
亚硒酸钠、酵母硒和富硒螺旋藻对杂交鲟幼鱼生长、抗氧化能力及组织硒含量的影响
2024-02-15 18:14刘昊昆朱晓鸣杨云霞金俊琰解绶启
水生生物学报 2024年1期
陈 政 刘 翠 刘昊昆朱晓鸣 韩 冬,杨云霞金俊琰解绶启,
(1.中国科学院水生生物研究所淡水生态与生物技术国家重点实验室,武汉 430072;2.中国科学院大学,北京 100049;3.湖北省水产动物营养与饲料工程技术研究中心,武汉 430072)
硒作为鱼体生长发育所必需的一种微量元素,不仅是谷胱甘肽过氧化物酶和硫氧还蛋白还原酶的活性中心[1],能有效地清除氧化应激所产生的有害自由基,阻止脂质过氧化,还是脱碘酶的活性中心,可调节甲状腺激素的代谢平衡[2]。同时,硒还能有效增强机体的免疫水平和抗病能力[3],提高鱼体的生长性能[4],改善鱼肉品质[5]。在诸多生理生化功能中,抗氧化功能是硒在体内发挥作用的基础,在斑点叉尾鮰(Ictalurus punctatus)[6]、异育银鲫“中科三号”(Carassius auratus gibeliovar.CAS Ⅲ)[7]和鲈(Lateolabrax japonicas)[8]等鱼类上的研究均显示,饲料中添加硒可显著提高谷胱甘肽过氧化物酶(GSHPx)等抗氧化酶的活性,从而提高机体的抗氧化能力。
硒在自然界的存在形式主要有有机硒和无机硒两种,饲料中添加的无机硒和有机硒主要是亚硒酸钠和酵母硒。亚硒酸钠便宜易得,但毒性较大、生物利用率低、在生物体内贮积率低。快捷有效的获得有机硒主要通过利用酵母菌株或藻类的富集能力,将无机硒转化为硒蛋白。相较亚硒酸钠,有机硒是安全有效的补硒形式,硒以硒代半胱氨酸或硒代蛋氨酸的形式存在于硒蛋白中,更易被机体消化吸收,生物利用度较高且具有较高的安全性和稳定性。除酵母外,藻类也是硒有机化的良好载体,其中螺旋藻(Arthrospira platensis)含有丰富的硒蛋白、具有较高的抗氧化能力[9],因此,富硒螺旋藻也是一种具有潜力、有待开发的有机硒源。在斑点叉尾鮰上的研究显示,相较无机硒,饲料中添加有机硒更能提高鱼体的免疫力和抵抗力[10],生物效价更高,鱼体硒蓄积量更高,肝脏GSH-Px活性更高,需要量也更低[11]。在虹鳟(Salmo gairdneri)上的研究结果显示,有机硒相比无机硒有更高的硒沉积率、更好的抗应激效果、不会诱导脂质过氧化,并且与其他微量元素没有明显的互作关系[12]。相比亚硒酸钠,酵母硒能显著提高黄颡鱼肌肉硒沉积、抗氧化及抗低温应激能力[13]。此外,石斑鱼(Epinephelus malabaricus)[5]和大西洋鲑(Salmo salar)[14]的研究也表明有机硒的利用率高于无机硒。
硒作为人体必需的微量元素之一,对人类健康起着至关重要的作用。在饲料中添加硒既可以促进鱼类的健康高效养殖,也有益于通过生产富硒水产品为我国缺硒地区居民补充膳食硒。健康的成年人每日硒摄入量应为55 μg,最低摄入量为每日40 μg[15]。硒缺乏已被证实与克山病、大骨节病、动脉粥样硬化等一系列疾病有关[16]。在饮食中补充硒以维持一个最佳的硒状态对于预防癌症、糖尿病、心血管疾病及提高免疫力、改善机体亚健康状态可能有帮助[17,18]。富硒水产品可作为人类补充硒的有效途径,湖北既有稳居全国第一的淡水养殖产量,又有全球唯一探明独立硒矿床,具有生产富硒饲料原料及富硒水产品得天独厚的条件。
鲟是一种个体大,肉质鲜美的软骨硬鳞鱼,鱼卵可以做成名贵的鱼籽酱。本实验所使用的杂交鲟(Acipenser baerii♂×Acipenser schrenckii♀)为西伯利亚鲟与史氏鲟的杂交种,具有生长速度快、饲料效率高、抗病力强、营养价值高和无肌间刺等优点,具有较高的经济价值,养殖前景相当广阔[19]。目前关于鲟饲料中添加硒的研究较少,本实验旨在通过研究饲料中添加不同类型和水平的硒源对杂交鲟生长性能、抗氧化状态和硒蓄积的影响,为杂交鲟高效健康养殖及富硒水产品的生产提供数据参考。
1 材料与方法
1.1 实验饲料
以鱼粉、豆粕和菜粕为蛋白源,鱼油、豆油1∶1混合作为脂肪源,配制粗蛋白含量为45%、粗脂肪含量为10%的基础饲料,基础饲料配方及营养成分见表1。另外在基础饲料中添加亚硒酸钠(国药集团化学试剂有限公司)、酵母硒(安琪酵母有限公司)及富硒螺旋藻(由中国科学院水生生物研究所藻类生物学重点实验室提供)三种不同来源的硒,硒的添加量设低、高两个水平,硒添加量分别为0.4和1.2 mg/kg,配制6种实验饲料。饲料硒含量采用原子荧光光度计法(GB 5009.93-2017 第一法)测得,对照组、低亚硒酸钠组、高亚硒酸钠组、低酵母硒组、高酵母硒组、低富硒藻组和高富硒藻组饲料中硒的实测含量分别为1.34、1.67、2.49、1.80、2.54、1.82和2.50 mg/kg,各组相应命名为C、S1、S2、Y1、Y2、P1和P2。在饲料制备前,将所有原料均经100目粉碎机粉碎后过40目筛,按硒的添加量将3种硒源准确称取,并采用逐级预混法将其与矿物质预混料及维生素预混料等成分充分混匀,再将所有成分按配方充分混匀,加水,经双螺杆膨化饲料机(济南鼎润机械设备有限公司)制成直径为1.5 mm的膨化颗粒饲料,70℃烘干后封口防止受潮,置于-20℃冰箱中保存。

表1 饲料配方和主要营养成分(干重)Tab.1 Formulation and proximate composition of the basal diet(% in dry matter)
1.2 实验鱼和饲养管理
实验用杂交鲟幼鱼购自湖北天峡鲟业有限公司,购回后暂养于湖北省荆州市石首市石首老河长江四大家鱼原种场网箱中,投喂商品饲料,驯养2周。实验开始时将实验用鱼饥饿24h,挑选体格健壮、规格均匀、初始体重为(7.82±0.12) g的杂交鲟各55尾分别放入21个网箱(2 m×2 m×2 m)中,将所有网箱随机分为7组,每组3个重复,分别投喂7组饲料。每天饱食投喂两次,投喂时间为8:00和16:00,并记录投喂量和水温,养殖周期为62d。实验光照为自然光,水温为15—25℃,水体溶氧>6 mg/L,氨氮<0.1 mg/L,养殖水体硒的含量低于检测限度(<0.001 mg/L)。
1.3 样品采集与分析
实验结束时,在饥饿处理24h后,将各个网箱中所有实验鱼捞取计数并称重,杂交鲟终末均重约为38.88 g。每个网箱随机选取6尾鱼用MS-222麻醉,其中2尾保存于-20℃冰箱,用于测定鱼体硒含量。其余4尾鱼,用肝素钠润洗过的注射器尾静脉取血,放入抗凝离心管中,4℃ 3000 r/min离心15min,取上清液血浆置于-80℃冰箱保存,其中2尾鱼的血浆用于测定谷胱甘肽过氧化物酶(GSH-Px)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)活性及丙二醛(MDA)含量,另外两尾用于血浆常规生化指标测定。选取抽血后的2尾鱼置于冰盘上解剖,取出鱼的肝脏、背肌分别装袋置于-20℃冰箱保存,用以测得组织硒含量,剩余空壳用于分离脊椎骨并用于脊椎骨硒含量测定。
饲料成分的分析参考AOAC[20]的方法测定,水分含量采用105℃常压干燥,粗蛋白含量使用凯氏定氮仪(KD-310,Opsis,Sweden)测定,粗脂肪含量使用索氏抽提法脂肪测定仪(SoxtecTM2055,Foss,Sweden)测定,灰分含量通过马弗炉(湖北英山县建力电炉制造厂,湖北英山) 550℃充分灼烧测定。饲料、全鱼、肝脏及肌肉组织均在彻底干燥后充分粉碎搅匀,采用原子荧光光度计法(GB 5009.93-2017 第一法)测得干样硒含量。血浆谷胱甘肽过氧化物酶(GSH-Px)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)活性及丙二醛(MDA)含量均采用南京建成生物工程研究所提供的试剂盒,按照使用说明进行测试。血浆总葡萄糖(GLU)、总蛋白(TP)、白蛋白(ALB)、总甘油三酯(TG)、总胆固醇(TC)、低密度脂蛋白胆固醇(LDL-C)和高密度脂蛋白胆固醇(HDL-C)含量,均采用全自动生化分析仪(迈瑞BS-460,广东深圳)及其相应试剂盒进行测定。
1.4 数据计算及统计分析
特定生长率(Specific growth rate,SGR,%/d)=100×(ln鱼体末重-ln鱼体初重)/养殖天数
饲料效率(Feed efficiency,FE,%)=100×(终末体重-初始体重)/摄食量
采用软件SPSS 22.0(IBM SPSS Statistics 23.0,IBM,USA)进行统计分析,经正态检验及方差齐性检验后,采用Two-way ANOVA计算硒源(Source)和硒水平(Level)间的差异及两者的交互作用。然后进行单因素方差分析(One-way ANOVA),当各处理组间差异显著,对各组数据进行Ducan’s多重比较,显著性水平设为P<0.05,数据分析结果用平均值±标准误差表示(means±SE)。使用R 4.1.0 (R Core Team,奥地利维也纳)和RStudio 1.4.1106 (RStudio,美国波士顿)进行Pearson相关分析,采用“psych”包版本2.1.9计算不同组织间硒含量的相关性及组织硒含量与血浆酶活、生化指标的相关性,并使用“pheatmap”包1.0.12版本进行可视化处理。
2 结果
2.1 不同硒源和硒水平对杂交鲟生长性能和饲料利用的影响
如图1所示,不同硒源及硒水平对杂交鲟幼鱼的特定生长率和饲料效率无显著影响(P>0.05)。
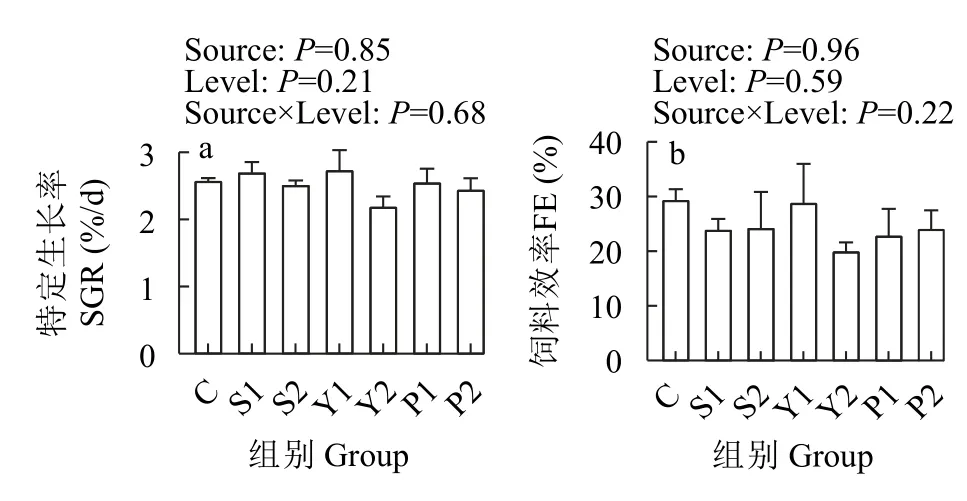
图1 不同硒源和硒水平对杂交鲟生长和饲料利用的影响Fig.1 Effects of different selenium sources and levels on growth and feed utilization of juvenile hybrid sturgeon
2.2 不同硒源和硒水平对杂交鲟组织硒含量的影响
如图2所示,不同水平的硒添加显著影响杂交鲟幼鱼全鱼和肝脏硒含量,其中低、高水平硒添加都能显著提高全鱼硒含量,但只有高水平硒添加显著提高肝脏硒含量(P<0.05)。不同硒源对杂交鲟全鱼及肝脏硒含量影响不显著(P>0.05)。硒源和硒水平对杂交鲟肌肉和脊椎骨硒含量的影响存在交互作用,其中高酵母硒组及高富硒螺旋藻组的肌肉和脊椎骨硒含量均显著高于其余各组,且高酵母硒添加组脊椎骨硒含量显著高于高富硒螺旋藻组(P<0.05)。
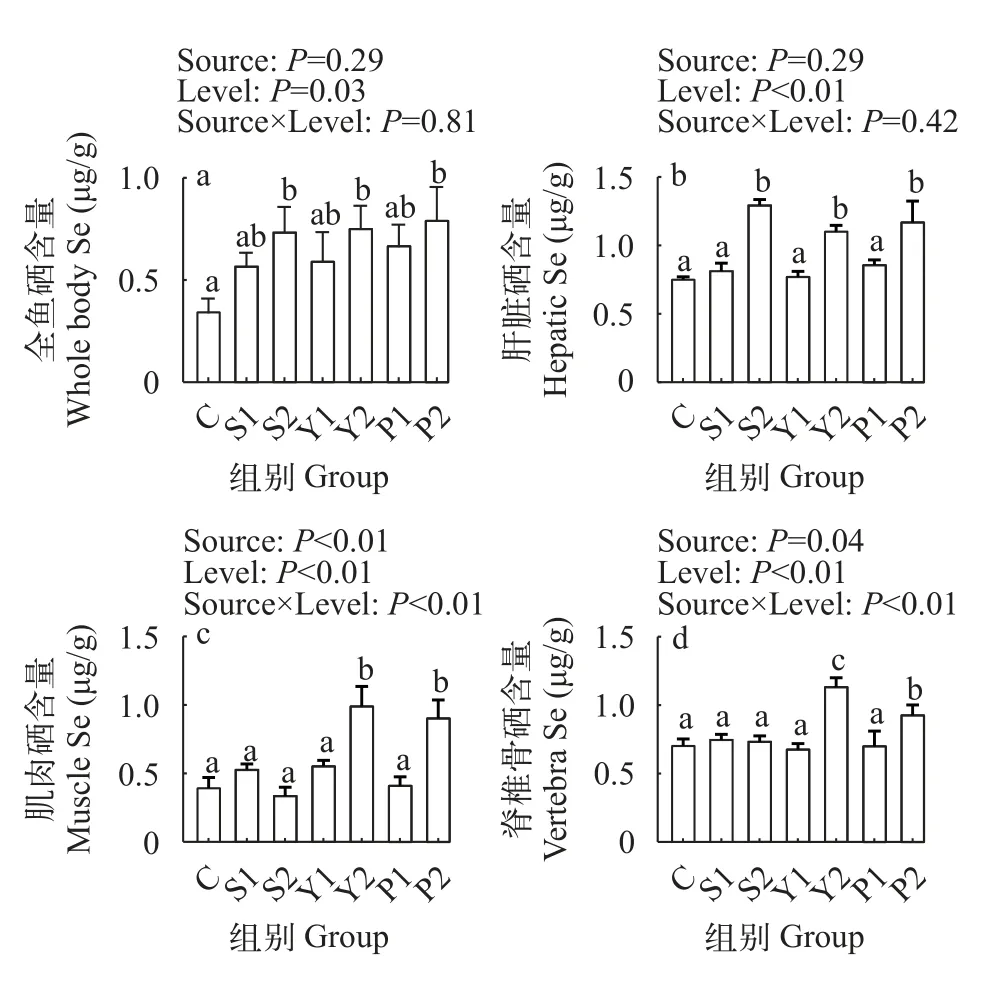
图2 不同硒源和硒水平对杂交鲟组织硒含量的影响Fig.2 Effects of different selenium sources and levels on tissue Se contents of juvenile hybrid sturgeon
2.3 不同硒源和硒水平对杂交鲟血浆抗氧化能力的影响
如图3所示,不同硒源对杂交鲟血浆GSH-Px活性影响不显著(P>0.05),但相较于对照组及低水平硒添加组,高水平硒添加可显著提高GSH-Px活性(P<0.05)。不同硒源和硒水平对杂交鲟幼鱼血浆SOD以及CAT活力无显著影响(P>0.05)。不同硒源显著影响血浆MDA含量,其中富硒螺旋藻组MDA含量显著低于亚硒酸钠组(P<0.05)。
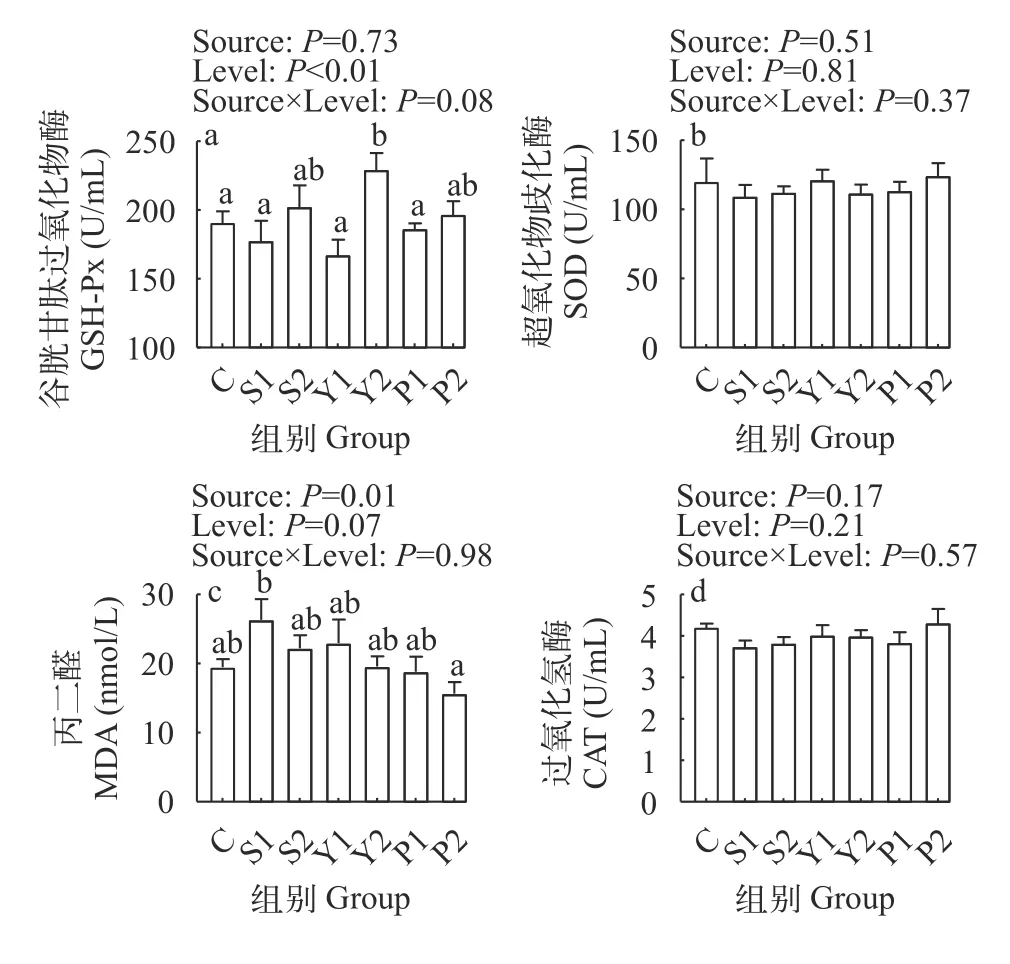
图3 不同硒源和硒水平对杂交鲟血浆抗氧化能力的影响Fig.3 Effects of different selenium sources and levels on plasma antioxidant capacity of juvenile hybrid sturgeon
2.4 不同硒源和硒水平对杂交鲟血浆生化指标的影响
如表2所示,酵母硒组血浆TP、LDL-C和TC含量均显著高于亚硒酸钠组,硒源和硒水平对杂交鲟血浆HDL-C含量的影响有交互作用,其中低酵母硒组杂交鲟血浆HDL-C含量显著高于对照组、高亚硒酸钠组、高酵母硒组和低富硒螺旋藻组(P<0.05)。血浆TC含量同时也受饲料硒水平显著影响,低硒添加可显著提高杂交鲟血浆TC含量(P<0.05)。不同硒源和硒水平对杂交鲟幼鱼的血糖、白蛋白和甘油三酯含量无显著影响(P>0.05)。

表2 不同硒源和硒水平对杂交鲟幼鱼血浆生化指标的影响Tab.2 Effects of different selenium sources and levels on plasma biochemical indices of juvenile hybrid sturgeon
2.5 相关性分析
如图4a所示,在亚硒酸钠组中,肌肉硒含量与全鱼硒含量呈显著正相关(P<0.05),但肝脏硒含量与肌肉硒含量呈显著负相关(P<0.05)。在酵母硒组中,脊椎骨硒含量与肌肉硒(P<0.05)和肝脏硒含量(P<0.01)均正相关。如图4b所示,血浆GSH-Px活性与肌肉(P<0.01)、脊椎骨(P<0.01)和肝脏硒含量(P<0.05)正相关,血浆总蛋白和白蛋白含量与肌肉硒含量正相关(P<0.05),但血浆低密度脂蛋白胆固醇与总胆固醇含量与肝脏硒含量呈显著负相关(P<0.05)。
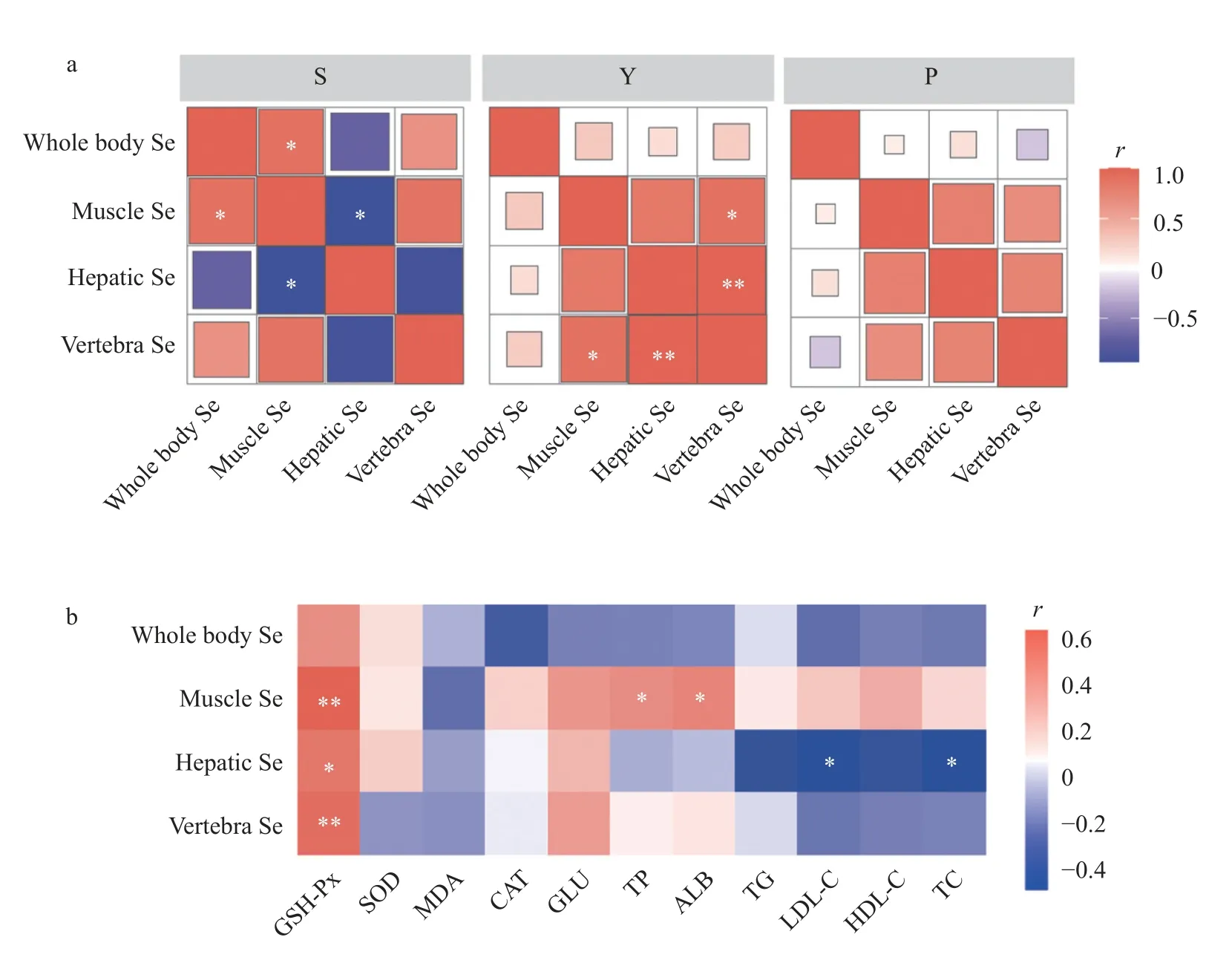
图4 杂交鲟幼鱼不同组织硒含量间的相关性(a)、杂交鲟幼鱼组织硒含量与血浆抗氧化和生化指标的相关性(b)Fig.4 Correlation plot of Se contents in different tissues of juvenile hybrid sturgeon (a),correlation plot between tissue Se contents and plasma antioxidant and biochemical indices of juvenile hybrid sturgeon (b)
2.6 富硒鲟鱼的参考摄入量
根据中国营养学会制定的中国居民膳食营养素参考摄入量,幼儿(1—3岁)每日的硒参考摄入量为20 μg,学龄期儿童(7—10岁)每日的硒参考摄入量为35 μg,青春期为40 μg,哺乳期妇女的硒参考摄入量达65 μg每日;对成年人而言,每日的硒摄入推荐量为50 μg,可耐受的每日最高硒摄入量为400 μg。在本实验中,高浓度酵母硒及富硒螺旋藻饲喂的杂交鲟肌肉硒含量较高,高酵母硒组及高富硒螺旋藻组杂交鲟鱼肉的硒营养素参考值(NRV)最高,分别为39.05%和34.14%(表3)。
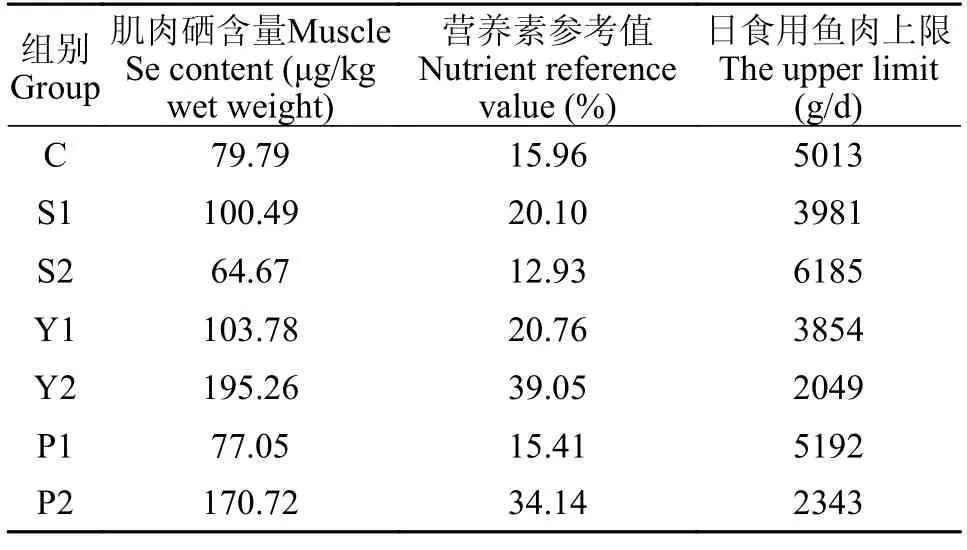
表3 成人摄入富硒杂交鲟鱼肉的硒参考摄入量及日食用量上限Tab.3 Recommended daily consumption and upper limit of fish meat when supplementing selenium for adults with only meat of hybrid sturgeon fed with different feeds
3 讨论
3.1 不同硒源和硒水平对杂交鲟生长性能和饲料利用的影响
饲料中硒的添加对养殖鱼类的生长和增重具有重要意义[6,7],但不同鱼类对硒的需求量存在差异,以亚硒酸钠为硒源时,斑点叉尾鮰的硒推荐量为0.25 mg/kg[6],大口黑鲈(Micropterus salmoide)的适宜添加水平为1.60—1.85 mg/kg[21],而黄尾鰤(Seriola lalandi)的需要量高达5.56 mg/kg[22]。作为机体必需的微量元素,硒在饲料中的含量不足或过量都会抑制幼鱼的生长[23]。目前,杂交鲟的硒需求量并不清楚,本实验中不同水平的硒添加对杂交鲟生长和饲料利用无显著影响,说明基础饲料中1.34 mg/kg的硒可能已满足杂交鲟幼鱼的需求,此时硒的额外添加对杂交鲟的生长无促进效果。不同鱼类对同一硒源的需求量不同,同种鱼类对不同硒源的需求量同样存在差异,在异育银鲫[24]、斑点叉尾鮰[11]、虹鳟[25]和石斑鱼[5]上的研究均表明有机硒相较于无机硒生物效价更高,效果更好,在饲料中的有效添加量也更低。但本实验中不同硒源对杂交鲟生长和饲料利用的影响差异不显著,原因可能是有机硒和无机硒的添加既未不足也未过量,短期内对杂交鲟的生长无促进或抑制作用,其生物效价的不同也多表现在组织蓄积等方面而非生长性能上。
3.2 不同硒源和硒水平对杂交鲟组织硒含量的影响
在本实验中,饲料中硒的添加水平对杂交鲟的全鱼和组织硒含量造成显著影响。但是,饲料中的硒仅在以高浓度有机硒(酵母硒和富硒螺旋藻)的形式添加时,才能显著提高杂交鲟肌肉和脊椎骨硒含量,亚硒酸钠或低水平硒添加提升效果不显著。这不仅说明硒在杂交鲟幼鱼体内的蓄积是剂量依赖型的,还说明不同硒源的代谢方式可能存在差异。
一般而言,相较于无机硒,有机硒更容易被机体吸收转化后富集在组织中,斑点叉尾鮰[11]上的研究表明,相较于无机硒,有机硒更容易沉积在肌肉中;且当石斑鱼[5]饲料中添加硒达到1.5 mg/kg时,有机硒添加组的肌肉硒含量是无机硒添加组的3倍多。饲料中的硒以有机形式添加时,多富集在蛋白质丰富的肌肉组织中[5,26],而以无机形式添加时,则在肝脏中代谢沉积,从而导致肝脏硒含量较高[16]。在虹鳟上的研究也表明,无机硒只能提高肝脏中的硒含量,而有机硒既可以提高肝脏硒含量又能提高肌肉中的硒含量[12],本实验的结果与之一致。高浓度有机硒(酵母硒和富硒螺旋藻)添加显著提高了杂交鲟全鱼、肝脏和肌肉硒含量,而高浓度无机硒(亚硒酸钠)添加只显著提高了杂交鲟全鱼和肝脏硒含量,且无机硒在肌肉中和肝脏中的沉积呈负相关,而有机硒则呈正相关,这说明不同类型的硒沉积模式不同。不同硒源的硒沉积规律不同,可能是因为代谢途径不同。有机硒中的硒以硒代氨基酸的形式结合在蛋白质中,能有效地增加机体硒沉积;无机硒结合人体蛋白的难度更大,大量沉积在肝脏中,大部分可能最终被肝脏或肾脏排泄至体外。另一方面,有机硒在体内的代谢周期比无机硒长[27],动物对有机硒的消化率更高,能更充分地被机体加以利用,这些都可能是导致有机硒的肌肉沉积量较无机硒高的原因。肌肉是鱼体的重要可食部分,饲料中添加适宜浓度的有机硒是生产富硒鱼肉制品的有效手段。若仅以不同饲料饲喂的杂交鲟鱼肉为成年人补硒,摄入更少的高浓度有机硒(酵母硒以及富硒螺旋藻)饲喂的杂交鲟鱼肉即可达到硒摄入的每日推荐量,饲料中高浓度有机硒添加是生产富硒水产品和为人类补硒的有效途径。
在军曹鱼(Rachycentron canadum)上的研究指出,脊椎骨硒含量随着饲料有机硒或无机硒水平的升高而升高[28],然而在本实验中,只有高水平酵母硒和富硒螺旋藻的添加显著提高了杂交鲟脊椎骨硒含量,并且高浓度酵母硒对脊椎骨硒含量的提升效果最显著,且其脊椎骨硒含量与肌肉硒含量高度正相关。但是,需要指出的是,与真骨鱼类的骨骼不同,鲟软骨不仅可食用,还具有独特的保健药用功效[29],本实验的这一发现对鲟富硒软骨的开发具有借鉴意义。
3.3 不同硒源和硒水平对杂交鲟血浆抗氧化能力的影响
高水平硒的添加可以显著提高杂交鲟幼鱼的血浆抗氧化能力。硒作为谷胱甘肽过氧化物酶(GSH-Px)的活性中心[1],其抗氧化作用主要就是通过GSH-Px清除脂质过氧化物,防止氧自由基对机体造成损伤。缺乏硒会降低GSH-Px活性从而导致机体抗氧化能力下降[23],在大口黑鲈[21]、鲈[8]、黄颡鱼(Pelteobagrus fulvidraco)[23]和石斑鱼[30]上的研究都指出,机体的GSH-Px活性会随饲料硒水平的升高而逐渐升高;类似的,本实验中饲料中高水平的硒添加显著提高了血浆GSH-Px活性,且血浆GSH-Px活性与杂交鲟肌肉、肝脏及脊椎骨硒含量正相关,说明血浆GSH-Px活性受到鱼体组织硒含量的影响。此外,在斑点叉尾鮰[11]和军曹鱼[28]上的研究都发现,有机硒相较无机硒更能提高机体GSH-Px活性,不同类型的硒提升机体GSH-Px活性的效果不一样。在本研究中,不同硒源对杂交鲟GSH-Px的影响并不显著,这与斑点叉尾鮰和军曹鱼上的结果不同,但与在黄颡鱼上发现亚硒酸钠和酵母硒都能显著提高幼鱼肝脏GSH-Px活性的结果一致[13]。
GSH-Px、SOD和CAT都具有抵抗机体氧化损伤的重要作用,是细胞防御的重要组成部分,但在本实验中,杂交鲟血浆SOD和CAT活性并未受到饲料中添加硒的类型或水平影响。之前的研究表明,饲料中过量的硒会降低异育银鲫血清CAT 和T-SOD活性[7],这也说明2.5 mg/kg的饲料硒含量未对杂交鲟的抗氧化系统造成显著的负面影响。MDA是脂质氧化产物,在西伯利亚鲟(Acipenser baeri)上的研究结果显示,不同浓度的硒添加对鲟鱼体内的MDA含量没有显著影响[31]。但虹鳟饲料中硒含量的增加能有效降低其血清和肌肉中MDA含量[32]。在本实验中,饲料硒水平对杂交鲟幼鱼血浆MDA含量影响不显著,但富硒螺旋藻组的血浆MDA含量显著低于亚硒酸钠组,这可能是由于亚硒酸钠添加减少脂质过氧化物产生的效果不显著,而螺旋藻的添加可以降低脂质过氧化物的生成[33]。
3.4 不同硒源和硒水平对杂交鲟血浆生化指标的影响
血浆总蛋白包括多种酶和抗体,具有重要的免疫作用,硒可以提高斑点叉尾鮰感染爱德华氏菌(Edwardsiella ictaluri)时血清抗体的水平,且有机硒效果更显著[10]。本实验中酵母硒的添加可显著提高杂交鲟血浆总蛋白含量,表明酵母硒的添加对提高杂交鲟的免疫力可能有一定的作用。许多研究都表明,饲料中硒的添加会影响机体的脂质代谢[34]。饲料中添加过量硒会通过促进脂质生成、下调脂质分解和激活内质网应激来增加黄颡鱼脂质沉积[35]。本实验中硒源和硒水平对杂交鲟幼鱼的脂肪代谢均产生了一定的影响,但具体机制仍需进一步的研究。
4 结论
综上所述,不同硒源和硒水平对杂交鲟生长和饲料利用无显著影响;高水平硒添加可以提高全鱼及肝脏中硒的含量,不同类型硒在鱼体的蓄积模式不一致,高水平的有机硒(酵母硒和富硒螺旋藻)添加相比无机硒(亚硒酸钠)更易在肌肉和脊椎骨中蓄积;硒与血浆GSH-Px活性呈正相关关系,高水平硒添加可显著提高杂交鲟抗氧化能力,不同硒源对杂交鲟的抗氧化能力无显著差异。通过估算,本研究中高硒饲料养殖的杂交鲟肌肉的安全食用上限可达2049 g/d,食用更少的高浓度有机硒(酵母硒及富硒螺旋藻)饲喂的杂交鲟鱼肉即可达到成年人硒的每日推荐摄入量。
猜你喜欢
石材(2020年9期)2021-01-07
医学新知(2019年4期)2020-01-02
中国盐业(2018年20期)2019-01-14
科技与创新(2017年20期)2017-11-30
山东青年(2016年2期)2016-02-28
食品界(2016年4期)2016-02-27
精神医学杂志(2015年5期)2015-12-29
世界博览(2015年17期)2015-09-10
华东理工大学学报(自然科学版)(2014年5期)2014-02-27
食品科学(2013年22期)2013-03-11
