
饲料混合植物油替代鱼油对不同品种异育银鲫生长性能和脂肪代谢的影响
2024-03-02 06:38李红燕巫丽云金俊琰朱晓鸣刘昊昆杨云霞解绶启
水生生物学报 2024年4期
李红燕 巫丽云 金俊琰 韩 冬 朱晓鸣 刘昊昆 杨云霞 解绶启
(1. 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室, 武汉 430072;2. 中国科学院大学现代农业科学学院, 北京 100049)
水产养殖为人类消费提供了近2/3的动物蛋白来源, 而水产养殖业的发展高度依赖于水产饲料业的发展。脂类是水产饲料中重要的营养素, 也是维持生物体细胞膜和亚细胞膜完整性的必需成分。脂肪作为激素的前体, 参与许多重要的生物过程,可以促进脂溶性维生素的吸收[1]。鱼油因其富含必需长链多不饱和脂肪酸(LC-PUFAs)、二十碳五烯酸(EPA)和二十二碳六烯酸(DHA), 在鱼体脂肪代谢和抗氧化等生理生化过程中发挥重要作用, 被认为是水产饲料中的优质脂肪源[2]。然而, 目前全球鱼油产量可能不足以满足未来水产养殖的需求, 导致了饲料成本的增加[3]。另外, 鱼油的生产造成野生鱼类资源和水资源环境的破坏, 阻碍了水产养殖业的快速发展。因此, 寻找适宜的饲料脂肪源来替代鱼油, 对水产养殖可持续发展具有重要意义[4]。
与鱼油相比, 植物油具备廉价易得、产量丰富及较易储存等优势, 具有替代鱼油的潜能[5]。研究表明, 大豆油、棕榈油、菜籽油和亚麻油等多种植物油, 已被用来部分或完全替代鱼油而被广泛应用于水产饲料中[6—8]。在大黄鱼(Larimichthys crocea)中, 饲料亚麻籽油和豆油完全替代鱼油, 鱼体增重率和特定生长率显著降低[6]。大豆油、菜籽油和亚麻油分别完全替代鱼油导致军曹鱼(Rachycentron canadum)生长迟缓、腹部和肝脏脂肪沉积[7,8]。植物油替代对鱼体造成不同的影响可能是不同脂肪源中脂肪酸组成差异引起的。因此, 通过多种植物油混合达到平衡的脂肪酸模式可能是提高植物脂肪源替代鱼油效果的潜在方式。
异育银鲫(Carassius gibelio)是我国重要的大宗淡水鱼养殖品种, “中科3号”和“中科5号”是目前广泛推广的两个重要品种。前期研究表明, 胰岛素对不同品种异育银鲫脂肪代谢的调控存在显著差异[9]。此外, 不同品种异育银鲫在饲料不同糖、脂水平下,对脂肪代谢响应存在品种差异性[10]。因此, 我们推测异育银鲫“中科3号”和“中科5号”对饲料脂肪源的利用可能存在差异。本实验以异育银鲫“中科3号”和“中科5号”为研究对象, 参考鱼油的脂肪酸组成配制混合植物油饲料, 评估不同饲料脂肪源对不同品种异育银鲫生长性能、鱼体基本组分、肌肉脂肪酸组成、血浆生化及脂肪代谢(脂肪合成、脂肪分解、脂肪酸β氧化和脂肪酸转运)的影响。本研究可为植物脂肪源替代鱼油提供数据支撑, 并为高效利用植物脂肪源的鱼类遗传选育提供理论基础。
1 材料与方法
1.1 实验饲料
本实验以鱼油为基础参照, 将菜籽油、大豆油、棕榈油按照55%∶25%∶20%的比例调配至与鱼油相似的脂肪酸组成。分别以鱼油(FO)和混合植物油(BVO)为脂肪源, 配制两种等氮(35%)等脂(9%)饲料, 实验饲料配方见表1。所有原料粉碎后经40目筛网过筛, 按配方比例充分混匀, 再加水搅拌后用制粒机(SLP-45, 上海渔业机械设备研究所,中国)制备成2 mm的饲料颗粒。为防止饲料中的脂肪酸氧化, 以上饲料在烘箱50℃烘干后于-20℃冰柜中保存备用。饲料脂肪酸组成见表2。
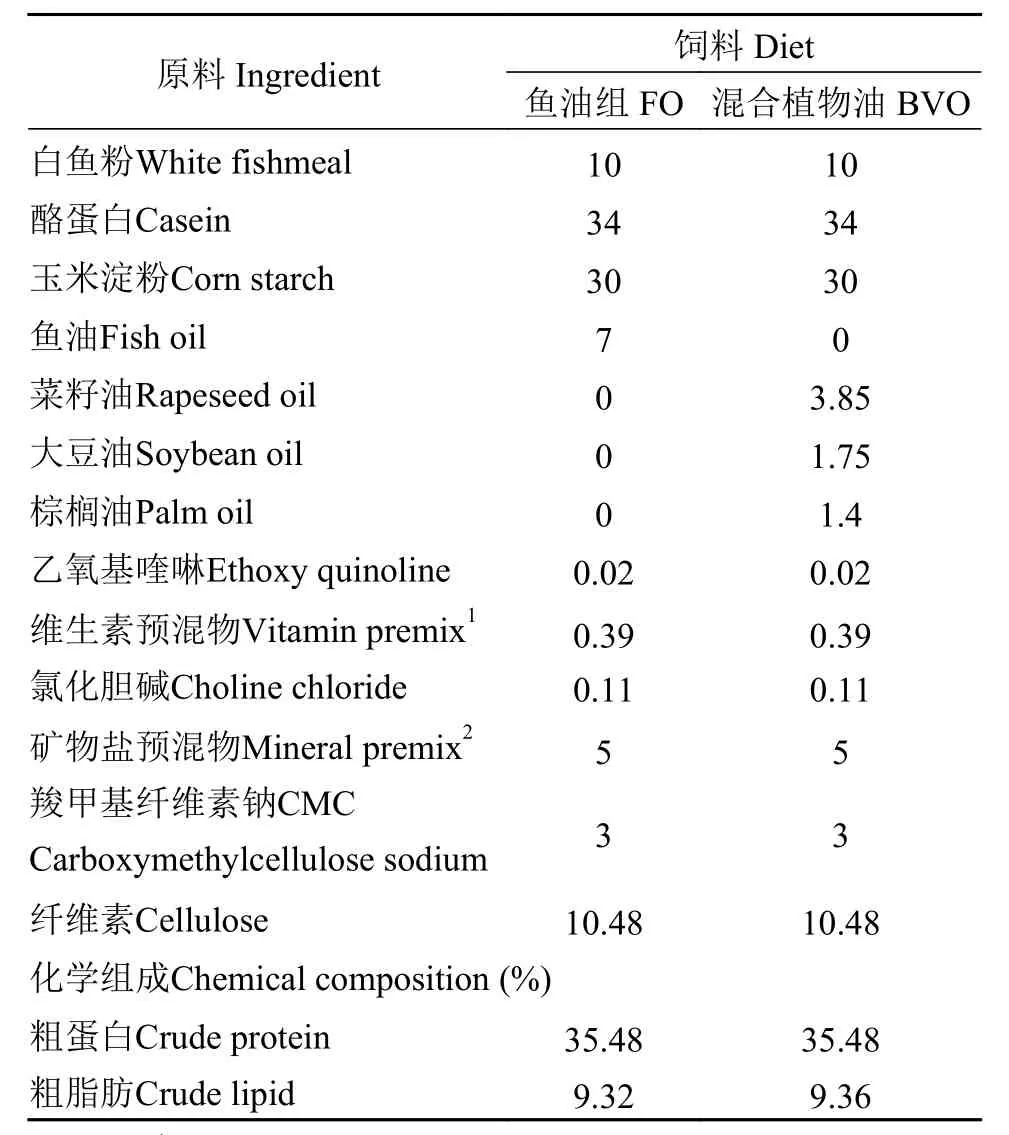
表1 饲料配方和化学组成(% 干物质)Tab. 1 Formulation and chemical composition of the experimental diets (% dry matter)
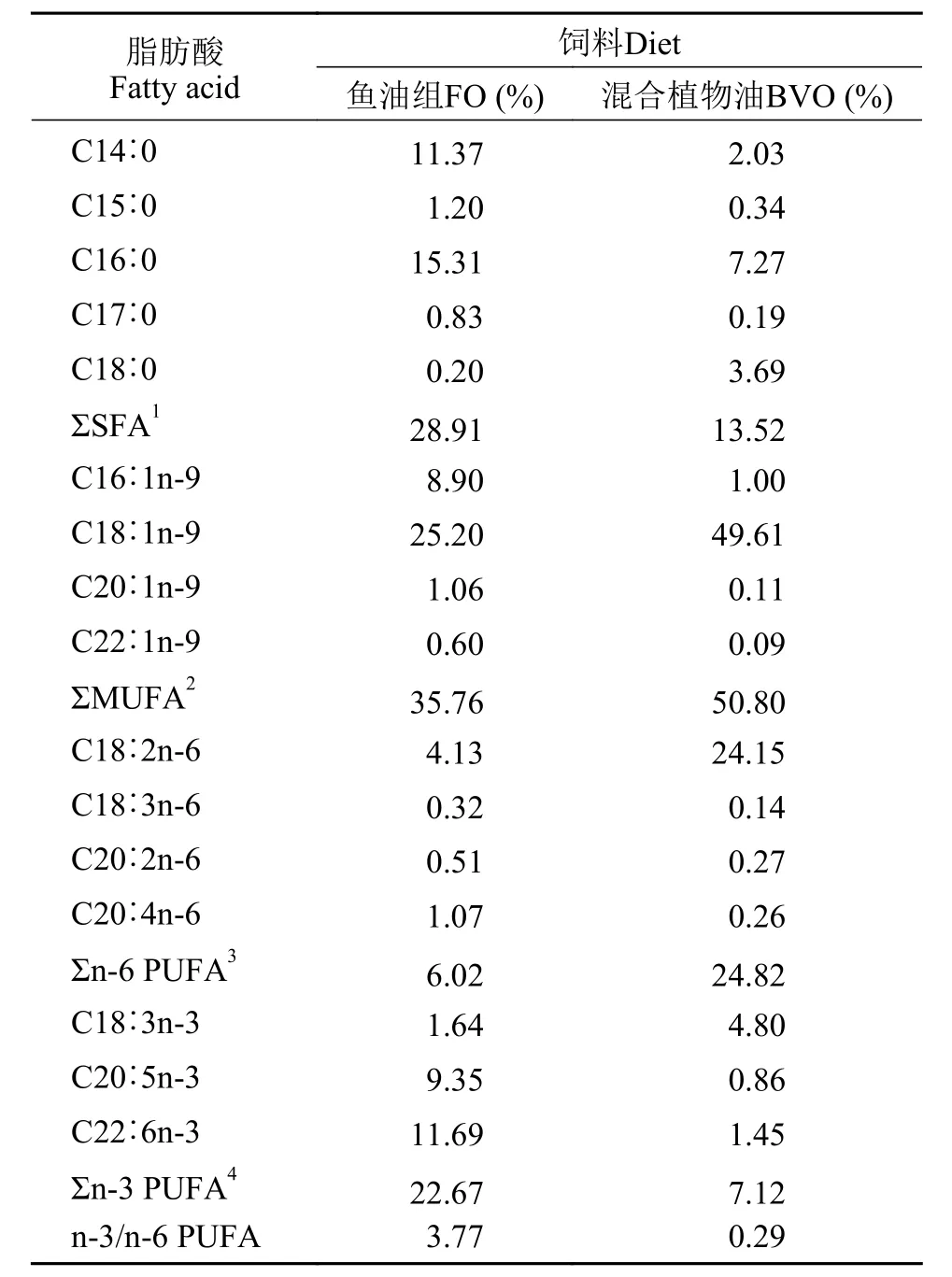
表2 饲料脂肪酸组成(% 总脂肪酸)Tab. 2 Fatty acid composition of the experimental diets (% of total fatty acids)
1.2 实验鱼及饲养
实验所需的异育银鲫“中科3号”(A strain)和“中科5号”(F strain)幼鱼均由中国科学院水生生物研究所(湖北, 武汉)提供。在正式实验前, 所有的鱼都在室内循环水养殖系统中暂养2周以适应养殖环境,期间投喂2种饲料的等量混合物。将实验鱼饥饿24h后, 随机挑取规格相近、外观健康的不同品种异育银鲫[“中科3号”: (5.01±0.05) g和“中科5号”:(5.02±0.05) g], 称重后放入养殖缸中进行养殖实验。每缸25尾, 每个处理3个平行。养殖实验持续70d, 每天表观饱食投喂3次(8:30、13:30和18:30)。实验期间的水温为(28.99±0.47)℃, 溶氧>7 mg/L, 氨氮<0.1 mg/L, pH为6.5—7.0。光照周期为12L∶12D(8:00—20:00光亮), 水面光照强度为2.79—3.32 μmol/(s·m2)。
1.3 样品采集
在养殖实验结束后, 将鱼饥饿8h后取样。用麻醉剂MS-222 (60 mg/L, Sigma, USA)将各处理缸中的实验鱼麻醉后计数称重, 随后进行取样。每缸随机挑选2尾鱼, 用肝素钠抗凝剂润过的无菌注射器从尾静脉取血并放入1.5 mL灭菌离心管中, 3000×g离心10min后得到血浆, 保存于-80℃用于后期分析。同时, 在冰浴条件下解剖实验鱼, 快速取得肝脏和肌肉样品, 将其放入液氮中速冻并保存在-80℃。另外, 取肌肉组织样品, 经冷冻干燥处理后用于测定脂肪酸组成。
1.4 样品分析方法
参照AOAC方法[11]分析实验饲料和鱼样的生化组成(水分、灰分和脂肪)。干物质在烘箱(电热恒温干燥箱, 精宏, 中国上海)于105℃干燥至恒重,通过失重法测定; 样品在550℃下的马弗炉(马弗炉,中国湖北)中燃烧3h至恒重, 采用失重法测定灰分;粗脂肪分析以乙醚作为抽提液, 在索氏抽提仪(Soxtec System HT6, Tecator, Haganas, Sweden)中进行。参照Folch等[12]方法, 使用氯仿∶甲醇=2∶1混合试剂提取饲料和肌肉组织总脂, 用气相色谱仪来测定脂肪酸的组成和含量(7890A-5975C, Agilents Technologies Inc., Santa Clara, CA, USA)。血浆葡萄糖、甘油三酯和胆固醇含量采用商品试剂盒(Fujifilm,Wako Pure Chemical Corporation, Osaka, Japan)进行测定。血浆低密度脂蛋白胆固醇(LDL-C)和高密度脂蛋白胆固醇(HDL-C)分别采用南京建成商品试剂盒A113-1-1和A112-1-1检测(南京建成生物工程研究所, 中国南京)。
1.5 基因表达分析
使 用TRIzol试 剂(Invitrogen, Carlsbad, California, USA)从肝脏和肌肉组织中提取总RNA, 通过逆转录试剂盒M-MLV First-Strand Synthesis Kit (Invitrogen, USA)将其逆转录为cDNA。实时荧光定量在Light Cycle 480 Ⅱ(Roche, Basel, Switzerland)仪器 上 进 行, 采 用SYBR®Green Ⅰ Master (Roche,Basel, Switzerland)荧光染色剂测定肝脏和肌肉中脂肪代谢相关基因的相对表达量。选择β-actin为内参基因, 基因相对表达量分析的计算方法参照Pfaffl[13], 引物信息见表3。

表3 本实验所用引物序列表Tab. 3 Primers used in the present study
1.6 数据分析
所有数据使用统计软件SPSS 18.0 (SPSS Inc.,Chicago, IL, USA)进行统计分析, 结果以均值±标准误(mean±SE,n=6)表示。采用双因素方差分析(Twoway ANOVA)来比较饲料(鱼油vs.混合植物油)和品种(中科3号vs.中科5号)两个因素的作用。当存在交互作用时(P<0.05), 用Student-Newman-Keuls多重比较分析各组间差异, 用小写字母a、b或c来表示各组之间差异。当不存在交互作用时(P>0.05), 用大写字母A和B表示品种间的差异, 大写字母X和Y表示饲料间的差异。
2 结果
2.1 混合植物油替代鱼油对不同品种异育银鲫生长和饲料利用的影响
如表4所示, 异育银鲫摄食混合植物油的摄食率均显著高于鱼油组, 但饲料效率显著降低(P<0.05)。与“中科3号”相比, “中科5号”摄食率显著降低, 但饲料效率显著增加(P<0.05)。摄食鱼油饲料组“中科5号”特定生长率显著高于其他处理组(P<0.05)。
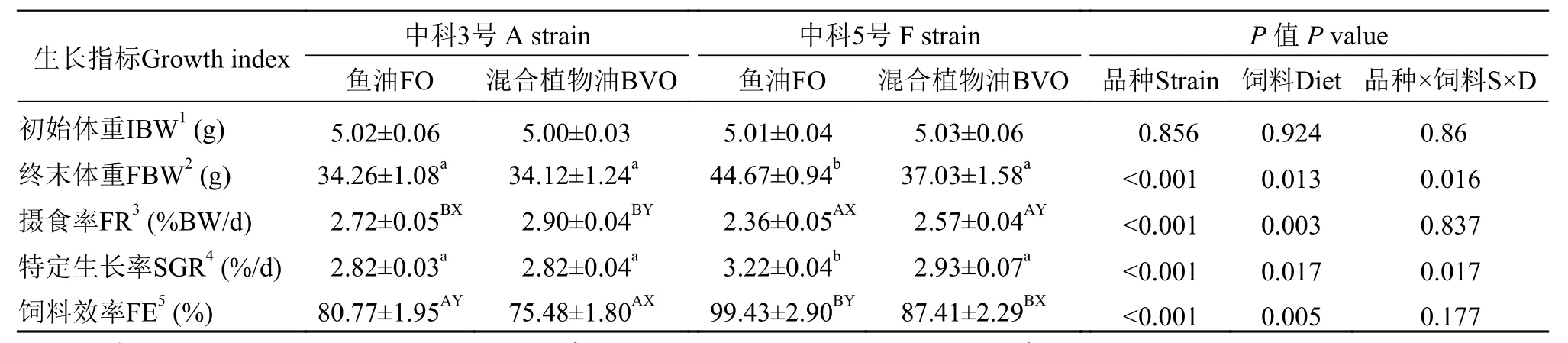
表4 饲料鱼油和混合植物油对不同品种异育银鲫生长和饲料利用的影响Tab. 4 Effects of dietary fish oil and blended vegetable oil on growth and feed utilization of two strains of gibel carp
2.2 混合植物油替代鱼油对不同品种异育银鲫鱼体生化和肌肉脂肪酸组成的影响
如表5所示, 异育银鲫摄食混合植物油饲料组鱼体粗脂肪含量显著高于鱼油组(P<0.05)。异育银鲫“中科3号”粗脂肪含量显著高于“中科5号”, 而水分含量显著低于“中科5号”(P<0.05)。灰分含量在各处理间无显著性差异(P>0.05)。

表5 饲料鱼油和混合植物油对不同品种异育银鲫鱼体基本组分的影响Tab. 5 Effects of dietary fish oil and blended vegetable oil on body composition of two strains of gibel carp
如表6所示, 与混合植物油组相比, 不同品种异育银鲫摄食鱼油饲料组Σn-6 PUFA相对含量显著降低, 而Σn-3 PUFA相对含量显著增加(P<0.05)。
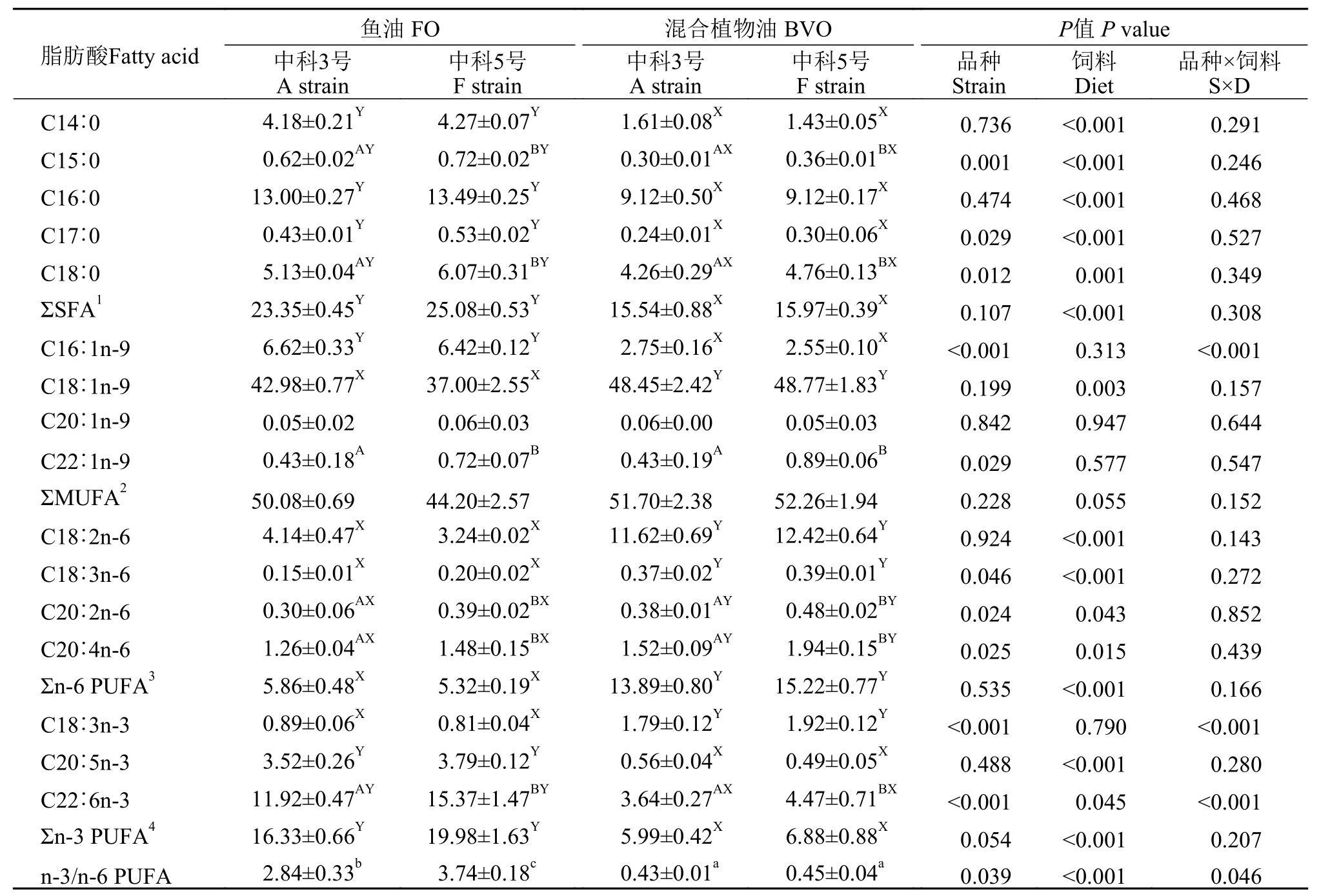
表6 饲料鱼油和混合植物油对不同品种异育银鲫肌肉脂肪酸组成的影响Tab. 6 Effects of dietary fish oil and blended vegetable oil on fatty acids composition in the muscle of two strains of gibel carp
2.3 混合植物油替代鱼油对不同品种异育银鲫血浆生化指标的影响
如表7所示, 不同脂肪源饲料对“中科3号”血糖含量无显著性影响, 而“中科5号”摄食混合植物油饲料组血糖含量显著高于“中科3号”(P<0.05)。“中科3号”血浆低密度脂蛋白胆固醇含量显著高于“中科5号”(P<0.05)。不论摄食何种饲料, 不同品种异育银鲫血浆甘油三酯、胆固醇和高密度脂蛋白胆固醇含量无显著性差异(P>0.05)。
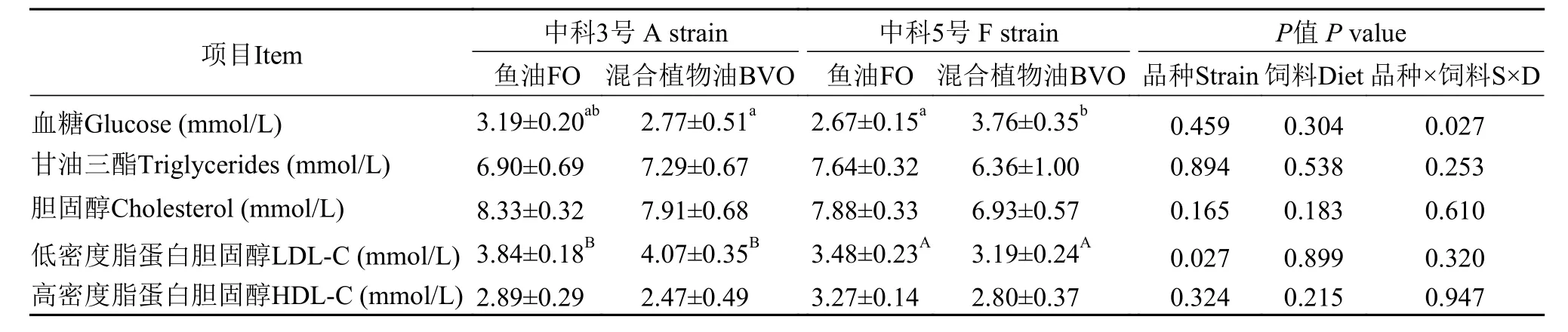
表7 饲料鱼油和混合植物油对两个品种异育银鲫血浆生化指标的影响Tab. 7 Effects of dietary fish oil and blended vegetable oil on plasma biochemical indices of two strains of gibel carp
2.4 混合植物油替代鱼油对不同品种异育银鲫肝脏和肌肉脂肪代谢相关基因表达的影响
不同品种异育银鲫肝脏和肌肉脂肪合成关键调控元件pparγ、srebp-1c及下游靶基因acly、acc和fas的基因表达量无显著性差异(图1A)。不同品种异育银鲫摄食混合植物油饲料组肌肉srebp-1c和acc基因相对表达量显著高于鱼油组(P<0.05, 图1B)。
不同品种异育银鲫摄食鱼油和混合植物油饲料后肝脏(图2A)和肌肉(图2B)脂肪分解代谢相关基因表达结果显示, 鱼油组肝脏脂肪酸β氧化关键基因pparα表达量显著高于混合植物油组(P<0.05)。异育银鲫“中科5号”肌肉脂肪分解相关基因hsl表达量显著高于“中科3号”, 且脂肪酸β氧化相关基因aco3在肝脏和肌肉中的表达量均显著高于“中科3号”(P<0.05)。脂肪分解代谢相关基因lpl(肝脏、肌肉)和cpt1a(肝脏、肌肉)、hsl(肝脏)和pparα(肌肉)的表达量均不受品种和饲料脂肪源的影响(P>0.05)。
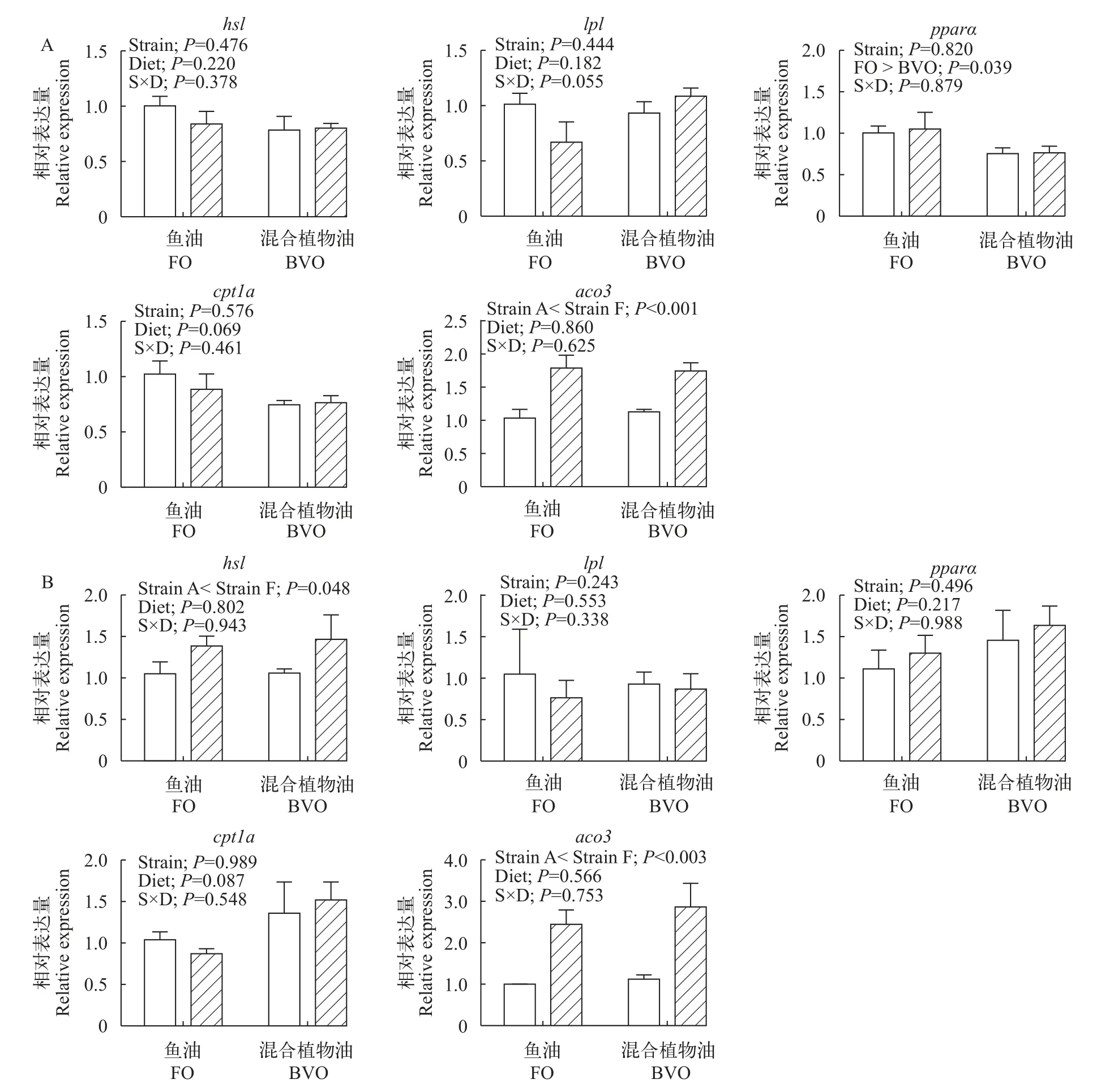
图2 饲料鱼油(FO)和混合植物油(BVO)对“中科3号”(空白柱)和“中科5号”(斜线柱)肝脏(A)和肌肉(B)脂肪分解和脂肪酸β氧化相关基因相对表达量的影响Fig. 2 Effects of dietary fish oil (FO) and blended vegetable oil (BVO) on the mRNA levels of gene involved in lipolysis and fatty acids β oxidation in the liver (A) and muscle (B) of gibel carp A strain (blank bars) and F strain (bias bars)
不论摄食何种脂肪源饲料, 异育银鲫“中科5号”肝脏脂肪酸转运相关基因fabp1b和fatp1的表达量均显著高于“中科3号”(图3A)。饲料脂肪源和品种对异育银鲫肌肉fabp1b和fatp1的表达均无显著性影响(图3B)。
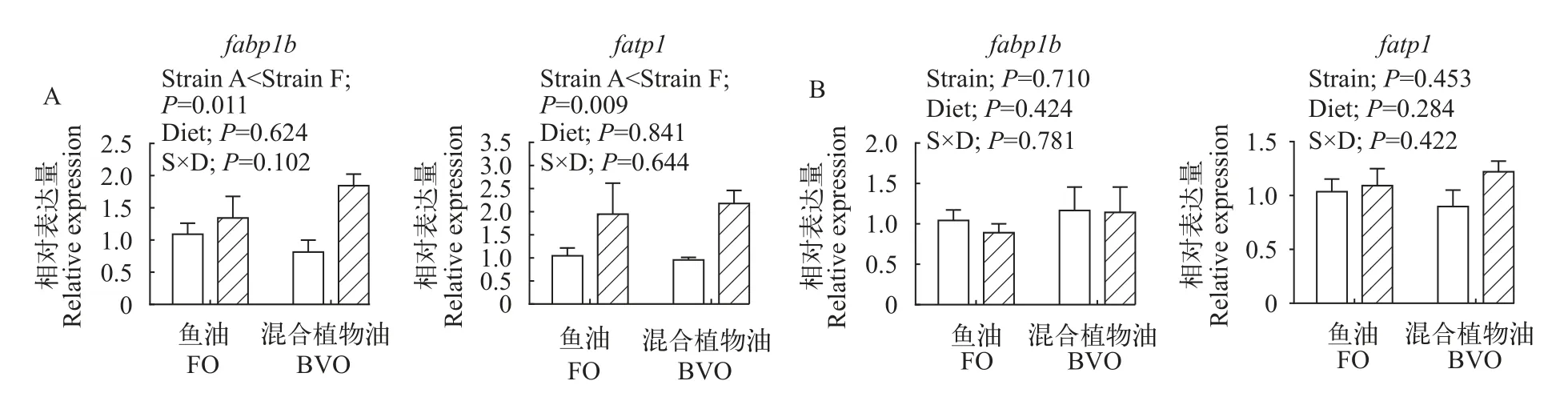
图3 饲料鱼油和混合植物油对“中科3号”(空白柱)和“中科5号”(斜线柱)肝脏(A)和肌肉(B)脂肪酸转运相关基因相对表达量的影响Fig. 3 Effects of dietary fish oil (FO) and blended vegetable oil (BVO) on the mRNA levels of gene involved in fatty acids transportation in the liver (A) and muscle (B) of gibel carp A strain (blank bars) and F strain (bias bars)
3 讨论
3.1 饲料脂肪源作用
本实验结果显示, 异育银鲫“中科3号”和“中科5号”摄食混合植物油饲料组摄食率显著高于鱼油组, 但饲料效率显著低于鱼油组, 表明异育银鲫对鱼油的利用优于混合植物油, 这与在鲑鳟鱼类中的研究结果一致[14,15]。陈家林等[16]的研究结果表示,与全鱼油组相比, 鱼油分别与椰子油、玉米油、亚麻油1∶1的混合替代显著提高异育银鲫的生长性能,但对饲料效率无显著影响。李新等[17]发现, 吉富罗非鱼(Oreochromis niloticus)摄食棕榈油饲料组摄食率显著高于棉籽油组, 但饲料效率无显著差异。饲料不同脂肪源对细鳞鲑(Brachymystax lenok)摄食率无显著性影响, 其中鱼油组的饲料效率最高[2]。紫苏油、菜籽油、葵花油和棕榈油完全替代鱼油对红罗非鱼(Oreochromissp.)生长性能无显著影响,棕榈油组的饲料转化率显著高于鱼油组[18]。因此, 鱼类对植物油的利用差异与鱼体本身(品种和规格)、脂肪源的种类和养殖条件等多种因素有关。不同品种异育银鲫摄食混合植物油饲料摄食率显著高于鱼油组, 可能是通过增加摄食来满足自身的能量需求。
在本研究中, 不同脂肪源对不同品种异育银鲫血浆代谢物水平(血糖、甘油三酯、胆固醇、低密度脂蛋白胆固醇和高密度脂蛋白胆固醇)无显著性影响。饲料脂肪酸组成影响鱼类的脂肪合成、分解和转运等脂肪代谢过程, 肝脏作为鱼类代谢调节的中枢器官在脂肪代谢过程中发挥重要作用。在虹鳟(Oncorhynchus mykiss)中, 植物脂肪源完全替代鱼油增强了肝脏脂肪合成相关基因的表达[19]。不同品系大西洋鲑(Salmo salar)摄食鱼油和混合植物油饲料后, 肝脏长链多不饱和脂肪酸代谢存在显著差异, 表现为Lean品系肝脏脂肪合成关键基因脂肪酸合成酶的表达显著高于Fat品系[20]。混合植物油(菜籽油∶棕榈油∶亚麻籽油=55%∶30%∶15%)完全替代鱼油对虹鳟肝脏脂肪合成无显著性影响[21]。由此可见, 鱼的种类和饲料脂肪酸组成影响鱼体的脂肪代谢过程。在本实验中, 摄食混合植物油饲料组异育银鲫肌肉脂肪合成关键基因srebp1和acc的表达量显著上调, 且鱼体脂肪含量显著增高, 表明不同品种异育银鲫摄食混合植物油饲料表现出更高的脂肪合成潜能。同样, 不同品系大西洋鲑在摄食混合植物油饲料后, Lean品系大西洋鲑脂肪酸合成酶的基因表达水平较鱼油组显著上调[22]。脂肪合成的增加可能和饲料中n3/n6值有关, n-3 PUFA和n-6 PUFA比例的不平衡可能是造成肝脏脂肪含量增加的原因[23]。另外, 鱼体的脂肪酸组成可以反映饲料脂肪酸组成[24,25]。在本实验中, 不同品种异育银鲫摄食鱼油饲料组肌肉中n-3 PUFA的含量显著高于摄食混合植物油饲料组, 而肌肉中n-6 PUFA的含量结果相反, 表现出与饲料中脂肪酸含量的一致性, 验证了肌肉脂肪酸体现饲料脂肪酸组成这一结论。
3.2 品种效应
无论摄食何种脂肪源饲料, “中科5号”摄食率显著低于“中科3号”, 而饲料效率显著高于“中科3号”, 表现出较好的饲料利用性能。“中科5号”摄食不同脂肪源饲料体脂肪含量显著低于而水分含量显著高于“中科3号”, 与不同品种异育银鲫摄食不同水平碳水化合物饲料和不同蛋白源饲料的结果一致[26,27]。机体脂肪积累是一个复杂的生理过程, 包含脂肪的转运、摄取、合成和分解代谢过程等。在脂肪合成方面, 不同品种异育银鲫肝脏和肌肉脂肪合成相关基因均无显著差异。“中科5号”肌肉脂肪分解相关基因hsl和脂肪酸β氧化酶基因aco3的相对表达量显著高于“中科3号”, 表明“中科5号”通过脂肪分解和脂肪酸β氧化过程导致了较低的体脂肪含量。该研究结果与不同品种异育银鲫摄食不同饲料糖、脂水平的研究结果一致, 表明“中科5号”脂肪分解和脂肪酸氧化潜力优于“中科3号”[10]。另外, “中科5号”肝脏中脂肪酸转运结合蛋白fabp1b和负责脂肪酸跨膜转运的脂肪酸转运蛋白1 (fatp1)的基因表达量均显著高于“中科3号”,表明“中科5号”肝脏组织中脂肪酸的转运摄取的潜力较强。综上所述, 异育银鲫“中科5号”对脂肪的转运、分解、脂肪酸β氧化水平均显著高于“中科3号”, 表现出对饲料脂肪更好的利用能力。
3.3 脂肪源和品种的交互作用
在本实验中, 饲料脂肪源和品种对异育银鲫特定生长率的影响存在交互作用。然而, 混合植物油完全替代鱼油对“中科3号”的特定生长率无显著性负面影响, 且“中科5号”摄食混合植物油可以达到与“中科3号”摄食鱼油组一致的生长性能, 表明在“中科5号”饲料中使用植物性脂肪源存在巨大潜能。血液生化指标能够反映鱼体的生理状态。在本研究中, 异育银鲫血糖水平受到饲料脂肪源和品种的交互作用影响。与鱼油组相比, 异育银鲫“中科5号”混合植物油组的血糖含量显著高于“中科3号”, 可能是由于“中科5号”倾向于利用脂肪酸氧化分解来为机体供能, 而非依赖于血糖的分解代谢供能。另外, 品种和饲料脂肪源的交互作用也体现在肌肉脂肪酸组成上, “中科5号”摄食鱼油组n-3/n-6 PUFA显著高于其他各组, 表明“中科5号”在利用鱼油时体现出的更优质的肌肉脂肪酸组成。
4 结论
本实验探究了饲料混合植物油完全替代鱼油对不同品种异育银鲫生长和脂肪代谢的影响。与鱼油组相比, 混合植物油组显著提高了不同品种异育银鲫的摄食率, 但饲料效率有所降低, 表明植物油脂肪源对不同品种异育银鲫的饲料利用存在一定的负面影响。在饲料脂肪源效应上, 混合植物油饲料通过增强肌肉组织脂肪合成相关基因srebp-1c和acc的表达来促进体脂肪的累积。在品种效应上, 不同品种异育银鲫在体脂含量和肌肉脂肪代谢上表现出品种特异性。“中科5号”体脂肪含量显著低于“中科3号”, 可能是其具有较高的肌肉脂肪分解、脂肪酸氧化及脂肪酸转运能力导致的。饲料脂肪源和品种对异育银鲫生长性能的影响存在交互作用, 但“中科5号”摄食混合植物油饲料可以达到与“中科3号”摄食鱼油组相似的特定生长率, 且具有较高的肌肉n-3/n-6 PUFA水平, 表明混合植物油在“中科5号”饲料中具有广泛应用的潜力。
猜你喜欢
垂钓(2023年10期)2024-01-02
信息安全与通信保密(2023年7期)2023-09-25
信息安全与通信保密(2023年5期)2023-08-15
中国化肥信息(2021年7期)2021-11-06
中国生殖健康(2020年5期)2021-01-18
科学养鱼(2020年5期)2020-12-20
当代水产(2019年8期)2019-10-12
中国生殖健康(2018年5期)2018-11-06
商周刊(2017年6期)2017-08-22
食品界(2016年4期)2016-02-27
