
小麦骨干亲本周8425B抗条锈病优异基因在其衍生品种中的遗传解析
2024-02-14 00:58:14李俣佳于士男唐建卫李巧云郑继周董纯豪袁雨豪郑天存殷贵鸿
作物学报 2024年1期
李俣佳 许 豪 于士男 唐建卫 李巧云 高 艳 郑继周 董纯豪 袁雨豪 郑天存 殷贵鸿,*
1河南农业大学农学院 / 省部共建小麦玉米作物学国家重点实验室 / 河南粮食作物协同创新中心 / 国家小麦工程技术研究中心, 河南郑州 450046, 2河南丰德康种业股份有限公司, 河南郑州 450001
小麦条锈病是一类由条形柄锈菌(Pucciniastriiformisf. sp.tritici, Pst)引起的真菌性病害[1]。中国是小麦条锈病的高发地区, 且发病规律较为复杂[2],常发地区主要分布在西北、西南、华北和黄淮海等地的冬、春麦区[3]。小麦条锈菌的频繁变异和新强毒性小种的出现和流行, 尤其新生理小种条中34号(CYR34)[4]的出现, 因具有毒性谱宽、致病性强的特性[5], 使大多数小麦主栽品种“丧失”抗性, 极有可能再次引起条锈病的大流行, 造成新一轮小麦主栽品种的大规模更替。黄亮等[6]检测全国近年来推广面积较大的14个主栽小麦品种对CYR34的苗期抗性,发现除周麦22外其他13个供试品种苗期均表现感病。吴舒舒[7]用小麦条锈菌混合小种CYR32、CYR33和CYR34对270份冬小麦主产区候选品种(系)进行条锈病抗性鉴定, 表现稳定成株期抗病的新品种(系)有35份(占比12.96%), 表现稳定全生育期抗病的新品种(系)仅有2份(占比0.74%)。管方念等[8]利用由条锈菌CYR32、CYR33、CYR34、水源11-4和水源11-5组成的混合小种进行接种鉴定, 发现152份黄淮海麦区小麦农家品种中仅35份表现为稳定的成株期抗性, 7份表现出全生育期抗性。胡朝月等[9]发现在所鉴定103份抗病基因载体品系中, 仅含有Yr5、Yr15和Yr45的载体品系全生育期高抗生理小种CYR32、CYR33和CYR34。鉴定并选育抗病品种是控制小麦条锈病最为简便、经济、环保且有效的方法[10], 在成株期持久多抗基因的基础上聚合全生育期抗条锈病基因的育种策略将成为今后育种工作的重中之重[11]。
周8425B是利用六倍体小黑麦与普通小麦远缘杂交、大群体回交、辐射诱变、阶梯杂交改良和一年2次加代等技术, 创制出的重要小麦骨干亲本。因其具有抗病、抗逆、矮秆、大穗、大粒等突出优点[12], 在河南、江苏、安徽、陕西、山东、河北、甘肃、湖北等13个省市291家单位用作供体亲本广泛利用, 至2022年已育成审定衍生新品种655个, 形成了以周麦、郑麦、百农、豫麦、豫农、存麦、中麦等为代表的系列品种, 种植面积占黄淮南片麦区65%以上, 为该麦区小麦品种更新换代、生产发展及我国粮食安全和农民增产增收做出了重要贡献[13-14]。这与周8425B[15]及其子一代品种周麦11、周麦13、周麦16[16]和周麦22[17-19]的优异性状具有良好的配合力和较强的传递力有着密切关系。周8425B携带YrZH84、YrZH84.2、YrZH22、Yr30和Yr9等抗条锈病基因[20], 其中YrZH84[21]和YrZH84.2[22]是从周8425B中发掘出的2个显性抗条锈病基因,YrZH84位于7BL染色体上与SSR标记Xcfa2040和Xbarc32紧密连锁,YrZH84.2位于1BL染色体上与EST标记BE497107和CD373538紧密连锁, 均为苗期抗性(全生育期抗性)基因;YrZH22[23]是从周麦22中发掘出的抗条锈病新基因, 该基因位于4BL染色体上, 与STS标记WGGB119和WGGB124紧密连锁, 属于成株期抗性基因。Yr30(成株期抗性基因)[24]是位于3BS染色体基因簇上有待进一步发掘的兼抗条锈病和白粉病的基因[25], 与SSR标记WMS533[26]紧密连锁。Crossa等[27]证实Yr30为非小种专化抗性基因, 且在不同环境下均表达中度抗性。Yr9(全生育期抗性基因)[28]位于1RS上, 与SSR标记Xgwm582紧密连锁,1985年流行的CYR29造成其丧失抗病性。然而, 在众多的周8425B衍生品种中, 这5个抗条锈病基因的遗传分布情况尚不明确。
本研究对周8425B及其衍生品种进行条锈病抗性鉴定和分子标记检测, 以期了解YrZH84、YrZH84.2、YrZH22、Yr30及Yr9在衍生品种中的分布情况和遗传规律, 同时筛选携带抗条锈病基因且农艺性状优良的优异小麦种质资源, 为骨干亲本周8425B的持续遗传研究、小麦抗条锈病育种及抗条锈病基因的挖掘利用提供重要信息。
1 材料与方法
1.1 试验材料
供试材料为骨干亲本周8425B的衍生品种222份, 其中, 68份为本课题组保存的种质, 其余166份由各品种培育单位提供。衍生品种的审定信息从网络查询(中国种子协会网http://seedchina.com.cn/; 种业商务网https://www.chinaseed114.com/)。
试验于2020—2021、2021—2022年度在黄淮麦区南部河南农业大学许昌校区小麦试验田的条锈病鉴定圃进行。试验田土质为壤土, 地势平整, 排灌方便。10月下旬播种, 采用人工开沟点播, 每份材料种植4行, 行长1 m, 行距20 cm, 株距10 cm, 试验采用随机区组设计, 3次重复, 以高感品种铭贤169作为诱发行种植于鉴定圃走道及周边。
条锈病鉴定使用的条锈菌种由甘肃省农业科学院植物保护研究所提供。苗期鉴定采用条中34号(CYR34)单一小种, 成株期鉴定采用混合菌种, 以条中34号为代表的贵农致病类群为主, 占50.0%(其中条中34号占21.8%), 以条中32号(CYR32)为主的Hy致病类群为次要类群, 占25.0%, 以条中33号(CYR33)为主的水源11类群约占16.0%, 新的感染中四的菌系占9.0%。
1.2 试验方法
1.2.1 DNA提取 取各供试材料标准种子2粒,采用改良的CTAB法[29]提取基因组DNA。用紫外分光光度计测定DNA浓度及纯度, 并用无菌超纯水将DNA稀释至50 ng µL–1, –20℃保存。
1.2.2 抗条锈基因分子标记检测 选用已报道的与YrZH84、YrZH84.2、Yr30、YrZH22和Yr9抗条锈病基因两侧紧密连锁的分子标记对周8425B及其222份衍生品种、感病对照铭贤169进行检测(表1)。引物序列信息来自文献或从Grain Genes 2.0(http://wheat.pw.usda.gov/)网站查获, 引物均由上海生工生物工程有限公司(https://www.sangon.com/)合成。PCR反应体系10 μL, 含5 μL的2×TaqMix, 上下游引物各0.5 μL, 模板DNA 2 μL, ddH2O 2 μL。引物的扩增程序: 94℃预变性5 min; 94℃变性1 min,52~60℃退火1 min (视引物情况而定), 72℃延伸1 min, 40个循环; 然后72℃延伸10 min。PCR反应在Applied Biosystems PCR仪(赛默飞世尔科技公司Thermo Fisher Scientific)上进行。扩增产物用8%非变性聚丙烯酰胺凝胶电泳检测, 检测结果在SYNGENE G:BOX F3凝胶成像仪中观察记录。
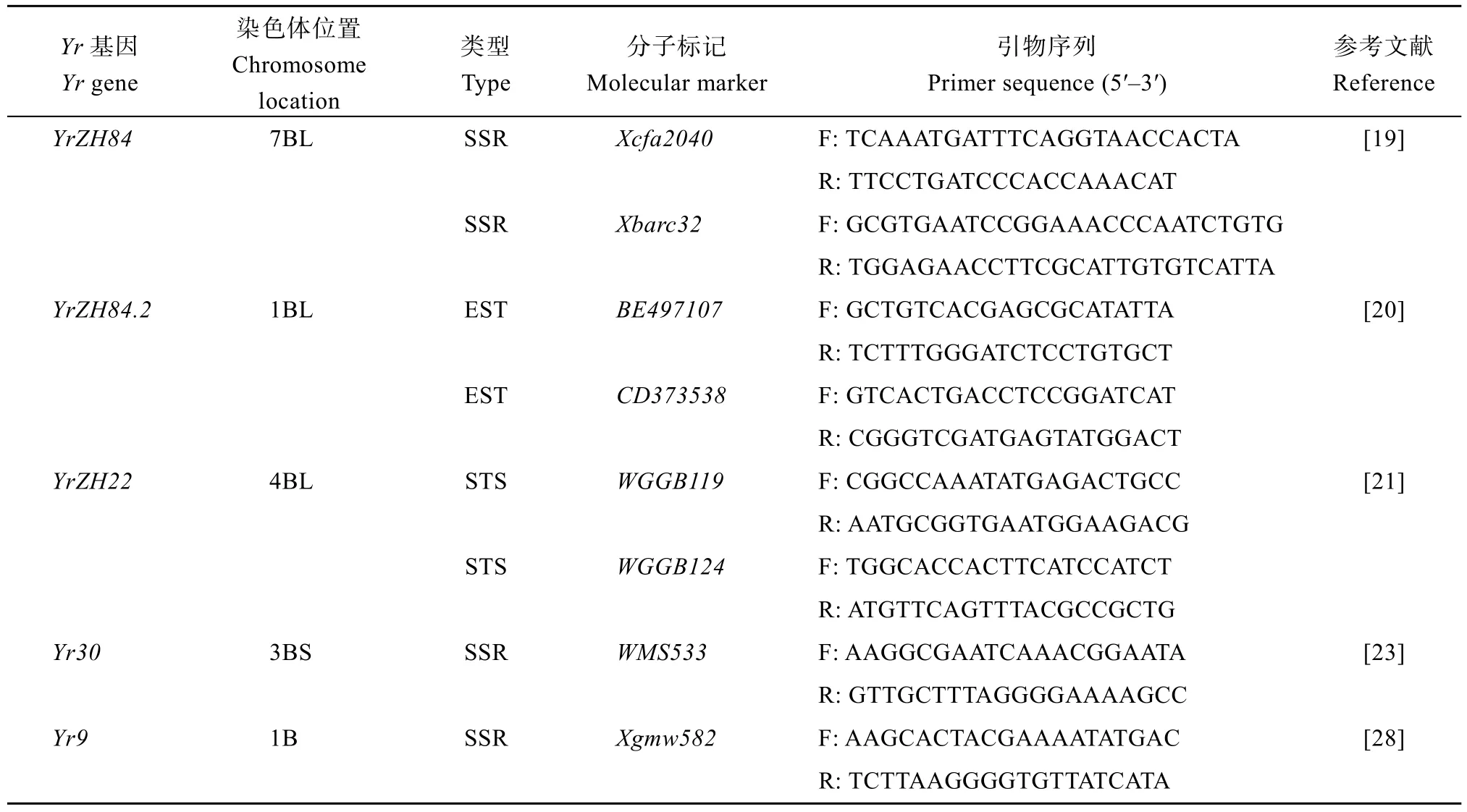
表1 抗条锈病基因的分子标记及其引物序列Table 1 Molecular markers and primer sequences for rust resistance genes
1.2.3 苗期接种抗性鉴定 2021年12月在河南农业大学农学院人工气候室进行苗期接种抗性鉴定。将供试材料种植于10 cm×10 cm口径的育苗盆中, 待小麦一叶一心时进行接种。在抖粉瓶中加入1 g的CYR34条锈菌孢子和200 g的滑石粉混合均匀。用喷壶加蒸馏水将小麦叶片多角度喷湿, 用手指蘸水脱去蜡质层, 再喷湿, 均匀抖孢子粉于叶片表面。盖上塑料罩保湿, 10℃黑暗保湿24 h。之后每天光照16 h, 温度15~20℃。在接种15~20 d后, 待感病对照材料铭贤169叶片充分发病时记载供试材料反应型, 鉴定标准按照0~9级标准记载侵染型[30],每3 d重复鉴定一次, 调查3次, 以最高反应型为最终鉴定结果[31]。
1.2.4 成株期抗性鉴定 在3月初无风阴天的下午, 将1 g以CYR34为主的条锈菌混合小种孢子溶于2 L水中, 加2 mL浓度96%的Tween-20配置成孢子悬浮液, 采用喷雾法将悬浮液喷施于诱发行进行接种。同时将1 g条锈菌混合菌种与200 g滑石粉充分混合后, 均匀撒在诱发行上进行接种, 接种后用薄膜覆盖保湿2 d。待感病对照充分发病后进行第一次抗病性调查, 分别记载反应型(infection type, IT)和严重度(disease severity, DS), 并计算病情指数。抗性记载标准, 反应型调查采用6级[32]标准, 即0、0; 、1、2、3、4, 严重度是根据病叶上孢子堆的大小和数量的多少来分级, 一般按照孢子堆占小麦叶片面积的百分数进行分级, 分别记载为0、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%和100%。普遍率用病叶数占调查总叶片数的百分比表示。病情指数等于平均严重度和普遍率的乘积。调查两次, 最终以所调查结果中最高等级为最终结果。
1.2.5 数据统计分析 利用Gephi 0.9.2软件[33]绘制周8425B衍生品种系谱图, 采用Microsoft Excel 2020和Origin 2021对数据进行整理和制图,用R语言circlize包(https://jokergoo.github.io/circlize/)进行数据可视化分析[35]。
2 结果与分析
2.1 条锈病抗性鉴定结果与综合评价
苗期条锈病接种鉴定结果显示(附表1), 骨干亲本周8425B苗期对强毒力优势生理小种条中34号表现高抗(IT=3)。222份衍生后代中, 表现中抗及以上的衍生品种共72个, 占比32.4%。其中表现免疫的衍生品种有2个, 占总数的0.9%; 表现高抗的衍生品种有8个, 占3.6%; 表现中抗的衍生品种有62个, 占27.9%。其他150份品种为感病类型, 占比67.6%。表明各主要小麦育种单位在新品种选育过程中, 周8425B苗期抗条锈病的优良性状部分得到较好的利用。
成株期条锈病抗性鉴定结果显示, 2021年和2022年衍生品种中抗病品种分别占材料数的30.0%和47.1% (表2和附表1)。周8425B及其222份衍生品种中, 只有兰天36连续2年对条锈病抗性表现近免疫, 占比0.5%; 驻麦305、安麦1350、宝景麦161、科达668、科林201、沃德麦365、西农805a、鑫华麦818、郑品麦26号和中育1220共10份衍生品种2年表现1年近免疫和1年高抗条锈病, 占比4.5%;周8425B (图1-a)和其9份衍生品种郑麦22、存麦13、百农5822、农麦22、平安518、泉麦29、天麦119、天麦166、周麦22 (图1-b) 2年均表现高抗条锈病, 占比4.1%; 许科918、周麦11、周麦26等23份衍生品种2年中表现1年高抗和1年中抗条锈病,占比10.4%; 宛麦66、周麦28、周麦36、矮抗58等21份衍生品种2年均表现中抗条锈病, 占比9.5%;其余158份衍生品种(系)表现1年中抗1年感病或2年均为感病, 这与苗期鉴定抗感品种数量(结果)较一致, 占比71.2%。

图1 周8425B和周麦22成株期抗条锈病表现Fig. 1 Resistance to stripe rust at the adult stage of Zhou 8425B and Zhoumai 22
结合成株期和苗期表型鉴定结果, 对周8425B及其222份衍生品种进行抗病类型综合评价, 周8425B表现稳定全生育期和成株期抗性; 昌麦9号、济研麦10、百农4199、赛德麦7号、郑麦103等共14份衍生品种2年表现稳定成株期抗性, 占比6.3%; 周麦11、周麦22、周麦26、周麦36、兰天36等52份衍生品种表现稳定全生育期抗性, 占比23.4% (表3)。以上筛选出的66份抗条锈病衍生品种均可作为小麦抗条锈病育种亲本材料或推广品种广泛利用。

?
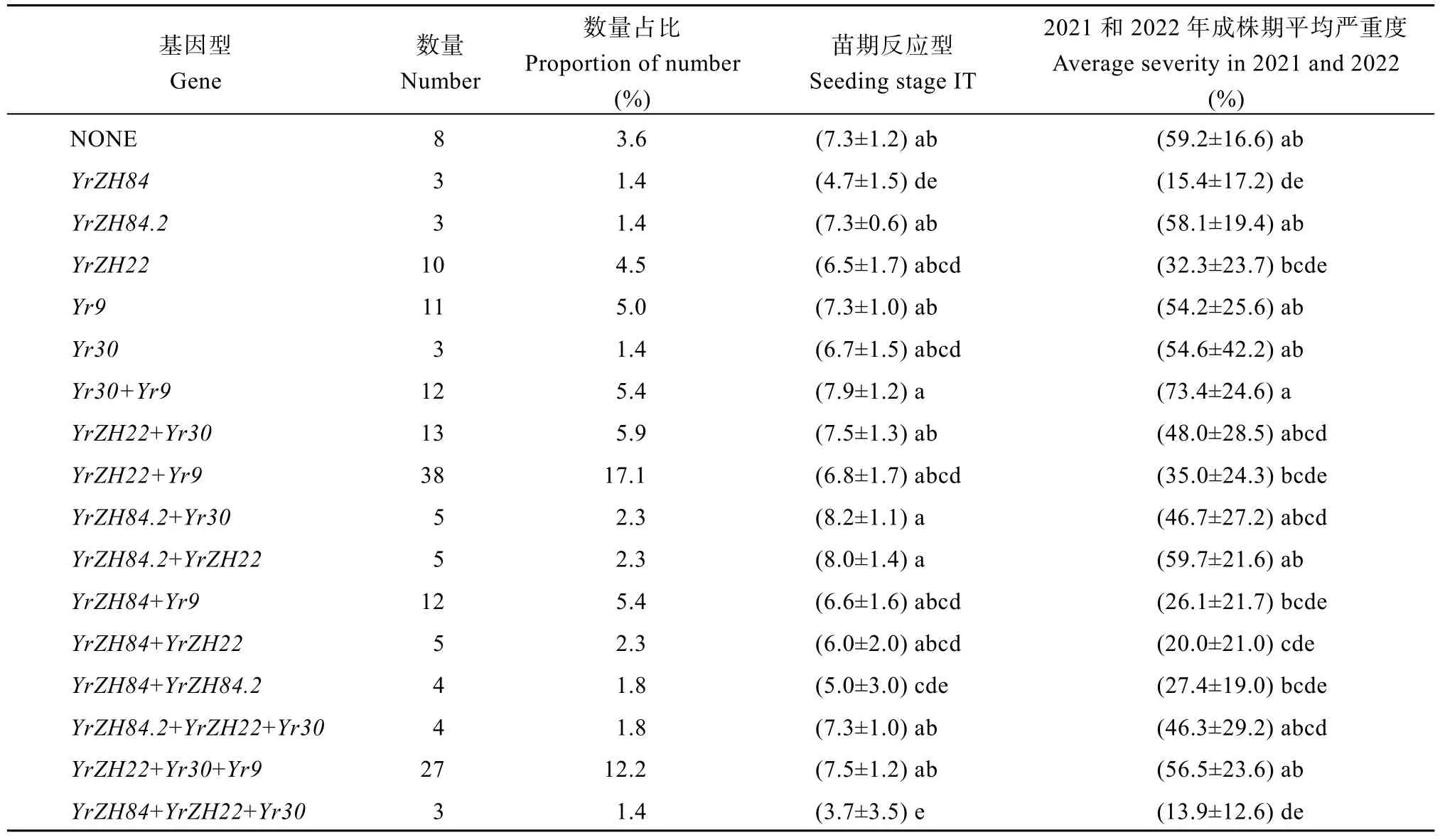
表4 不同抗条锈病基因及组合在衍生品种中的抗性效应差异Table 4 Difference of resistance effects of different stripe rust resistance genes and combinations in derived varieties
2.2 抗条锈病基因类型分布
利用紧密连锁的分子标记对周8425B及其222份衍生品种进行基因型检测(图2~图6)。结果显示周8425B携带YrZH84、YrZH84.2、Yr30、YrZH22和Yr9等5个抗条锈病基因; 在222份衍生品种中, 77份材料携带YrZH84, 占34.7%; 33份材料携带YrZH84.2, 占14.9%; 147份材料携带Yr30, 占66.2%;93份材料携带YrZH22, 占41.9%; 149份材料携带Yr9, 占67.1%。骨干种质周8425B携带的抗条锈病基因Yr9、Yr30、YrZH22和YrZH84在衍生品种中出现的频率相对较高。

图2 标记Xcfa2040检测骨干种质周8425B衍生品种YrZH84基因结果Fig. 2 PCR gragments of Xcfa2040 for YrZH84 gene in some wheat germplasms
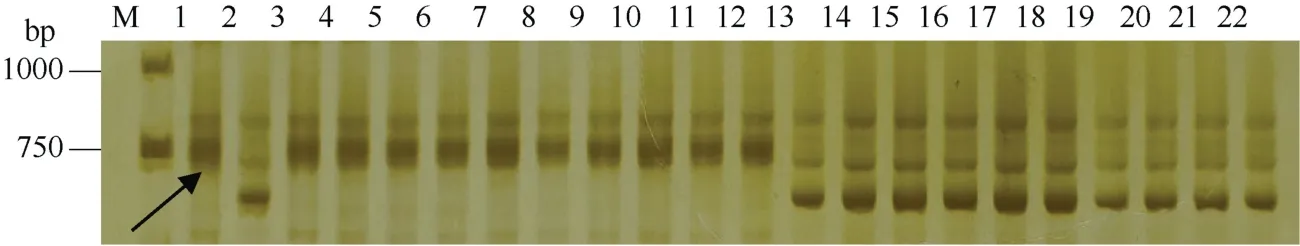
图3 标记CD373538检测骨干种质周8425B衍生品种YrZH84.2基因结果Fig. 3 PCR gragments of CD373538 for YrZH84.2 gene in some wheat germplasms

图4 标记WGGB124检测骨干种质周8425B衍生品种YrZH22基因结果Fig. 4 PCR gragments of WGGB124 for YrZH22 gene in some wheat germplasms

图5 骨干种质周8425B衍生品种的Yr30基因检测结果Fig. 5 PCR gragments of Yr30 gene in some wheat germplasms
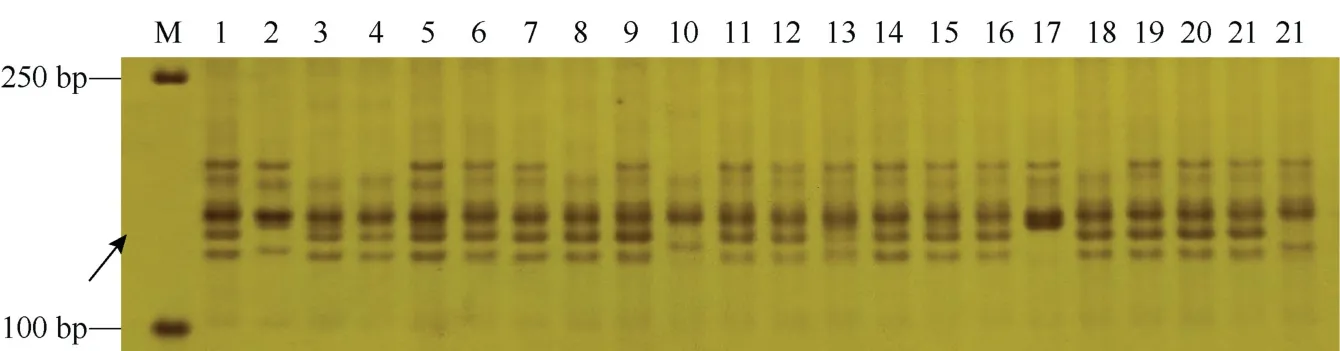
图6 骨干种质周8425B衍生品种的Yr9基因检测结果Fig. 6 PCR gragments of Yr9 gene in some wheat germplasms
2.3 不同抗条锈病基因及组合抗性效应分析
成株期抗性效应分析显示(过滤样本数小于3份的抗病基因类型), 在检测的5个基因中, 仅携带其中1个抗病基因的衍生品种以携带YrZH84的平均严重度最低, 为15.4%; 携带YrZH84.2的平均严重度最高, 为58.1%; 单个抗病基因的抗病效应为YrZH84>YrZH22>Yr9>Yr30>YrZH84.2。聚合2个抗病基因的衍生品种中, 以携带YrZH84+YrZH22的平均严重度最低, 为20.0%, 携带Yr30+Yr9的平均严重度最高, 为73.4%, 聚合2个抗病基因的抗病效应为YrZH84+YrZH22>YrZH84+Yr9>YrZH84+YrZH84.2>YrZH22+Yr9>YrZH84.2+Yr30>YrZH22+Yr30>YrZH84.2+YrZH22>Yr30+Yr9。聚合3个抗病基因的衍生品种中, 以携带YrZH84+YrZH22+Yr9的平均严重度最低, 为12.0%; 携带YrZH22+Yr30+Yr9的平均严重度最高, 56.5%, 聚合3个抗病基因的抗病效应为YrZH84+YrZH22+Yr9>YrZH84+YrZH22+Yr30>YrZH84+YrZH84.2+Yr30>YrZH84.2+YrZH22+Yr30>YrZH22+Yr30+Yr9。聚合4个抗病基因的衍生品种中, 携带YrZH84+YrZH22+Yr30+Yr9的平均严重度为16.9%, 携带YrZH84.2+YrZH22+Yr30+Yr9的平均严重度为38.4%。表明携带YrZH84或YrZH84+YrZH22、YrZH84+YrZH22+Yr9、YrZH84+YrZH22+Yr30基因组合的衍生品种成株期均具有良好的抗病性, 与不携带这5个抗条锈病基因的品种相比, 差异达显著水平。
苗期抗性效应分析显示, 在检测的5个基因中,仅携带其中1个抗病基因的衍生品种以携带YrZH84的反应型最低, 为4.7; 携带Yr9的反应型最高, 为7.3;单个抗病基因的抗病效应为:YrZH84>YrZH22>Yr30>YrZH84.2>Yr9。聚合2个抗病基因的衍生品种中,以携带YrZH84+YrZH84.2的平均反应型最低, 为5.0;以携带YrZH84.2+Yr30的反应型最高, 为8.2; 聚合2个抗病基因的抗病效应为:YrZH84+YrZH84.2>YrZH84+YrZH22>YrZH84+Yr9>YrZH22+Yr9>YrZH22+Yr30>Yr30+Yr9>YrZH84.2+YrZH22>YrZH84.2+Yr30。聚合3个抗病基因的衍生品种中,以携带YrZH84+YrZH22+Yr30的反应型最低, 为3.7;以携带YrZH22+Yr30+Yr9的反应型最高, 为7.5; 聚合3个抗病基因的抗病效应为:YrZH84+YrZH22+Yr30>YrZH84+YrZH22+Yr9>YrZH84+YrZH84.2+Yr 30>YrZH84.2+YrZH22+Yr30>YrZH22+Yr30+Yr9。聚合4个抗病基因的衍生品种中, 携带YrZH84+YrZH22+Yr30+Yr9的反应型为5.3, 携带YrZH84.2+YrZH22+Yr30+Yr9的反应型为7.0。表明苗期以携带全生育期抗性基因YrZH84或含有YrZH84的基因组合的衍生品种的抗病性较好, 与不携带YrZH84抗条锈病基因的衍生品种差异达显著水平。
2.4 抗病基因位点遗传通路分析
图7显示, 周8425B众多的衍生品种主要通过周麦11、周麦12、周麦13、周麦15、周麦16和周麦17等6个子一代再次衍生到子二代、子三代。子一代中周麦16和周麦13由于具有较好的农艺性状直接衍生出较多品种, 而周麦15、周麦17衍生品种较少; 周麦12与周麦13培育出子二代周麦22进而衍生出45个子三代, 周麦11培育出矮抗58进而衍生出54个子三代。
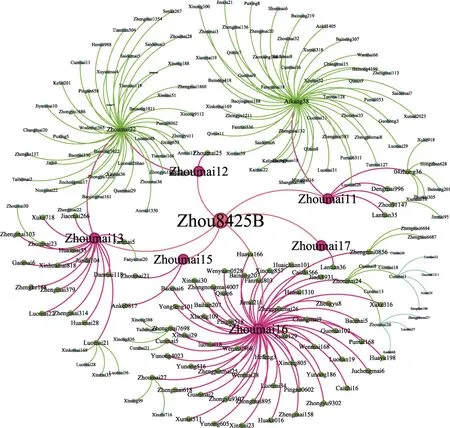
图7 周8425B衍生品种系谱图Fig. 7 Family tree of Zhou 8425B
根据携带抗条锈病基因情况, 将6个子一代与各自子代分类进行聚类分析(图8)。从图中可以看出, 子一代周麦16 (中至高感)遗传了周8425B的YrZH22+Yr9抗条锈病基因, 其衍生的68个子二代中: 携带YrZH22的有54个品种, 携带Yr9的有44个品种, 聚合了YrZH84的有16个品种, 聚合了YrZH84.2的有7个品种, 聚合了Yr30的有33个品种。

图8 周8425B及其衍生品种携带的抗条锈病基因和严重度Fig. 8 Stripe rust resistance gene and severity carried by Zhou 8425B and its derivative lines
子一代周麦11 (中至高抗)遗传了周8425B的YrZH84+YrZH22+Yr30+Yr9基因组合, 用其培育的重大品种子二代品种矮抗58 (中抗)也遗传了优异抗性基因YrZH84+Yr9组合。使用矮抗58培育的子三代54个品种中, 携带YrZH84基因的有27个品种,携带Yr9基因的有45个品种; 再次聚合YrZH22基因的有21个品种, 聚合Yr30基因的有24个品种,聚合YrZH84.2基因的有11个品种。
周麦12和周麦13培育的重大品种子二代周麦22 (高抗)携带YrZH84+YrZH22+Yr30+Yr9基因, 利用周麦22培育的子三代45个品种中携带YrZH84基因的有22个品种, 携带YrZH22基因的有40个品种, 携带Yr30基因的有24个品种, 携带Yr9基因的有35个品种, 聚合了YrZH84.2基因的有4个品种。表明周8425B的优异抗条锈病基因在遗传育种的过程中并不是完全传递到子代中, 而是在不断地分离与聚合, 可能与该4个基因分布在不同的染色体上、对手亲本抗病基因遗传背景和育种选择压有关。
3 讨论
抗病育种的选育伴随抗病性鉴定和选择, 通常需要有发病充分的鉴定条件和稳定可靠的鉴定技术体系。小麦条锈病抗性的鉴定一般分温室苗期鉴定和田间成株期鉴定。在每年都发生条锈病的地区也可以单纯依靠自然发病进行抗条锈病鉴定, 但遇到发病不利的气候和生态条件往往影响鉴定的结果。不同年份间小麦生长发育过程条锈菌侵染时的温度和湿度等环境条件会影响条锈病鉴定结果。前期温暖湿润、后期低温多雨的气候条件适宜条锈病病害流行[35]。本研究田间抗性鉴定时, 从2021年3月1日接种至5月16日表型调查期间有22天降雨, 平均最高气温18.5℃, 最低气温9.1℃; 2022年2月28日接种至5月14日表型调查期间有14天降雨, 平均最高气温19.9℃, 最低气温9.4℃ (河南省许昌市建安区历史天气https://tianqi.2345.com/wea_history/72167.htm)。因此, 在2022年的成株期抗病鉴定结果中, 抗病材料相比2021年偏多, 并且2021年的39份感病衍生品种在2022年表现抗条锈病, 这可能与2021年的气候更有利于条锈病发病有关。也有研究认为, 条锈病菌的毒性频率年度间会存在差异,即使从条锈病菌繁殖单位购买的相同混合小种在不同年份间也可能会存在一定的差异。刘志勇等[20]发现在2021年度的条锈病菌混合小种毒性较强, 而2022年度条锈病菌混合小种毒性偏弱, 相同的抗条锈病基因抗性表现比2021年要好, 本研究年度间的抗性差异可能也与混合小种毒性差异有关。
周8425B作为中国重要的小麦骨干亲本之一,在全国范围内被广泛用作亲本使用[36]。尽管周8425B对目前流行的条中32号、条中33号及条中34号小种具有较好的抗性, 但苗期和成株期条锈病抗性鉴定发现, 其部分衍生品种对强毒力优势生理小种条中34号抗病性一般。究其原因, 主要与衍生品种所含抗病基因类型和数量与周8425B存在较大差异有关, 周8425B含有的5个抗条锈病基因分布在多条染色体上, 不利于多个基因同时传递到子代中, 在222个衍生品种中仅有15个品种携带了4个基因。特别是以携带全生育期抗性基因YrZH84或含有YrZH84的基因组合的衍生品种的苗期抗病性较好。其次, 衍生品种是由全国10多个省市290多家小麦育种单位选育, 育成时间从2000年一直延续到2022年, 育成时间跨度大, 地域覆盖广, 选育时所处的条锈病菌优势生理小种和选择压不同所致。周8425B衍生后代多具有优良的丰产性、株叶型, 这类性状容易选择, 且受环境、气候影响较小, 易被育种家选择进而传递, 而抗条锈病选择容易受到年度间、麦区间、抗性选择压等因素有较大影响, 没有达到像株叶型改良利用一样的明显效果。对比发现,本研究检测衍生品种的条锈病抗性与当初审定时鉴定结果存在部分不一致, 214个衍生品种当年审定时,其中慢条锈病品种34个, 占15.9%, 对条锈病表现抗性的品种102个, 占47.7% (附表1)。肖永贵等于2019—2010年利用条锈菌优势小种CYR32对周8425B及其50个衍生品种(与本试验中17个衍生品种重复)进行成株期抗性检测, 供试材料整体趋向抗条锈病表现型[12]。而在本试验的条锈病成株期检测结果中仅有28.8%的衍生品种对条锈病表现稳定的中抗及以上抗性水平, 抗性品种数量显著减少, 可能与条锈菌优势小种CYR34的毒性更强有关, 需要今后要高度重视对CYR34的监测和抗性遗传育种研究应用。总之, 在聚合多个抗条锈病基因育种时,不仅需要田间人工接种菌种进行抗性鉴定, 还需要借助快速发展的分子标记辅助选择进行抗条锈病基因的精准鉴定。
抗病基因多样化是防止育成品种中抗条锈病基因单一化, 延缓抗病品种抗病性丧失, 延长其生产上推广利用时间的有效策略。抗病基因聚合是培育持久多抗小麦品种的重要途径。在抗病基因聚合的策略上, 国外的育种家, 尤其是以国际玉米小麦改良中心(CIMMYT)为代表的育种家倡导聚合多个成株期抗条锈病基因, 并且育成了一些聚合3~4个成株期抗条锈病基因的品种, 可达到成株期高抗甚至免疫程度的条锈病抗性[37]。而结合我国的小麦条锈病菌传播特点和育种实践, 需要采用在成株期持久多抗基因的基础上聚合全生育期抗条锈病基因的策略, 使育成品种在苗期和成株期均保持较好的条锈病抗性, 以达到有效控制小麦条锈病的传播和危害。周8425B能够作为骨干种质被广泛应用, 可能与其聚合了多个全生育期抗性和成株期抗性基因,具有持久抗性有关。鉴于周8425B衍生品种众多且利用的更加广泛, 今后需要加强周8425B携带的优异抗条锈病基因的克隆及分子机制研究, 同时做好周8425B携带的优异抗条锈病基因在西北春麦区、长江中上游麦区等条锈病发生危害的“策源地”抗性布局和利用, 减缓强毒力生理小种出现频率, 严防黄淮等小麦主产区小麦品种的抗条锈病突然“丧失”,保障国家小麦生产持续增产。
4 结论
周8425B苗期和成株期对目前强毒力优势菌种CYR34均表现高抗条锈病。筛选出稳定成株期抗病的衍生品种14份, 稳定全生育期抗病的衍生品种52份。周8425B携带的YrZH84、YrZH84.2、YrZH22、Yr30和Yr9抗条锈病基因主要通过周麦11、周麦12、周麦13、周麦15、周麦16和周麦17等6个子一代再次传递和聚合到衍生品种中。在衍生后代中的频率分别为34.7%、14.9%、41.9%、66.2%和67.1%。携带全生育期抗性基因YrZH84或含有YrZH84的基因组合的衍生品种具有较好的苗期和成株期条锈病抗性。研究结果为中国小麦骨干种质周8425B的持续改良利用提供了参考。
附表请见网络版: 1) 本刊网站http://zwxb.chinacrops.org/; 2) 中国知网http://www.cnki.net/;3) 万方数据http://c.wanfangdata.com.cn/Periodicalzuowxb.aspx。
猜你喜欢
西北园艺(果树)(2024年3期)2024-05-25 08:46:31
扬州大学学报(农业与生命科学版)(2023年4期)2024-01-09 13:54:56
Computer Modeling In Engineering&Sciences(2022年6期)2022-07-02 07:43:24
南方农业学报(2022年11期)2022-03-18 05:55:26
种子(2020年9期)2020-10-22 13:22:48
农民致富之友(2019年31期)2019-11-23 06:07:01
中国种业(2019年6期)2019-06-25 02:28:44
植物保护(2017年4期)2017-08-09 01:18:38
中国马铃薯(2017年1期)2017-03-02 09:15:51
植物保护(2017年3期)2017-01-21 11:38:48
