
发酵乳后酸化的影响因素及其控制措施
2024-02-26 09:21:56朱曼李宝磊侯保朝洪维錬何剑张宇姜毓君满朝新
食品研究与开发 2024年4期
朱曼,李宝磊,侯保朝,洪维錬,何剑,张宇,姜毓君,满朝新*
(1.东北农业大学食品学院乳品科学教育部重点实验室,黑龙江哈尔滨 150030;2.国家乳业技术创新中心,上海 201111)
发酵乳是指牛(羊)乳或乳粉经乳酸菌发酵工艺而制成的一类乳制品[1],其含有丰富的活性蛋白质、碳水化合物以及维生素和矿物质,并且具有较高的生物利用率,对人体健康具有积极的影响,因此发酵乳在人们日常生活中的地位逐渐提高[2]。近年来,我国乳制品行业持续发展,产量稳步提升。发酵乳制品虽然对人体具有积极的影响,但因其活性乳酸菌代谢活动的持续进行,产品在储存、运输以及销售过程中易产生后酸化现象,严重影响产品的品质。
后酸化是指发酵乳在发酵结束后至食用前的阶段中,因乳酸菌继续生长导致发酵乳的pH 值不断降低,出现过酸味甚至导致感官质量下降的现象。后酸化除了缩短货架期外,还会导致许多缺陷,例如乳清脱水、乳酸菌数量减少同时不利于益生菌在发酵乳中的稳定性。发酵乳产生后酸化现象是制约发酵乳发展的关键因素之一[3]。本文将通过分析导致后酸化的因素来揭示可能产生后酸化的机制,并对减弱后酸化的策略(物理、化学和微生物措施)进行综述,以期为减弱发酵乳后酸化提供一些可供参考的思路,为解决这一制约发酵乳制品行业的难题提供方向。
1 影响后酸化的因素
发酵乳制品的后酸化程度会受到多种因素的影响:发酵剂类型、乳的成分、温度和pH 值、均质与搅拌、益生菌和益生元。
1.1 发酵剂类型
发酵剂是指用于酸奶、奶酪、纳豆和其他发酵产品生产的细菌以及其他微生物的培养物。根据发酵途径可分为同型发酵、兼性异型发酵和异型发酵3 种发酵类型。
保加利亚乳杆菌是导致发酵乳后酸化的主要菌株。在发酵过程中,当pH 值降低到5.0 左右时,保加利亚乳杆菌成为造成后酸化现象的优势菌株。关于后酸化对发酵乳品质的影响因素研究发现,具有球菌/杆菌的高比例发酵剂的应用可减弱发酵乳贮藏期的酸度下降程度[4],因为保加利亚乳杆菌的细胞膜或者细胞壁对乳糖酶活性具有保护作用,使乳糖酶贮藏过程中持续产生乳酸,导致后酸化现象的发生。Akgun 等[5]在研究不同的发酵剂类型对水牛奶酸奶品质的影响时,发现发酵剂类型改变了水牛奶酸奶样品在冷藏过程中的后酸化动力学,显著影响了冷藏过程中乳酸、乙酸和柠檬酸的产生。综上,发酵剂的类型可以影响发酵乳制品的后酸化程度,从改变发酵剂类型的角度出发,有望找到降低后酸化现象的新途径。
1.2 发酵乳的成分
酸化和酸化后的动力学取决于不同动物的乳、乳成分等的相互作用。羊乳在酸奶生产过程中表现出更快的酸化,在储藏第29 天时,羊乳的pH 值为3.9,而牛乳与羊乳的体积比为1∶1 时的pH 值为4.1,而且羊乳的后酸化程度高于牛乳[6]。由此可以看出乳成分对发酵乳后酸化有一定的影响,羊乳后酸化程度高与羊乳中乳糖的含量有关,由于羊乳中的乳糖含量高于牛乳[7],乳糖在储藏期可继续发酵产生乳酸使得羊乳的后酸化现象增加。发酵前向乳中添加物质也可改变发酵乳的后酸化程度。发酵前将胡萝卜汁添加到脱脂乳中显著降低发酵乳的pH 值,在贮藏期内后酸化作用增强,可滴定酸含量增加,胡萝卜汁的添加加重了后酸化,这是由于胡萝卜汁中含有的黏性多糖等膳食纤维在发酵过程中促进乳酸菌的增殖[8]。
个别牛奶成分的修饰尤其是蛋白质的改变可介导发酵前和发酵后的酸化动力学。在发酵前用转谷氨酰胺酶处理脱脂奶和全脂奶(40 ℃/2 h)可降低酸奶在25 d 贮藏期内的后酸化程度,原因是酶处理使得蛋白质凝胶孔径减小、蛋白质分布规则、(γ-谷氨酸)-赖氨酸键的持水能力提高,从而改善了发酵乳的保水性能。同时也导致微生物生长所需的低分子量肽减少,使得产酸菌的生长减缓,产生乳酸减少,从而降低发酵乳的后酸化[9]。因此,乳成分的种类、乳中添加的物质以及乳成分的改变都会影响发酵乳的后酸化。
1.3 温度
发酵温度可能通过影响微生物生长来影响发酵乳的后酸化程度。Sanusi 等[10]的研究表明,相较于低发酵温度(39 ℃)的复合发酵乳,较高发酵温度(41~43 ℃)的复合发酵乳酸化速率提高。除了发酵温度外,冷藏温度对发酵乳后酸化的控制也起到关键的作用。发酵乳在后发酵过程中迅速冷却至冷藏条件(<10 ℃),是通过限制发酵剂的代谢活性来控制其最终pH 值(4.0~4.4)的最关键因素。但是由于微生物的残留活性,此方法仍然无法防止后酸化。低温可以抑制发酵乳中微生物的活性,降低微生物发酵乳糖产生乳酸的能力;高温则能增强微生物发酵乳糖产生乳酸的能力,从而加重发酵乳后酸化的程度。
1.4 均质和搅拌
发酵乳生产过程中,一般在温度55~65 ℃、压力10~18 MPa 的条件下对牛奶进行均质处理,使得脂肪球的大小减小,从而增加了脂肪与其他成分(如酪蛋白和乳清蛋白)之间的接触面积。这使得脂肪能够更好地结合到蛋白质的网络中,脂肪的结合能力增强,从而在酸化过程中能更好地保持脂肪与酪蛋白和乳清蛋白的结合,从而更好地改善乳的凝胶特性[11]。冷冻/解冻的发酵乳的后酸化更为严重,Tribst 等[12]研究表明,均质后的发酵乳在贮藏28 d 时的酸度值较未均质发酵乳有所降低。Sert 等[13]的研究表明高压均质后的绵羊奶酸奶酸化速率提高,并在贮藏第14 天和第28 天时,经过均质后的绵羊奶发酵乳的pH 值降低。虽然以上的研究表明均质对发酵乳的后酸化有一定的作用,但是出现了不同的影响效果,均质工艺对后酸化的影响可能基于不同的基质和发酵体系表现出不同的影响作用,仍需要展开进一步的探究。
按照组织状态和制备方法,发酵乳分为搅拌型和凝固型两种类型。相较于在容器中发酵的凝固型发酵乳来说,搅拌型发酵乳具有高黏度以及光滑的质地[14]。将凝固型酸奶转化为搅拌型酸奶后,显示出更好的保水能力,但由于搅拌型凝胶在冷藏后酸化过程中能够恢复其结构,因此质地粗糙且呈颗粒状。
1.5 益生菌和益生元
益生菌是通过定殖在人体内从而改变宿主某一部位菌群组成的一类活性微生物,益生菌数量充足时能够促进宿主的身体健康。发酵乳制品中添加益生菌不仅能够提供充足的营养,还可在发酵阶段促进益生菌的生长。Zhang 等[15]的研究表明LactobacillusplantarumK25 和酸奶发酵剂制成的益生菌发酵乳比仅用酸奶发酵剂制成的发酵乳风味好,且添加益生菌后该发酵乳在冷藏过程中后酸化程度降低。添加Lactobacillusacidophilus(DD910)和Bifidobacteriumlactis(DD920)降低了酸奶贮藏过程中乳酸的形成,在一定程度上降低酸奶后酸化程度[16]。单一益生菌或多种益生菌组合在贮藏过程中表现出不确定的pH 值变化,但对降低发酵乳后酸化程度有一定的效果。
益生元是指一些不被宿主消化吸收却能有选择性地促进其体内有益菌的生长和繁殖,从而改善宿主健康的有机物质,包括功能性低聚糖类、多糖类等,其中功能性低聚糖类是最常见的益生元。Yang 等[17]研究表明添加0.1%委陵菜多糖(Potentillaanserinepolysaccharide,PAP)可延缓牦牛酸奶在储存过程中的后酸化过程。有研究表明益生元参与放大发酵行为和后酸化速率。乳果糖与益生菌共培养提高发酵脱脂乳的品质,与不添加乳果糖的对照发酵相比,在脱脂乳中添加乳果糖增加了所有益生菌的数量,提高发酵乳的酸化速率和乳酸酸度,同时缩短了发酵时间并降低了冷藏结束1~35 d 的pH 值[18]。综上所述,益生元可以放大发酵乳制品中的后酸化,主要是因为它们被特定的益生菌菌株选择性利用。
2 后酸化的产生机制
乳酸菌等细菌在酸奶等酸性环境中主要是通过从细胞中排出H+来维持pH 值稳态。研究发现:当外界环境的pH 值低于胞内正常范围的pH 值且不超过一定程度时,乳酸菌能够依靠自身的耐酸机制抵御外界的干扰。但当环境介质的pH 值过低时,乳酸菌的耐酸机制可能无法有效应对,导致细胞内的pH 值稳态无法维持,这会影响其正常的新陈代谢过程。细胞的新陈代谢紊乱会导致重要的生物化学反应受阻,细胞无法正常运作。如果pH 值过低的环境持续存在,最终导致乳酸菌死亡。乳杆菌用以维持细胞内pH 值稳态的主要机制是谷氨酸脱羧酶(glutamate decarboxylase,GAD)机制和FoF1-三磷酸腺苷酶(adenosine triphosphatase,ATPase)。
2.1 GAD
GAD 系统是乳杆菌维持胞内pH 值稳态的有效途径之一,谷氨酸脱羧酶是GAD 系统的核心酶,可以催化谷氨酸发生脱羧反应生成γ-氨基丁酸(γ-aminobutyric acid,GABA),GABA 在反向运输系统作用下运至细胞外,使得细胞内H+减少,pH 值升高,维持细胞内pH 值稳态。因为H+被排到外界环境中,从而使得外界环境的pH 值下降,导致发酵乳制品后酸化的产生[19]。
有研究表明:添加转谷氨酰胺酶(transglutaminase,TG)的低脂发酵乳中,TG 能够减慢发酵乳的后酸化过程[20]。同样,Chr Lorenzen 等[9]的研究也表明了TG酶和嗜热酸奶发酵剂发酵的发酵乳在储藏期间的后酸化程度较低。TG 对菌株的生长有一定的抑制作用,因此在发酵乳中添加TG 可抑制保加利亚乳杆菌的生长,降低发酵乳的后酸化[21]。TG 具有促使蛋白质水解的功能,使得菌株因营养缺乏生长受到阻碍,使得发酵乳在贮藏期后酸化程度降低。但是发酵菌株本身不能生产TG,仅能通过人为添加,这也无形之中增加了产品生产的成本。
2.2 FoF1-ATPase
在FoF1-ATPase 中,F1部分位于细胞质侧,其α、β、γ、δ、ε 5 类亚基共同形成一个结构,这个结构在酶催化二磷酸腺苷(adenosine diphosphate,ADP)和无机磷酸转变为三磷酸腺苷(adenosine triphosphate,ATP)的过程中起到关键作用。而Fo部分则嵌入在细胞膜中,它由a、b、c 3 个亚基组成,形成一个质子通道,负责质子转运。
当乳酸菌处于低pH 值环境时,FoF1-ATPase 会增加其酶活性,通过排出胞内的H+来提高活力,并维持跨膜pH 值梯度。这个跨膜pH 值梯度对于维持细胞内环境的稳定非常重要,它参与调控多种细胞功能和代谢过程。然而,当胞内pH 值进一步下降时,FoF1-ATPase 的酶活性也会急剧下降。这可能是由于胞内pH 值低迫使酶的构象发生改变,从而导致其催化能力降低。当FoF1-ATPase 活性下降,乳酸菌可能无法有效排出H+,维持跨膜pH 值梯度,这会对胞内pH 值平衡产生一定的影响,并可能进一步影响细胞的代谢和生存能力[22]。而质子转移ATP 酶(plasma membrane ATPase,H+-ATPase)活性作为可逆的离子转位泵,催化氢离子穿过细胞膜,使细胞内的pH 值保持中性,导致发酵乳后酸化的产生。因此,很多学者通过筛选具有低H+-ATPase 活性的保加利亚乳杆菌来作为发酵菌株,以此来降低发酵乳制品在贮存期间的后酸化。
3 后酸化的控制措施
发酵乳在储藏和运输过程中的冷藏温度(2~10 ℃)是防止后酸化的首要步骤。但是资源贫乏的国家和地区对制冷条件无法持续维持,本文将对除了制冷条件外的后酸化的控制措施进行阐述,并将其分为三大类:物理措施、化学措施以及生物措施(见图1)。
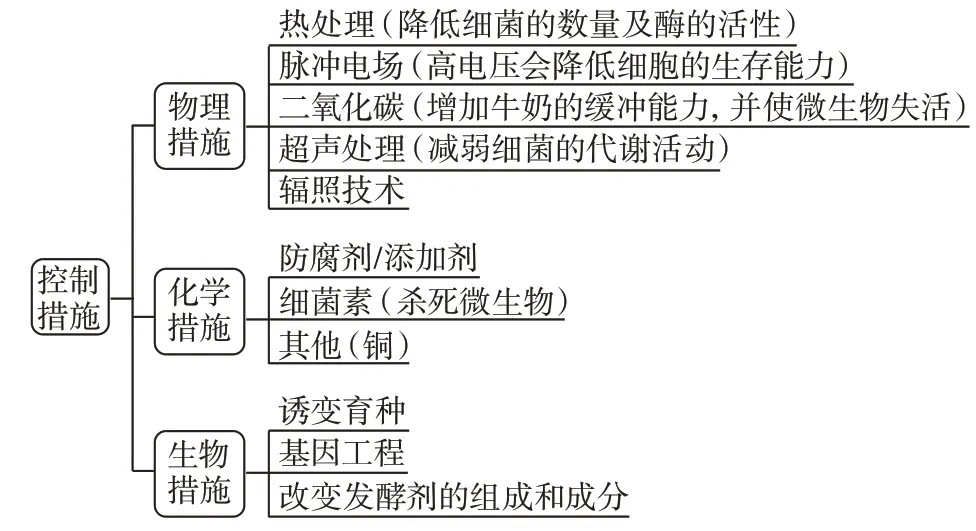
图1 适用于发酵乳制品的控制措施Fig.1 Control measures applicable to fermented dairy products
3.1 物理措施
3.1.1 热处理
热处理是在乳制品加工过程中使用的一种常见方法,通过高温短时间的加热,可以有效杀死存在于发酵乳中的有害菌。高温也会对乳酸菌产生一定的影响,使部分乳酸菌失去活性,从而防止酸度的持续增加,又可延长发酵乳的货架期,在控制发酵乳后酸化的发生中得到广泛应用,但是热处理的时间和温度必须根据产品的需求来使用。杨新尧等[23]在58 ℃下热处理发酵乳4 min,显著地改善发酵乳在冷链缺失状况下的后酸化,降低发酵乳在贮藏过程中的后酸化水平。吴彤等[24]以不同的热处理条件对凝固型酸乳进行处理,酸乳温度从70 ℃升高到95 ℃可以降低贮存24 h 和21 d时的pH 值,从而降低酸乳后酸化的水平。有些研究通过对益生菌进行轻度热胁迫处理来控制菌株带来的后酸化,通过短时间的高温处理,可以降低益生菌的活性,减少其在酸乳中的代谢活动,从而降低后酸化的风险。Zhang 等[25]研究发现轻度热胁迫处理(46 ℃,1 h)可通过降低Lactobacillusrhamnosushsryfm 1301 的代谢和增殖来限制其引起的后酸化。虽然,经热处理的发酵乳保存良好,也避免了后酸化,但受限于消费者对产品中有益微生物活性的需求。因此,采用热处理来控制后酸化需要进一步的探索。
3.1.2 脉冲电场
脉冲电场(pulsed electric fields,PEF)是一种新型的非热食品杀菌技术,以较高的电场强度(10~50 kV/cm)作为电脉冲通过食物传导到微生物细胞膜上,引起电穿孔,在一定强度下造成不可逆的细胞膜损伤,导致细胞内物质泄漏和细胞死亡[26]。尽管PEF 的发展潜力很大,但仅限于没有任何气泡的液体食品,且取决于PEF 参数、培养基组成和细胞类型、食物特性(pH 值、离子强度、抗菌性)[27]。用S.thermophilusDIL 5218 和L.delbrueckiisubsp.bulgaricusDSMZ 20081 酸化的重组脱脂乳显示,与未经PEF 处理的样品相比,PEF 处理的样品酸化速度加快12 min,氧化还原电位急剧下降。PEF 样品中发酵速率的提高是由于培养细胞的电渗透作用,从而增加了营养物质的吸收,缩短了延滞期[27]。Stühmeier-Niehe 等[28]的研究表明经PEF 处理的培养物比未经PEF 处理的培养物的发酵延滞期平均缩短22 min,对发酵剂进行PEF 预处理,通过降低pH 值滞后期导致酸化的显著加速。综合以上研究,为了在发酵乳生产中成功应用PEF,通过优化操作参数和试验方案,在确保控制发酵乳后酸化的同时,最大程度地发挥益生效果,提高产品质量和生理功效。
3.1.3 二氧化碳(CO2)处理
CO2具有可溶于水并与水反应生成碳酸的性质,从而抑制一些不耐酸微生物的生长,达到防腐、延长货架期的作用。研究表明,向液态乳中充入适量CO2,可延长其保质期:1)与普通牛奶相比,CO2牛奶在pH 4.95 和pH5.40 处有2 个缓冲的极值峰,在pH 5.10处有1 个峰值。2)高密度CO2(dense phase carbon dioxide,DPCD)以通过加压CO2或者超临界CO2的分子效应来灭活牛奶中广泛存在的细菌和真菌的营养型和内生孢子型。
在高压状态下,CO2分子溶解在微生物细胞膜的脂质双分子层中,导致细胞膜中的脂质物质被提取出来,进而增加了膜的流动性和通透性。同时,在压力作用下,CO2可以降低细胞内的pH 值,使与糖代谢相关的酶钝化,减弱乳酸菌产生乳酸的能力,进而能够实现降低发酵乳后酸化现象的目的。特别是对于发酵乳,使用CO2处理的牛奶,酸奶的保质期延长至4 个月,而未碳化的样品在30 d 内变质[29]。同样,保加利亚乳杆菌和嗜热链球菌发酵的发酵乳在CO2处理后的乳酸产量略低。
3.1.4 超声处理
超声(ultrasound,US)也是一种非加热的技术,不仅可以保留营养,还可以延长保质期和提高乳制品的品质。高强度的超声波可以提高用于发酵的发酵剂的益生菌株的活力,也可以释放酶和产酸来刺激乳糖水解,从而缩短发酵的时间[30]。Delgado 等[31]发现US 处理降低了酸奶贮藏28 d 内的后酸化,并且US 处理6 min 时,发酵乳后酸化的降低程度较显著。US 处理可以破坏β-半乳糖苷酶的活性,从而降低酸奶的后酸化程度。研究表明未经过US 处理的酸奶,酸化至pH2.0可导致酶活性快速并永久丧失。在pH4.0(接近酸奶的自然pH 值)条件下,酶的活性至少稳定60 min。对超声酸奶进行体外酸化处理,即使在pH4.0 的条件下,也会导致酶的活性快速永久损失,60 min 后仅剩20%的酶活。在较低的pH 值下培养,活性完全丧失[32]。同样,US 处理对乳酸菌代谢活动产生刺激作用,从而缩短发酵时间;对L.plantarumA3 产酸能力的抑制作用(10 min,600 W,33 kHz,20 ℃),成功实现了(后)发酵过程中的酸化调控,以缓解后酸化问题或提高工艺效率[33]。此外,与传统均质化相比,低频(20~100 kHz)高强度(10~1 000 W/cm2)超声具有投入少且易于清洁的优点。
低水平的US 处理导致β-半乳糖苷酶释放,刺激乳糖水解和半乳糖的转化,而乳糖和半乳糖的转化会增加发酵液中的可用碳源,从而加速乳酸生成速率,导致酸化加快和酸度过高,在发酵乳的后续酸化过程中不利于酸度的控制[34]。较高水平的US 处理可能造成细胞膜的物理破坏,使细胞内容物外泄。乳酸菌的大量死亡会导致发酵乳的后续酸化能力降低和失去益生效果。因此采用US 处理控制后酸化,要精确量化和控制工艺参数。
3.1.5 辐照技术
辐照技术是一种非热加工技术,γ 射线(1~10 kGy)辐照酸奶在4、20、35℃均能延长产品的货架期,且对感官性状、氨基酸含量等均无不良影响,降低过敏性[35]。在研究辐照杀菌技术对发酵乳的影响时发现,发酵乳中的乳酸菌数量与辐照剂量反比,在一定程度上降低后酸化的产生[36]。但是辐照技术在发酵乳中的应用存在一定的争议,同时有关辐照酸奶的法律在辐照食品的法律中也是非常多样化的。
3.2 化学措施
3.2.1 添加化学添加剂
通过使用商业批准的化学品限制菌株的生长来控制后酸化,从而提高发酵产品的保质期是普遍采用的做法。山梨酸和苯甲酸及其盐类对真菌和大部分细菌有抑制作用,因此常用这两类化学品进行后酸化的控制。但是苯甲酸进入人体后,能与人体内氨基乙酸化合生成马尿酸,还能与人体内的葡萄糖醛酸结合生成葡萄糖苷酸而随尿液排出体外,因此相较于山梨酸及其盐类,苯甲酸及其盐类对人体有一定的毒副作用。山梨酸钾通过抑制参与碳水化合物和柠檬酸盐利用的关键代谢酶如乳酸脱氢酶、苹果酸脱氢酶、延胡索酸酶等来抑制微生物生长。张小涛等[37]发现采用0.35%的山梨酸钾可控制山竹风味酸乳。同样,在控制红枣酸羊乳的后酸化时,发现添加0.3%的山梨酸钾可有效抑制后酸化现象[38]。
不仅山梨酸钾能够控制后酸化现象,铜在抑制发酵乳后酸化现象时也有一定的作用。Zhang 等[39]发现LDB_RS05285 是L.delbrueckiisubsp.BulgaricusATCC11842 后酸化的特异性基因,在其基因表达上调后有效抑制酸奶的后酸化。Wang 等[40]发现2.5 mg/L的Cu2+能上调LDB_RS05285 的基因表达。铜可以在不影响酸奶品质的情况下有效控制酸奶的过度酸化,改善酸奶的风味。铜还可以促进乳糖的水解,富集半乳糖和葡萄糖(从而抑制乳酸的产生),增加L-异亮氨酸、苯丙氨酸和3-甲基丁醛苯甲醛的丰度,有助于酸奶整体风味的形成。此外,发酵前向全脂牛奶中添加1.25 mg/kg 的Cu2+可有效减缓发酵乳的后酸化,并对发酵时间和发酵菌株嗜热链球菌的活菌数均无显著影响[41]。但是由于消费者对添加剂的认识不足,限制其在增强货架期和控制后酸化现象中的应用。
3.2.2 细菌素
细菌素是由细菌核糖体合成的具有抗菌活性的多肽,通常对邻近细菌具有抑制作用[42]。许多人类微生物能够产生细菌素,据报道,细菌素与人类健康密切相关,如促进肠道菌群平衡、抑制外来致病菌入侵等[43]。
细菌素作为食品添加剂在后酸化调控中具有广泛的应用前景,可以有效延长食品保质期并提供其他功能。乳酸链球菌素、片球菌素、肠球菌素和乳酸菌素是目前较为常见的细菌素类型。Hussien 等[44]研究表明2%菊糖、2%低聚果糖和嗜酸乳杆菌细菌素结合可将酸奶的保质期延长5 周以上。邱爽等[45]通过研究发现添加终浓度为28 mg/mL bifidocin A 到凝固型酸奶中可替代乳酸链球菌素,使得凝固型酸奶贮存过程中后酸化的程度得到一定的抑制。同样,研究表明产细菌素LactobacillusplantarumQ7 对保加利亚乳杆菌和嗜热链球菌的生长有一定的抑制作用,在酸奶贮藏期具有缓解后酸化的效果[46]。并且添加细菌素的样品活菌数减少,有效地解决发酵乳后酸化的问题。虽然细菌素对抑制后酸化现象有一定的效果,但是考虑到添加的细菌素可能具有细胞毒性,因此在对细菌素进行安全性评估方面应给予更多的关注。
3.3 生物措施
3.3.1 诱变育种
诱变育种是利用物理或化学方法处理菌株,使突变频率和数量增加以获得突变体,并通过采取有效的方法从突变体中筛选出所需目的菌株。诱变育种是延缓发酵乳发生后酸化的方法之一,常用的诱变方法主要有3 种:物理诱变、化学诱变和复合诱变。3 种诱变方法对发酵乳制品后酸化的影响如表1所示。

表1 3 种诱变育种对发酵乳后酸化的影响Table 1 Effects of three mutation breeding methods on post-acidification of fermented milk
物理诱变育种是通过一系列物理方法[如紫外线、微波、常压室温等离子体(atmospheric and room temperature plasma mutagenesis,ARTP)等]对菌株进行诱变,以获得具备特定形状的突变菌,并通过筛选找出符合预期目标的变异菌株。周虹瑾等[47]采用ARTP 对菌株进行诱变,用新霉素筛选出低H+-ATPase 活性的突变菌株,其中突变菌株BJT-7 经全脂牛乳发酵在20 d储藏期间几乎无明显酸度上升,并且具有较好的遗传稳定性。同样,有研究表明通过低能N+注入对保加利亚乳杆菌DL1 进行诱变后,经过青霉素筛选,获得一株后酸化能力较弱的突变菌株DL1-3。与出发菌株DL1 相比,突变菌株DL1-3 在25 ℃条件下贮藏后,后酸化程度降低18.8%,且在42 ℃条件下发酵脱脂乳的产酸能力差异较小,并可稳定遗传[49]。
化学诱变育种是通过使用一定剂量的诱变剂(如化学试剂或者是化学药物),干扰DNA 复制或者损伤DNA,来引起生长良好的菌株发生突变,从而改变其生长及其发酵特征。Wang 等[53]从新霉素诱变的德氏乳杆菌保加利亚亚种成功分离到2 株H+-ATPase 缺陷变种,突变菌株的H+-ATPase 活性分别降低了51.3% 和34.3%,其生长情况以及产酸能力较原始菌株明显减弱。将突变菌株接种于发酵乳中,结果表明发酵乳在贮藏过程中的后酸化现象被显著抑制。张祥等[54]利用硫酸新霉素诱变L.plantarumZUST 菌株,筛选出两株低H+-ATPase 活性植物乳杆菌,从而使突变菌的产酸能力下降,对缓解后酸化现象有一定的作用。同样,使用硫酸新霉素诱变的L.delbrueckiissp.BulgaricusND06的突变菌株ND06-2 具有较低的H+-ATPase 活性,其在发酵或者储存过程中产生乳酸的能力明显低于原始菌株[50]。杨玲等[55]也是用硫酸新霉素诱变,获得1 株弱后酸化LactobacillushelveticusL551-1。与原始菌株比,L551-1 延迟了发酵过程中的产酸时间,同时使得发酵结束时的酸度更容易控制,L551-1 发酵乳货架期后酸化问题得到显著改善。
复合诱变是将两种以上的方法进行结合,相较于单一诱变的方法,复合诱变可以弥补单一诱变的缺陷。李学丛[52]对LactobacilluscaseiN1115 进行紫外诱变和硫酸二乙酯诱变,采用新霉素进行筛选,诱变菌株C1可延长发酵乳冷藏期间保质期5 d 左右,并且能很好地改善原菌株N1115 的感官品质。诱变育种虽然能在短时间内得到具有弱后酸化的突变菌株,但是在突变过程中容易出现突变性状不稳定,甚至出现突变逆转到原始状态的情况。
诱变育种方法可以改变菌株的性状,延缓发酵乳的后酸化。通过增加突变频率和数量,可以获得更适合生产需求的菌株,以提高产品的质量和稳定性。但需要注意的是,在诱变过程中,需要对处理条件进行仔细控制,以确保获得安全和有效的突变体。
3.3.2 改变发酵剂比例和组成
发酵剂是发酵乳发酵的基础,其在发酵完成后同样可代谢产生乳酸,造成后酸化现象。为了降低发酵乳的后酸化现象,研究者们采用改变发酵剂组成的策略,即从改变发酵剂的组成和比例两个方向入手,来抑制发酵乳制品的后酸化现象。Settachaimongkon 等[56]对L.plantarumWCFS1 在亚致死胁迫(NaCl 升高和pH值较低)条件下进行预培养,将亚致死预培养的LPWCFS1 与传统酸奶发酵剂共培养,发现添加亚致死预培养的LP-WCFS1 显著抑制了保加利亚乳杆菌亚种的存活,从而减少了冷藏期间酸奶的后酸化。杜昭平等[57]的研究表明在发酵乳发酵过程中适当提高球杆菌比例,有利于减弱发酵乳后酸化的现象。综上,改变发酵剂的比例和组成能在一定程度上降低发酵乳制品的后酸化现象。
3.3.3 基因工程
基因工程是在基因水平上对发酵剂进行改造,通过这种方法来达到降低发酵乳后酸化现象的目的。基因工程经常被应用于所需功能的乳酸菌的生物技术升级。与诱变育种相比,基因工程能够跨越难以突破的物种界限,有目的地改变乳酸菌基因特征,以获取具有稳定遗传特性的突变菌株。李晨等[58]使用带有ISS1的载体pGhost9:ISS1 引入到保加利亚乳杆菌中,经复制型转座后获得了3 株抗后酸化的突变菌株,并且得到的菌株突变性状稳定。Zhang 等[59]评估了69 个后酸化候选基因与不同保加利亚乳杆菌发酵酸奶酸度变化的相关性,从而筛选出用于后酸化的生物标志物LDB_RS00370,并通过此标记物筛选发现100 mg/kg的烟酸可抑制发酵乳的后酸化。但是基因操作可能会使菌种间的划分界限变得模糊,使得对发酵剂中菌种的鉴定不精确。因此,基因工程技术在抑制发酵乳制品后酸化现象中的应用还需要进一步的探究。
4 总结与展望
发酵乳因其独特的口感和较高的营养价值备受消费者的青睐,但后酸化现象破坏了保质期内发酵乳的品质,制约了发酵乳的销售和产业发展。本文阐述了发酵乳后酸化与发酵剂类型、乳的成分、温度和pH值、均质和搅拌、益生菌和益生元等因素有关,并讨论了其后酸化的产生机制以及控制措施。在不影响发酵乳的流变、感官性状以及货架期的情况下,本文中讨论的各种策略可以单独或者联合应用,来防止后酸化的现象。但是,每种方法都有自己的优缺点。发酵乳的热处理虽然延长了发酵乳的货架期,减少了后酸化,但由于失去了不耐热的生物活性肽和有益微生物,不适合作为功能性发酵产品。研究发现,CO2添加有助于延长发酵乳制品的货架期和延缓酸的形成,但其对感官特性和后酸化的平行影响尚未见报道。新兴的非热技术(超声、辐照等)需要进一步探索其对发酵乳或者人体是否有不利影响以及制定更加完整、有效的监管审批。细菌素在酸奶中的直接掺入因其高昂的纯化成本和在食品基质中的稳定性而受到阻碍。添加防腐剂比添加细菌素的成本更加低廉,但是必须通过安全评价的验证。进行诱变育种虽然是缓解后酸化现象的一个途径,但是一定要经过严格的筛选,同时还要防止突变的菌株出现性状的退化或者是逆转到原来性状。
未来有望通过以下方法来缓解后酸化:1)通过诱导成分变化来增强缓冲能力,即通过膜过滤来调整牛奶中的蛋白质/乳糖比例,从而开发用于制造温和风味发酵产品的牛奶。2)使用包裹GRAS 抗菌剂的pH 值敏感凝胶,以便在达到所需pH 值后允许抗菌药物的持续释放。3)探究允许的/天然添加剂对后酸化的影响,同时结合经济有效的加工方法进行尝试。
总之,对于所讨论的技术需要进一步的努力,以开发缓解后酸化现象且不影响其理化性状的产品,并将其转化为工业化生产。希望该文能够为未来制定新的成本低且效益高的策略来缓解发酵乳后酸化的相关工作提供参考。
猜你喜欢
中老年保健(2022年3期)2022-08-24 02:59:30
中国食物与营养(2021年11期)2021-12-21 06:34:58
中国土壤与肥料(2021年5期)2021-12-02 01:05:40
中国生殖健康(2020年8期)2021-01-18 03:05:44
中国生殖健康(2018年3期)2018-11-06 07:20:20
启蒙(3-7岁)(2018年7期)2018-07-10 09:38:16
山东工业技术(2016年15期)2016-12-01 05:30:45
发明与创新(2015年21期)2015-02-27 10:39:11
水土保持通报(2014年5期)2014-06-09 08:27:10
中国酿造(2014年9期)2014-03-11 20:21:04
