in ZmCYP90D1 was associated with PH but not yield.ZmCYP90D1 may be useful in breeding lodging-resistant maize cultivars for high-density planting.
CRediT authorship contribution statement
Canran Sun:Conceptualization, Investigation, Data curation,Methodology, Writing – original draft, Writing – review & editing.Yang Liu:Conceptualization,Investigation,Data curation.Guofang Li:Investigation, Data curation, Methodology.Yanle Chen:Conceptualization, Investigation.Mengyuan Li:Conceptualization,Investigation.Ruihua Yang:Investigation.Yongtian Qin:Data curation,Methodology.Yongqiang Chen:Investigation,Data curation.Jinpeng Cheng:Investigation,Data curation.Jihua Tang:Conceptualization, Visualization, Supervision, Funding acquisition,Writing – review & editing.Zhiyuan Fu:Conceptualization, Data curation, Writing – original draft, Writing – review & editing,Supervision, Funding acquisition, Project administration.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (U2004144, 31971893, 32101743), the Key Technologies R & D Program of Henan Province (232102111080), and Yunnan Academician Expert Workstation (202305AF150082).
Appendix A.Supplementary data
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2023.11.002.

- The Crop Journal的其它文章
- Flowering-time regulation by the circadian clock: From Arabidopsis to crops
- Global characterization of OsPIP aquaporins reveals that the H2O2 transporter OsPIP2;6 increases resistance to rice blast
- Drought-triggered repression of miR166 promotes drought tolerance in soybean
- The OsBSK1-2-MAPK module regulates blast resistance in rice
- Natural variation of an autophagy-family gene among rice subspecies affects grain size and weight
- Rice gene OsUGT75A regulates seedling emergence under deep-sowing conditions