
松褐天牛中脊茧蜂生物学特性研究
2024-02-26 05:40王少博韩孟娇王小艺杨忠岐
昆虫学报 2024年1期
王少博, 韩孟娇, 魏 可, 王小艺, 杨忠岐
(中国林业科学研究院森林生态环境与自然保护研究所, 国家林业和草原局森林保护学重点实验室, 北京 100091)
中国作为目前遭受松材线虫病(pine wilt disease)危害最为严重的国家,在传入40余年的时间内,我国数亿株松树因感染松材线虫Bursaphelenchusxylophilus而死亡,造成的经济损失高达上千亿元。根据国家林业和草原局最新公告(2023年第7号),当前全国共有19个省(自治区、直辖市)701个县级行政区发生松材线虫病疫情,疫情发生态势十分严峻,并面临着“北扩西进”持续扩散的态势。我国6 000万hm2松林时刻面临着松材线虫病侵袭的风险,尽快遏制松材线虫病的传播蔓延是当前主要防控目标(叶建仁, 2019)。媒介昆虫作为松材线虫完成扩散的必须环节,只要能切断这个环节就能阻止松材线虫病的传播,是目前防治松材线虫病的主要措施之一(张彦龙, 2012; 高尚坤, 2016)。因为媒介昆虫一旦羽化出孔,便具备了随时传播松材线虫的可能,因此在媒介昆虫未成熟期将其灭杀是阻断传播媒介的关键(杨忠岐, 2004; 杨忠岐等, 2018)。
松褐天牛Monochamusalternatus是当前中国乃至整个亚洲地区松材线虫病最主要的传播媒介之一(Kobayashietal., 1984; Mota and Vieira, 2008; Futai, 2013)。由于其整个幼虫阶段均隐蔽在树体内钻蛀危害,常规的防治措施难以接触到虫体发挥作用(张彦龙等, 2022)。天敌昆虫在与寄主害虫的长期协同进化过程中,形成了特有的寄主搜索机制,总是能够快速地找到寄主并发挥控制作用(王小艺和杨忠岐, 2008),是目前能够有效利用防治蛀干类害虫的最佳途径之一。当前,花绒寄甲Dastarcushelophoroides和肿腿蜂Sclerodermusspp.被广泛应用于松褐天牛的防治,均取得了一定的防控成效(来燕学等, 2012; 杨远亮等, 2013; 展茂魁等, 2014; 魏可等, 2015; 温小遂等, 2017),但同时也暴露出一些技术瓶颈。例如,花绒寄甲主要寄生松褐天牛老熟幼虫和蛹;而肿腿蜂类寄生蜂寄生谱较宽、扩散能力较弱,主要寄生3龄以下松褐天牛幼虫,在防治应用过程中对松褐天牛的控制作用有限。目前,在松褐天牛幼虫期尚无专化性较强的天敌,且也没有应用于防治松褐天牛中老龄幼虫的天敌昆虫。
此前,本研究室在贵州遵义和湖北恩施调查松材线虫病时,发现了一种寄生3-5龄松褐天牛幼虫的茧蜂。该蜂野外寄生率最高可达30%,在其生境内尚未发现寄生其他昆虫,在1头松褐天牛身上最多发现15头寄生蜂幼虫。基于以上特点,推测该蜂是一种极具防治潜力与应用价值的松褐天牛幼虫期天敌。经鉴定,该寄生蜂为前期报道的一种寄生松褐天牛幼虫的茧蜂——Bracomorphaninghais(Wangetal., 2009)。2021年,Bracomorpha被划入中脊茧蜂属Cyanopterus的亚属(Lietal., 2021),该寄生蜂的学名遂更改为Cyanopterusninghais,其中文名暂定为松褐天牛中脊茧蜂。目前,关于该寄生蜂的研究除成虫形态学描述外(Wangetal., 2009),无其他相关结果。为充分了解该寄生蜂的生防潜能,更好地开发利用于松褐天牛的生物防治,本研究对该寄生蜂的基础生物学特性进行系统实验观测,以期为该寄生蜂的人工大规模繁育和野外应用提供理论基础与技术指导。
1 材料与方法
1.1 供试昆虫
松褐天牛中脊茧蜂建群蜂于2022年3月采自贵州省遵义市龙坑镇共青湖(27°35′51.87″N,106°50′21.07″E)感染松材线虫病的马尾松Pinusmassoniana上。松褐天牛卵或1龄幼虫采集自上述松林内,并在实验室内用人工饲料饲养繁殖,作为繁蜂寄主。在室内用3-4龄松褐天牛幼虫作为寄主繁育松褐天牛中脊茧蜂,获得试验种群。
繁殖寄生蜂带寄主木段的制作。将直径3~4 cm粗的新鲜马尾松木段截成20 cm长。用刀沿木段纵向将树皮划成“U”型切口,从U型开口端撬开树皮但不割断,沿“U”型切口边缘在木质部上切开一个凹槽(长3.5 cm,宽1 cm,深1 cm)。将1头松褐天牛幼虫放置于槽内,合上U型树皮,用PARAFILM封口膜封上切口上端,木段两端用石蜡封口防止失水。每根木段上制作6~8个凹槽,接入同等数量的松褐天牛幼虫。
根据前期预试验结果,松褐天牛中脊茧蜂通常在羽化3-5 d后才会发生寄生行为,雌成蜂每天通常只进行1次寄生行为,单次寄生持续时间为2~3 h。 本研究中所有试验处理均在实验室温度(23±1) ℃和相对湿度50%±5%条件下开展。
1.2 羽化节律测定
将150头已交配但无寄生经历的7-14日龄松褐天牛中脊茧蜂雌成蜂平均接入6个养虫笼(长×高×宽=25 cm×25 cm×25 cm)中,笼内置有带寄主的木段,蜂虫比为1∶1。寄生24 h后移除雌蜂。待子代开始羽化时,每2 h(6:00-22:00)记录1次雌雄子代在各时段的羽化数量及日羽化节律,直至羽化结束。
1.3 交配行为观测
待松褐天牛中脊茧蜂结茧后收集茧蛹并单头存放于玻璃指形管内,待其羽化后,将50头尚未交配的1日龄成蜂按照雌雄1∶1的比例接入养虫笼内,观察和记录交配(求偶、交配前期、交尾与交配后期)全过程及日节律,累计观察发生交配行为的寄生蜂20对。将上述已交配过的成蜂分雌雄转入新的养虫笼中,在笼中引入同等数量尚未交配的1日龄异性成蜂,观察已交配过的成蜂是否存在二次交配行为。将1日龄尚未交配的雌雄成蜂以1∶3和3∶1的比例分别置于养虫笼中,每笼成蜂数量均为40头,观察记录是否存在交配竞争现象。
1.4 寄生行为观测
将已交配但无寄生经历的7-14日龄松褐天牛中脊茧蜂雌成蜂接入养虫笼中,笼内置有带寄主的木段,蜂虫比为2∶1。观察记录雌蜂对寄主松褐天牛的寄生全过程(搜寻、检查、产卵与梳理),共观察有发生寄生行为的雌蜂20头。
1.5 生殖方式及雌蜂交配与否对寄生的影响测定
已交配雌蜂:将1头已交配但无寄生经历的7-14日龄松褐天牛中脊茧蜂雌成蜂接入养虫罐(底径8 cm,高22 cm)中,罐中置有带寄主的木段,蜂虫比为1∶1。24 h后移除雌蜂,继续培养至子代羽化。未交配雌蜂:将上述处理中已交配的雌蜂替换为未交配的雌蜂,其余处理保持一致。2组处理均重复20次。
待子蜂羽化后统计雌蜂在不同交配状态下所产子代性比,并解剖带虫木段统计寄生率,记录单头寄主所育寄生蜂子代的数量。生殖方式:若交配后的雌蜂所产子代可发育成新个体,则认为该寄生蜂可进行两性生殖;若未交配的雌蜂所产子代也可发育成新个体,则认为该寄生蜂同时可进行孤雌生殖。
1.6 寄主龄期偏好性测定
非选择性测定:每根马尾松木段上分别接入1头松褐天牛1-5龄幼虫或蛹,将单根带虫木段与1头已交配但无寄生经历的7-14日龄松褐天牛中脊茧蜂雌成蜂置于同一养虫罐(底径8 cm,高22 cm)中,24 h后移除雌蜂,继续培养至子代蜂羽化,解剖木段统计雌蜂对不同龄期寄主的寄生率及单头寄主所育的寄生蜂子代数量。每个龄期的松褐天牛测试20头,即接种20根木段。
选择性测定:在1根马尾松木段上接入松褐天牛1-5龄幼虫和蛹各1头,共计6头。随后,将1头已交配但无寄生经历的7-14日龄松褐天牛中脊茧蜂雌蜂与1根带寄主木段置于养虫罐中,24 h后移除雌蜂,继续培养至子代羽化,解剖木段统计雌蜂对寄主的选择情况及单头寄主所育的寄生蜂子代数量。共测试雌蜂20头。
1.7 成蜂寿命测定
待松褐天牛中脊茧蜂结茧后,收集茧蛹并单头存放于玻璃指形管内,每隔12 h检查1次是否有成蜂羽化。待寄生蜂羽化后,收集羽化12 h内未交配的成蜂,分雌雄单头置于养虫盒中(底径6 cm,高4 cm)中。盒内分别提供浸有20%蜂蜜水、纯水和无任何物质(对照)的脱脂棉棒补充营养,每48 h更换一次棉棒。记录不同营养源条件下成蜂的寿命,直到所有供试个体死亡。每个处理测试雌雄成蜂各20头,共计120头。
1.8 数据分析
数据分析采用EXCEL 2019计算平均值与标准误,利用SPSS 26软件进行数据处理,交配或未交配松褐天牛中脊茧蜂雌成蜂对松褐天牛幼虫的寄生率,以及雌蜂对不同龄期寄主寄生率的差异采用卡方检验法分析。子代蜂数量及成蜂寿命的差异采用单因素方差分析(one-way ANOVA)和Duncan氏新复极差法或独立样本t检验进行分析。
2 结果
2.1 成蜂羽化节律
共收集到松褐天牛中脊茧蜂茧蛹368头,羽化出359头成蜂,其中包括雌蜂168头,雄蜂191头,羽化率达97.55%。羽化共持续4 d,雄蜂比雌蜂羽化平均早0.81 d。雄蜂在第2天达到羽化高峰,占雄性羽化总数的34.55%;雌蜂在第3天达到羽化高峰,占雌性羽化总数的37.50%(图1: A)。雌雄成蜂日羽化节律相似,均在上午8:00-10:00达到羽化高峰,上午6:00-12:00羽化成蜂数占总羽化成蜂数的61.00%;其次是下午12:00-18:00,占比33.15%;晚间18:00-22:00羽化数量最低,占比5.85%(图1: B)。
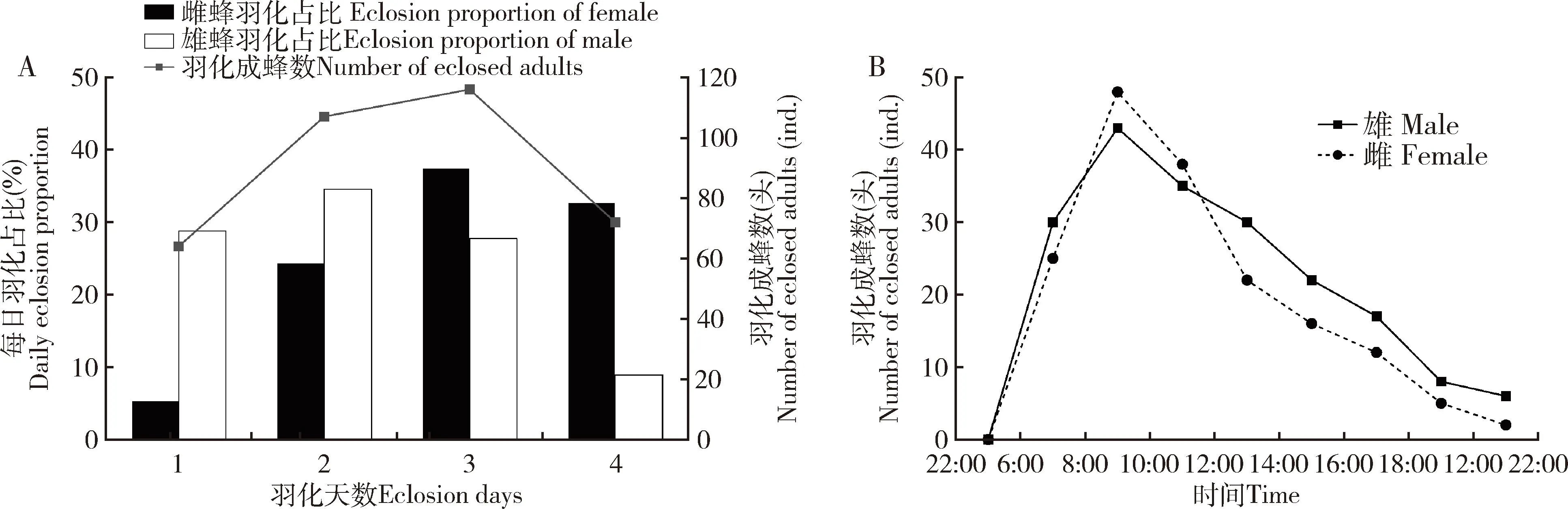
图1 松褐天牛中脊茧蜂不同羽化天数(A)与不同羽化时间段(B)的羽化节律
2.2 松褐天牛中脊茧蜂成蜂的交配行为
松褐天牛中脊茧蜂成蜂交配行为可划分为求偶、交配前期、交尾与交配后期4个阶段。(1)求偶:雄蜂飞至雌蜂身后,连续快速振动翅,爬行追逐雌蜂(图2: A);(2)交配前期:雄蜂追上雌蜂后,雄蜂爬上雌蜂背部,试图用前足控制住雌蜂(图2: B);用触角不断敲击雌蜂触角的同时,腹部向下弯曲尝试与雌蜂生殖器完成对接,试探雌蜂是否接受交配(图2: C);雌蜂或同意完成交配,或迅速向前爬行逃避拒绝此次交配。交配前期持续时间较短,约2~5 s内完成;(3)交尾:雌蜂接受交配后,雄蜂后退同时腹部向下弯曲完成与雌蜂生殖器对接,触角有规律的上下摆动。整个交尾过程,雄蜂翅保持竖立;雌蜂或静止不动,由雄蜂用足抱握;或不断向前爬行,此时雄蜂整个身体后仰,仅通过交配器与雌蜂相连,任由雌蜂拖动前行,整个交尾过程可持续120~180 s(图2: D);(4)交配后期:交配完成后,雌雄蜂分离,静止休息。雌蜂面对其他雄蜂寻求交配时迅速逃避,拒绝交配。雄蜂休息片刻后,用口器和前足梳理翅与触角,为再次交配做准备。

图2 松褐天牛中脊茧蜂成蜂的交配行为
松褐天牛中脊茧蜂成蜂羽化后即可进行交配。成蜂从下午14:00开始逐渐进入交配活跃期,交配高峰期集中发生在17:00-21:00,占整个交配观察次数的65%; 22:00至次日整个上午,该蜂交配行为不够活跃,常保持静止状态。雌蜂在交配时对配偶具有明显的选择性,常出现在交配前期,雄蜂剧烈振翅寻求交配,而雌蜂一直逃避造成交配失败,雄蜂首次求偶并交配成功的概率仅为40%。雌蜂一生只交配1次,雄蜂可多次交配,交配过的雌蜂再次面对雄蜂求偶行为时会表现出明显的逃避行为,而交配过的雄蜂在获得短暂的休息后即可进行二次交配。雌雄蜂比例为1∶3时,会出现交配局部配偶竞争现象,表现为雌雄蜂交配过程中多头雄蜂同时竞争进行交配,导致交配中断;雌雄蜂比例为3∶1时,直至雄蜂全部死亡,未发现任何一例多头雌蜂追逐1头雄蜂,也未发现未在交配状态的雌蜂干扰正在交配雌雄蜂的现象。
2.3 寄生行为
松褐天牛中脊茧蜂整个寄生过程可分为以下4个阶段:(1)寄主搜寻:当雌蜂搜寻到含寄主的带虫木段时,雌蜂会在木段上边迅速移动边用左右运动的触角不断横扫敲击树皮,直至找到寄主所存在的位置(图3: A);(2)寄主检查:雌蜂在找到寄主位置后,并不会立即产卵,而是先对寄主进行仔细的检查。雌蜂会利用产卵器尝试刺探感受可刺入的孔隙,不断地调整刺入的位置,以完成产卵器与寄主相接触,判定该寄主是否适合寄生,做出产卵或放弃继续搜寻的决定,直到找到适合寄生的寄主(图3: B);(3)产卵:找到合适的寄主后,雌蜂先利用后足保持身体稳定,然后腹部向下弯曲伸出产卵器开始蜇刺攻击寄主。在此过程中,雌蜂会先分泌毒液麻醉或杀死寄主,而后再进行排卵活动,整个产卵过程可持续2-3 h(图3: C);(4)结束后梳理:产卵结束后,雌蜂会缓慢的将产卵器抽出,利用前足和口器对触角和产卵器进行梳理,并用后足整理翅(图3: D)。梳理完毕后,会在产卵地点附近休息一段时间,为下次寄生做准备。
2.4 生殖方式及雌蜂交配与否对寄生的影响
松褐天牛中脊茧蜂雌蜂无论交配与否,均可完成寄生并产生子代,交配后的雌蜂所产子代性比为1∶1,未交配的雌蜂所产子代均发育成雄性,结果表明松褐天牛中脊茧蜂营两性生殖和孤雌产雄生殖(表1)。该蜂在寄生过程中会将自身所产生的毒液注射到寄主体内,使寄主永久麻醉或被杀死,以保证寄生蜂子代幼虫孵化后就能够在无防御抵抗能力的寄主上取食发育,试验过程中几乎所有被寄生的寄主均在寄生蜂幼虫孵化前(1-2 d内)即丧失运动能力甚至死亡。寄生蜂幼虫孵化后群集分布在寄主体表,吸食寄主血淋巴完成发育(图4)。该蜂属典型的抑性-外寄生蜂。

表1 松褐天牛中脊茧蜂不同交配状态下的寄生率、单寄主育雏数及子代雄性占比
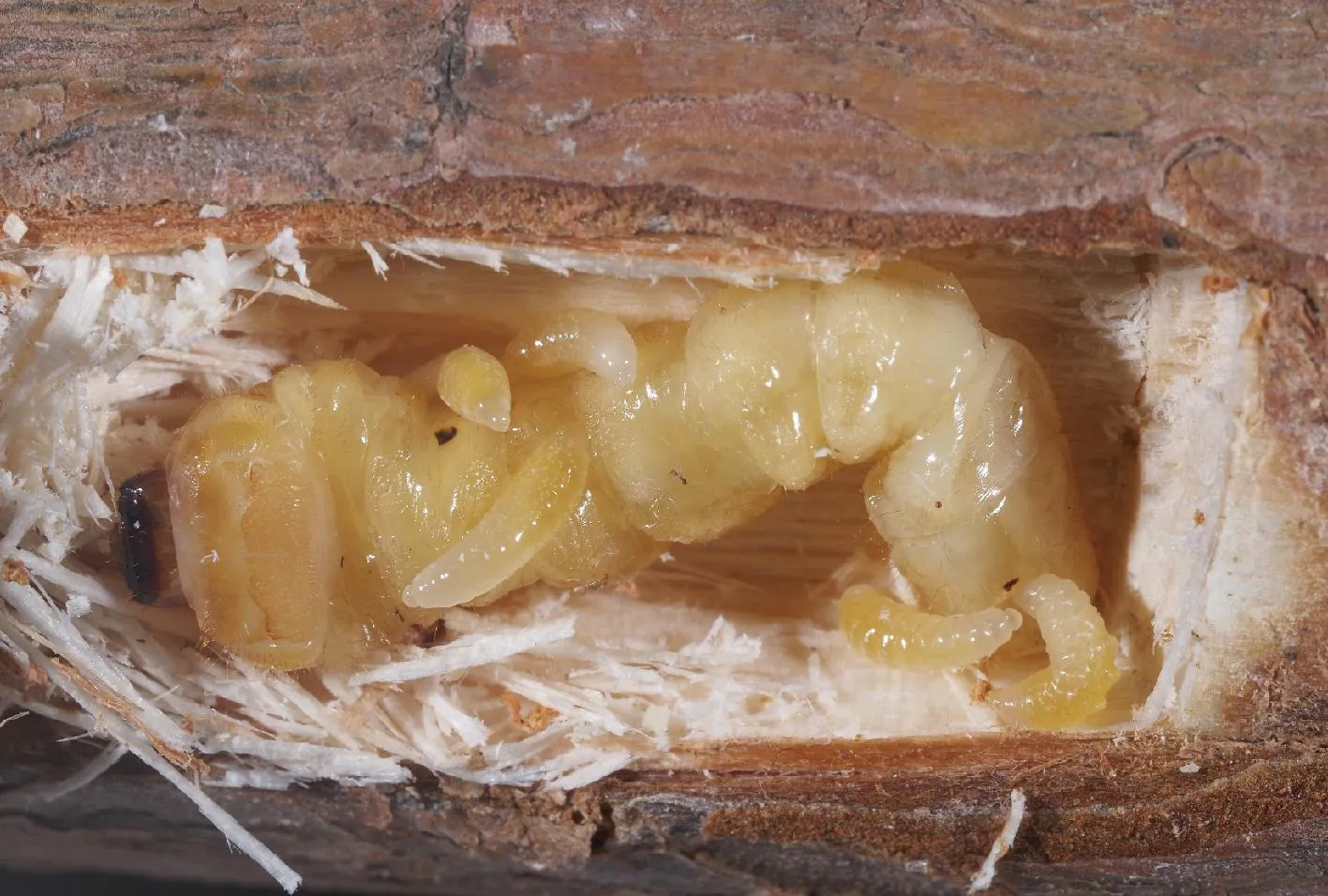
图4 松褐天牛幼虫上松褐天牛中脊茧蜂幼虫寄生状
在24 h寄生时间内,已交配的雌成蜂对寄主松褐天牛幼虫的寄生率达85%,未交配雌蜂对寄主的寄生率为75%,雌蜂交配与否对寄主的寄生率无显著差异(χ2=0.625,df=1,P=0.429),单头寄主上所产子代数量差异也不显著(t=0.648,df=30,P=0.522)。交配后的雌蜂所产子代中雌性45头,雄性47头,在单头松褐天牛幼虫上所产的子代蜂数为3~11头,其中雌蜂平均(2.64±0.33)头,雄蜂平均(2.76±0.38)头,子代雌雄性比在0.6~2.0∶1之间。未交配的雌成蜂所产子代均为雄性,单头松褐天牛幼虫上所产子代蜂数为4~10头,平均每头松褐天牛幼虫上可育出寄生蜂雄性子代(5.93±0.50)头(表1)。
同列数据后相同小写字母表示经t检验差异不显著(P>0.05)。The same lowercase letters following the data in the same column indicate no significant difference byt-test (P>0.05).
2.5 对不同龄期寄主的寄生偏好性
在非选择性测试中,松褐天牛中脊茧蜂雌成蜂可成功寄生2-5龄松褐天牛幼虫,松褐天牛1龄幼虫和蛹不能被寄生。其中,对松褐天牛3龄幼虫的寄生率最高,为85%;其次是对4龄幼虫,寄生率为65%;对5龄与2龄幼虫的寄生率相对较低,分别为30%与20%。该蜂对2-5龄松褐天牛的寄生率差异显著(χ2=22.000,df=3,P<0.00 1)。5龄松褐天牛幼虫所育寄生蜂子代数量最多,平均为(7.83 ± 0.79)头;4龄与3龄幼虫所育子代蜂数量次之,分别为(4.77±0.50)和(3.35±0.33)头;2龄幼虫所育子代蜂数量最少,仅为(1.25±0.25)头。不同龄期松褐天牛所育子代蜂数量差异显著(F=17.801,df=3, 36,P<0.001)。
在选择性测试中,该蜂仅寄生3龄和4龄松褐天牛幼虫,但对3龄幼虫的寄生率显著高于对4龄幼虫的寄生率(χ2=4.900,df=1,P=0.027),而3龄与4龄松褐天牛幼虫所育出的子代蜂数量差异不显著(t=0.988,df=18,P=0.336)(表2)。
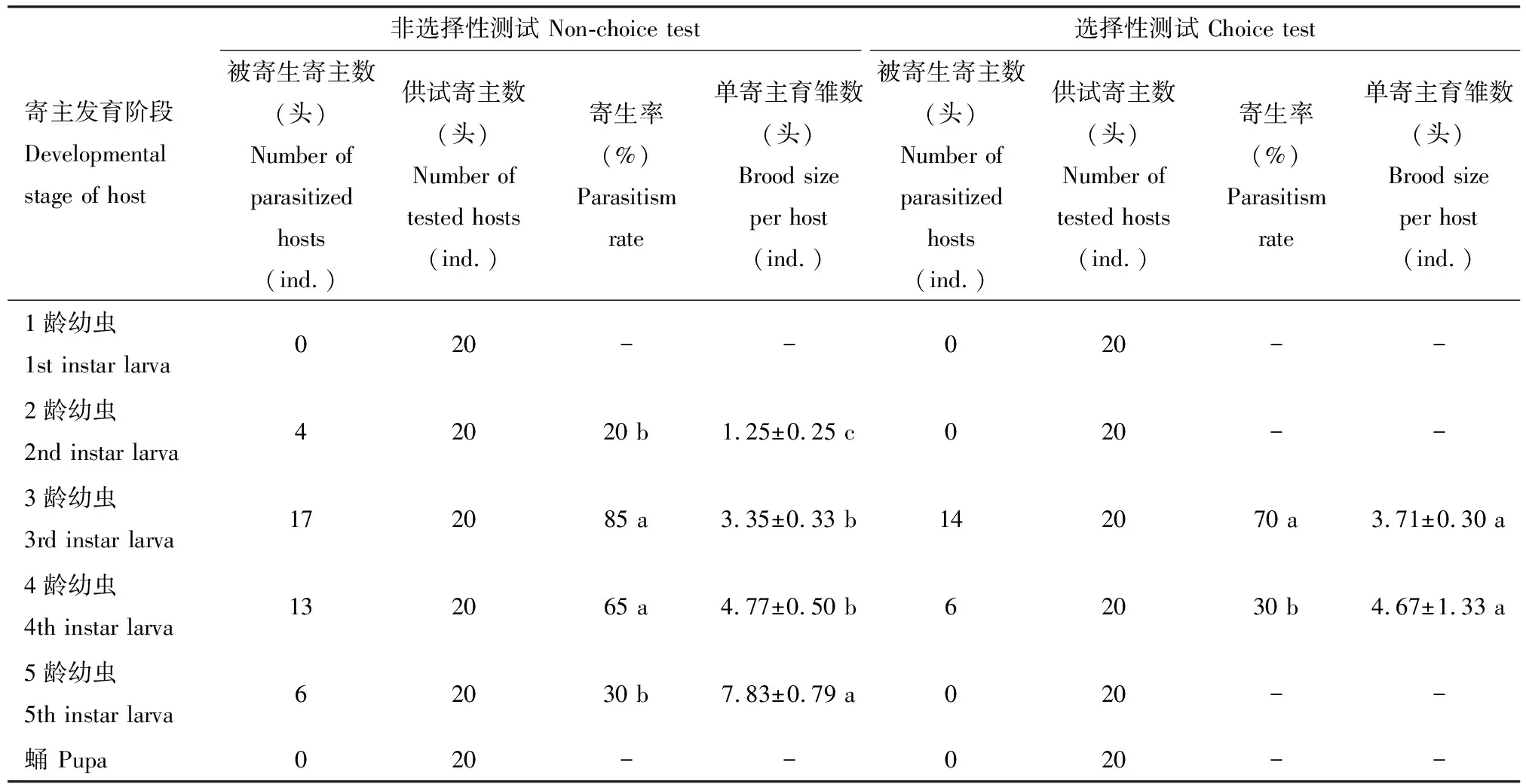
表2 松褐天牛中脊茧蜂雌成蜂对不同龄期寄主松褐天牛幼虫和蛹的寄生选择
2.6 不同营养条件下的成蜂寿命
补充营养能够明显延长松褐天牛中脊茧蜂成蜂寿命,不同营养源下雌雄蜂寿命存在显著差异。在无任何物质喂食情况(对照)下,松褐天牛中脊茧蜂雌雄成蜂寿命分别为(7.00±0.70)和(7.20±0.76) d,提供纯水与20%蜂蜜水均能在不同程度上延长雌雄成蜂寿命。雄蜂在20%蜂蜜水、纯水与无喂食处理下的寿命均存在显著差异(F=19.83,df=2, 57,P<0.001),20%蜂蜜水条件下平均寿命为(20.80±2.00) d,最长可存活40 d;纯水喂养条件下的寿命为(12.40±1.60) d,生存曲线如图5(A)。雌蜂提供20%蜂蜜水平均寿命为(35.50±3.31) d,最长可存活64 d,显著高于纯水和对照处理下的寿命(F=61.452,df=2, 57,P<0.001);提供纯水时雌蜂平均寿命为(9.00±0.95) d,与对照差异不显著(P>0.05),生存曲线如图5(B)。
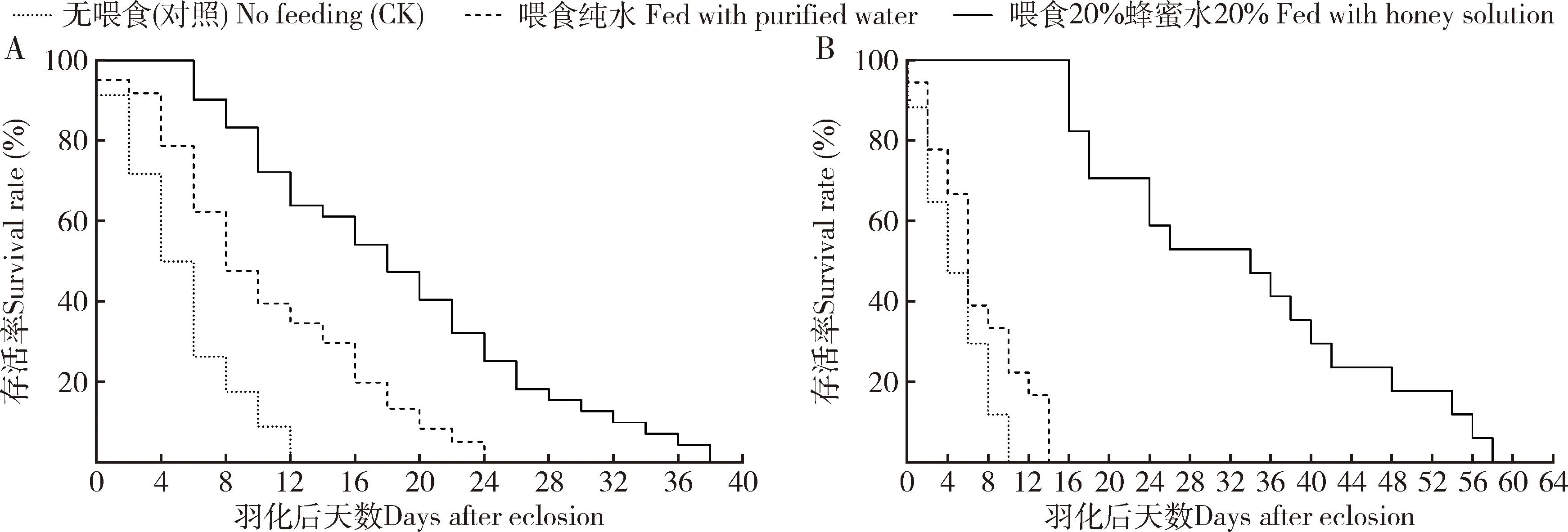
图5 不同营养条件下松褐天牛中脊茧蜂雄(A)和雌(B)成蜂寿命
3 讨论
松褐天牛中脊茧蜂成蜂的羽化行为表现出很强的昼夜节律性。羽化主要集中在白天,高峰期为6:00-12:00,雌雄成蜂在羽化期内保持着几乎相同的羽化节律,这与褐带卷蛾茧蜂Braconadoxophyesi(罗旭初等, 2017)、绿眼赛茧蜂Zelechlorophthalmus寄生率和单寄主育雏数数据后的不同小写字母分别表示经卡方检验和Duncan氏新复极差多重检验差异显著(P<0.05)。Different lowercase letters following the parasitism rate and the brood size per host indicate significant differences based onchi-square test and Duncan’s new range multiple test (P<0.05).
(李倩等, 2017)、黄色潜蝇茧蜂Opiusflavus(杨琪, 2017)等茧蜂科的昆虫羽化节律类似,这可能与寄生蜂长期进化所形成的昼夜节律相关,许多膜翅目昆虫羽化过程中也同样表现出这一规律。
在实验中我们发现,松褐天牛中脊茧蜂雌成蜂总会在交配前做出伸展翅、翘起腹部的动作,随后雄蜂好像受到某种物质的吸引,准确定位到雌蜂的位置,并在靠近雌蜂身后时开展求偶行为,这其中就包括翅的快速振动以及与雌蜂触角相互触碰,而后由雌蜂决定是否接受交配。在寄生蜂交配过程中,雌蜂伸展翅、翘起腹部这一行为被认为是释放性信息素的信号(Dancietal., 2010)。此前,化学信号已被证实在棉铃虫齿唇姬蜂Campoletischlorideae的两性识别和选择过程中发挥重要作用(Guoetal., 2022),雌蜂释放的长距离信息素会引起雄蜂逆风飞行搜寻异性(Villagraetal., 2011),而短距离的信息素会刺激雄蜂的求偶行为(McClureetal., 2007)。振翅和触角触碰作为雄蜂求爱的始发信号,寄生蜂交配过程中振翅的频率和振幅已被证实对于交配的成功发挥着决定性的作用(Kimani and Overholt, 1995; Avilaetal., 2017),这可能与雌蜂抉择雄蜂的质量和状态有关(Daretal., 2021)。在阿尔蚜茧蜂Aphidiuservi(Villagraetal., 2011)与茶足柄瘤蚜茧蜂Lysiphlebustestaceipes(Benellietal., 2016)等寄生蜂中都证实了这种现象对于交配成功的重要性。另外,交配过程中雌雄蜂触角的相互触碰被证实与触角腺分泌物相关,雄蜂利用触角表面的微孔释放化学信息素,对于两性间的相互识别和交配发挥着举足轻重的作用(Battagliaetal., 2002)。
对于寄生隐蔽性寄主的寄生蜂而言,能够发现并接受寄主是成功寄生的关键,在寄生过程中很大程度上依赖寄生蜂在栖境中对寄主的定位能力(Hausmannetal., 2005)。寄主、寄主粪便或蛀道等挥发物以及寄主取食或运动产生的振动信号是寄生蜂准确定位和选择是否寄生寄主的重要线索(Sullivan and Berisford, 2004)。此前在白蜡吉丁啮小蜂Tetrastichusplanipennisi(Ulyshenetal., 2011)、白蜡吉丁柄腹茧蜂Spathiusagrili(Wangetal., 2010)和黑腹单节螯蜂Haplogonatopusoratorius(Hirota and Mita, 2021)等寄生蜂的寄生过程中,已证实寄主植物所产生的化学挥发物与寄主的机械振动对于寄主定位的重要性。作为抑性外寄生蜂,松褐天牛中脊茧蜂搜寻到合适的寄主后,首先会通过产卵器分泌毒液以麻痹或杀死寄主,然后再进行排卵活动,几乎所有被寄生的寄主均在寄生蜂卵孵化前就已丧失活动能力甚至死亡。以往的研究表明上述特点是抑性外寄生蜂的最重要的生物学特征,雌蜂将毒液注射到寄主体内引起寄主停止发育、永久麻醉或死亡,以保证其子代发育不会受到寄主的驱赶、伤害甚至杀死的风险(Gauld, 1988; Whitfield, 2003)。
在寄主龄期偏好性测试当中,松褐天牛中脊茧蜂对3-4龄松褐天牛幼虫具有更高的选择性(表2),这不仅表明松褐天牛3-4龄幼虫是繁育该蜂最适合的龄期,同时也是野外最适合应用于防治松褐天牛的时期。而导致这种偏好性的原因可能是由于寄生蜂与寄主经过长时间协同进化。在自然界中,松褐天牛幼虫3龄及之前均在树皮下与韧皮部之间的蛀道内取食,幼虫4龄时虽逐渐开始为侵入木质部做准备,但仍有较长的时间在树皮下活动,在这一阶段中被寄生蜂搜寻寄生的概率也就相对较高;而当松褐天牛幼虫进入5龄后,就开始在木质部内蛀造蛹室为化蛹做准备,更多时间存在于较深木质部中,被成功寄生的概率也就变得很低。经过长时间的协同进化,遂产生了这种寄主龄期偏好性。在非选择性测试当中,不同龄期寄主所育子代蜂数量符合“寄主大小-质量假说”(Wangetal., 2008),雌蜂在寄生时会权衡(trade-off)出最适合的寄主去产卵,会排除掉1龄、绝大部分2龄以及部分过大的5龄寄主幼虫。子代数量会随着寄主龄期的增大逐渐增长,这是因为太小的寄主不能满足寄生蜂子代发育的营养需求,较大的寄主虽然能够满足更多数量的子代蜂发育营养需求,但通常也需要花费更多的寄生用时与更大的自身消耗(Weietal., 2014)。
补充营养能够对寄生蜂的寿命和寄生行为产生重要的影响(Harveyetal., 2010)。在自然界中,已证实寄生蜂羽化后主要通过取食蜜源植物的花蜜、蜜露等糖原物质作为能量来源,来满足其生长、发育和繁殖的需要(Benellietal., 2017)。在实验室内,蜂蜜被认为是完美的花蜜替代品,通常被用来饲养寄生蜂,其主要成分为果糖和葡萄糖,对昆虫有很高的营养价值。在本研究中,相比于纯水,补充20%的蜂蜜水能够显著延长松褐天牛中脊茧蜂成蜂寿命,且对雌成蜂寿命的影响较雄成蜂更为明显(图5)。结果表明,在繁育过程中及时补充糖原物质,对于延长成蜂存活时间、提升寄生效率及生殖力有着巨大的帮助;在野外应用时,配套种植合适的蜜源植物也将为防治效果的提升发挥重要的作用。
猜你喜欢
湖南林业科技(2023年5期)2023-10-31
南方林业科学(2021年3期)2021-07-28
昆虫学报(2021年5期)2021-07-07
浙江大学学报(农业与生命科学版)(2019年4期)2019-09-19
河南农业科学(2018年6期)2018-01-18
山东林业科技(2016年5期)2016-07-05
西藏科技(2015年3期)2015-09-26
意林(2015年1期)2015-05-14
科学大众·小诺贝尔(2015年5期)2015-05-04
生态学报(2013年14期)2013-09-07
