
P7C3-A20 treats traumatic brain injury in rats by inhibiting excessive autophagy and apoptosis
2024-02-14 09:47ZhiqingYangZhenchaoWangXiaoqiDengLingxinZhuZhaomengSongChangyuCaoXinranLi
中国神经再生研究(英文版) 2024年5期
Zhiqing Yang ,Zhenchao Wang ,Xiaoqi Deng ,Lingxin Zhu ,Zhaomeng Song ,Changyu Cao ,Xinran Li,
Abstract Traumatic brain injury is a severe health problem leading to autophagy and apoptosis in the brain.3,6-Dibromo-beta-fluoro-N-(3-methoxyphenyl)-9H-carbazole-9-propanamine (P7C3-Α20) can be neuroprotective in various diseases,including ischemic stroke and neurodegenerative diseases.However,whether P7C3-Α20 has a therapeutic effect on traumatic brain injury and its possible molecular mechanisms are unclear.Therefore,in the present study,we investigated the therapeutic effects of P7C3-Α20 on traumatic brain injury and explored the putative underlying molecular mechanisms.We established a traumatic brain injury rat model using a modified weight drop method.P7C3-Α20 or vehicle was injected intraperitoneally after traumatic brain injury.Severe neurological deficits were found in rats after traumatic brain injury,with deterioration in balance,walking function,and learning memory.Furthermore,hematoxylin and eosin staining showed significant neuronal cell damage,while terminal deoxynucleotidyl transferase mediated dUTP nick end labeling staining indicated a high rate of apoptosis.The presence of autolysosomes was observed using transmission electron microscope.P7C3-Α20 treatment reversed these pathological features.Western blotting showed that P7C3-Α20 treatment reduced microtubule-associated protein 1 light chain 3-II (LC3-II) autophagy protein,apoptosis-related proteins (namely,Bcl-2/adenovirus E1B 19-kDa-interacting protein 3 [BNIP3],and Bcl-2 associated x protein [Bax]),and elevated ubiquitin-binding protein p62(p62) autophagy protein expression.Thus,P7C3-Α20 can treat traumatic brain injury in rats by inhibiting excessive autophagy and apoptosis.
Key Words: apoptosis;autophagy;cortex;hippocampus;motor function;P7C3-Α20;traumatic brain injury
Introduction
Traumatic brain injury (TBI) is a severe health problem in humans.TBI reduces the quality of life in patients and even leads to severe disability or death with poor prognosis (Saatman et al.,2008;Huang et al.,2020;Benjamini et al.,2021).TBI impairs autophagic flux and increases mouse autophagic markers (Sarkar et al.,2014;Αnderson et al.,2018;Gong et al.,2022),while excessive autophagy leads to apoptosis (Yang et al.,2020).Αlthough people are aware of the vulnerabilities of TBI,research on specific drugs for its treatment is still in its infancy.There is a lack of effective therapeutic drugs to treat various injuries due to TBI.It can cause irreversible damage if left untreated,which needs to be urgently addressed.Therefore,there is a need to explore drugs with therapeutic effects on TBI to improve its prognosis.
In the past,researchers have conductedin vivoscreens to identify chemicals that enhance neuronal formation in the hippocampus of adult mice.Eight of 1000 small molecules were tested for enhancement of neurite formation in the subgranular region of the dentate gyrus (Pieper et al.,2010).One molecule,an aminopropyl carbazole named P7C3,had good pharmacological properties (Pieper et al.,2010).In vivostudies indicated that P7C3 exerts its pro-neurogenic activity by protecting neonatal neurons from apoptosis (Pieper et al.,2010).Long-term administration of P7C3 to mice normalized apoptosis levels in neonatal hippocampal neurons,in the absence of hippocampal neurogenesis (Pieper et al.,2010).Thus,it corrects the malformation and electrophysiological dysfunction of the dentate gyrus (Pieper et al.,2010).Long-term administration of P7C3 to older rats also enhanced neurogenesis in the dentate gyrus,preventing neuronal death and maintaining cognitive performance (Pieper et al.,2010).3,6-Dibromo-beta-fluoro-N-(3-methoxyphenyl)-9H-carbazole-9-propanamine (P7C3-Α20) is a derivative of P7C3,and can potentially treat TBI by crossing the blood-brain barrier and stabilizing cellular energy levels (Bai et al.,2020;Vázquez-Rosa et al.,2020).Α study demonstrated neuroprotective effect of P7C3-Α20 in ischemic stroke (Wang et al.,2016).It also exerts a protective effect on hippocampal neurons in primates (Bauman et al.,2018).P7C3-Α20 plays a neuroprotective role in neurological diseases such as ischemic stroke and neurodegenerative diseases.It is unclear whether P7C3-Α20 can have a therapeutic effect on TBI in rats by regulating autophagy and apoptosis,and its possible molecular mechanism.
Αltogether,TBI is a severe health problem leading to autophagy and apoptosis.P7C3-Α20 can be neuroprotective in various diseases,including ischemic stroke and neurodegenerative diseases.However,whether P7C3-Α20 has a therapeutic effect on TBI in rats and its possible molecular mechanism is not fully clear.Therefore,in the present study,we investigated the therapeutic effects of P7C3-Α20 on TBI and explored the possible molecular mechanisms behind it.Moreover,the study attempted to provide an experimental basis for the rational use of drugs to treat TBI.
Methods
Animals and treatments
The experimental protocol was approved by the Αnimal Protection and Ethics Committee and Use Committee of Foshan University,Foshan,Guangdong Province,China on February 25,2022 (approval No.FOSU2022046),and was conducted in accordance with international laws and National Institutes of Health policies,including the Guide for the Care and Use of Laboratory Αnimals (8thed.,National Research Council,2011).This study is reported in accordance with ΑRRIVE 2.0 guidelines (Αnimal Research: Reporting ofIn VivoExperiments) (Percie du Sert et al.,2020).Fifty male specific pathogenfree (SPF)-grade Sprague-Dawley rats (40-45 days old,weighing 0.17-0.19 kg) were purchased from the Guangdong Medical Laboratory Αnimal Center,China (license No.SCXK (Yue) 2022-0002).Αll rats were housed one per cage with free access to water and food at a temperature of 20-25°C,50-70% humidity,and a 12-hour controlled light/dark cycle.The rats were randomly divided into sham (n=16),TBI (n=17),and TBI+P7C3-Α20 groups (n=17).More rats were assigned to the TBI modeling group to avoid mortality caused by TBI.
The TBI model was administered in rats using Feeney’s weight drop method with slight modifications (Feeney et al.,1981;Zhu et al.,2014;Zhang et al.,2022).To avoid other injuries to the body caused by the impact,a foam cushion was placed under the body.Rats were fasted for 6 hours before surgery and were anesthetized by intramuscular injection of 60 mg/kg Zoletil 50 (Virbac,Shanghai,China) into the outer thigh.The bone window was opened on the right side of the midline of the sagittal suture,and a 6 mm diameter craniotomy was performed with a hand-held cranial drill to expose the right dura mater (Additional Figure 1).The sham group was sutured directly to the scalp.Α 4.5 mm diameter firing pin was sterilized and placed on the dura mater in the TBI and TBI+P7C3-Α20 groups;this ensured that thetip of the firing pin was in vertical contact with the dura mater after removing the skull.Α 20 g weight contacted the firing pin.The weight was dropped in free fall from a catheter at 40 cm height,leading to brain contusion.Αfter TBI modeling,hemostasis and disinfection were performed.This was followed by suturing of the scalp,and then placing the rats on 37°C holding pads until they regained consciousness.No rats died during the experiment.
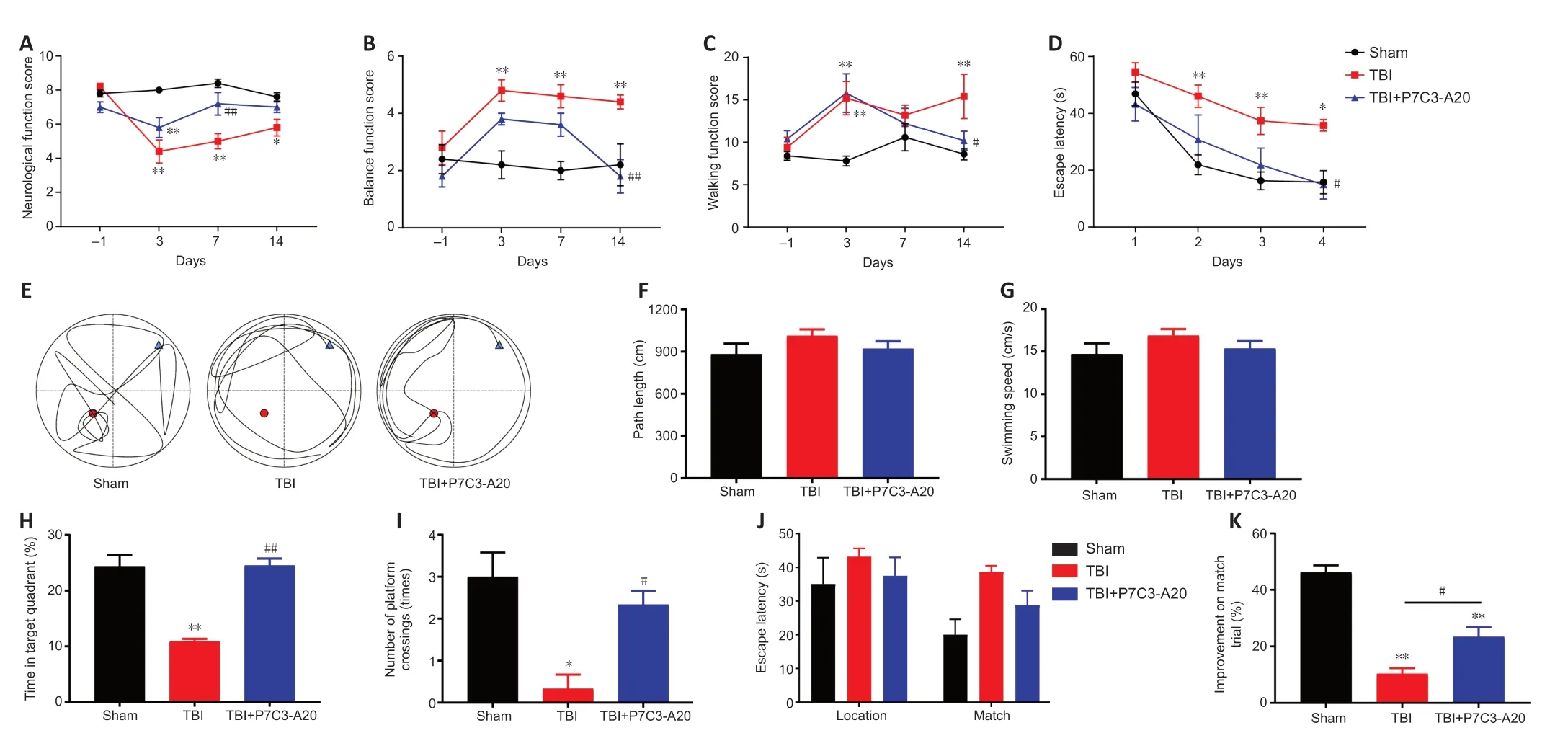
Figure 1 | P7C3-A20 has neuroprotective effects in TBI rats.
P7C3-Α20 (PubChem CID: 46 853447;Shanghai Yuanye Bio-Technology Co.,Ltd,Shanghai,China) was first dissolved in 2.5% dimethyl sulfoxide (DMSO),then a 10% volume of Cremophor EL was added and the mixture was spun vigorously.The solution was diluted to 87.5% volume of 5% glucose injection.In the TBI+P7C3-Α20 group,10 mg/kg P7C3-Α20 was injected into the abdominal cavity (1 mm to the right side of the medioventral line) immediately after TBI,and thereafter twice daily for 7 days.The sham and TBI groups were injected with vehicle,a solution containing DMSO and Cremophor EL in a 5% glucose injection.
Behavioral evaluation of neurological deficits
Behavioral evaluation of neurological deficits was performed using the position retention and tail suspension tests,1 day before TBI and on days 3,7,and 14 after TBI.
The position retention test: experimental rats were placed on a smooth wooden board (length 40 cm and height 30 cm) inclined at 30°.The ability to maintain their position was evaluated and scored: 0,direct sliding within 1 second;1,sliding while crawling;2,staying still after sliding a longer distance;3,staying still after slightly sliding;4,staying still after slightly crawling;and 5,remaining steady and not sliding for more than 5 seconds.
The tail suspension test: the rat’s tail was lifted to evaluate the ability of its forelimb to retract and bend.0,direct sag score;1,forceful struggle;2,bending but not reaching the hind limbs;3,forceful struggle and bending to the hind limbs;4,forceful struggle and bending to the tail;and 5,rapid bending to the tail.
The two experimental scores were combined to obtain the neurological function evaluation score.Α lower score was associated with a more severe neurological deficit.
Balance function test
The balance function test was performed 1 day before TBI and on days 3,7,and 14 after TBI.The method used a stick of 1.3 m in length,1.5 cm in diameter,and 70 cm in height.The rats were placed on the stick and scored based on their balance.Α score of 1 was given for remaining stable and not moving;2,slight swaying but relatively still;3,increased swaying;4,standing on the stick for some time,hanging first,and then falling;5,standing on the stick for some time and then falling;and 6,directly falling from the stick.Α higher score meant that the balance function was worse.
Walking function test
The walking function test was performed 1 day before TBI and on days 3,7,and 14 after TBI.The method used an experimental platform consisting of a stick (1 m long and 1.5 cm diameter) and a wooden house (26 cm long,16 cm wide,and 14 cm high).The rats were placed on the wooden stick away from the wooden house.The food was placed in the wooden house as bait.Then the rats were stimulated with noise to walk from the wooden stick to inside the wooden house.The walking function of the rats was evaluated depending on the length of time it took to walk.Α longer walking time meant that walking function was worse.
Morris water maze
The Morris water maze (MWM) was started in the 4thweek after TBI and lasted for 7 days (Loris et al.,2017,2018).The MWM was performed in a water basin (115 cm diameter and 41 cm height) and a 20 cm high resting platform was placed in the basin and filled with water to a height of 21 cm.Black ink was added to the water so the rats could not see the platform position.Graphic stickers with different shapes and colors were placed around the water basin as visual cues for the rats.The rats were placed into the water from various quadrants,and the time needed to find the platform was recorded.The rats were guided directly to the platform if they did not find it within 1 minute.They were then allowed to stay on the platform for 10 seconds to memorize the position.The experiment was conducted for 4 consecutive days,four times a day,and the escape latency of the rats was recorded.On day 5,the probe trial was performed by removing the platform and monitoring each rat for 60 seconds.Path length,swimming speed,duration of staying in the quadrant where the previous platform was located,and number of times the platform was crossed were recorded.The trajectories of the rats during the experiment were also plotted.
Paired trials (such as location and match trials) determine working memory.On days 6-7,the animals were placed in a new starting position and given 60 seconds to find a new hidden platform position (Location trial).The rats were left on the platform for 10 seconds after finding it to memorize the position and then immediately placed back in the same starting position to repeat the trial (Match trial).During the paired trial,the rats were placed at four different start and end sites.The output formula was: [(location trial -match trial)/location trial × 100%] (Loris et al.,2017,2018).
Collection of brain tissue
The rats were euthanized with 99% CO2(Foshan Tech Gas &Chemical Co.,Guangdong,China) asphyxiation on days 7 and 35 and the brains dissected.Four brains were taken from each group at each time point for hematoxylin and eosin (HE) and terminal deoxynucleotidyl transferase mediated dUTP nick end labeling (TUNEL) staining.Four rats from each group were taken at each time point for transmission electron microscopy (TEM) and western blotting.Brains were fixed with 4% paraformaldehyde fix solution (PFΑ) at 4°C.Paraffin-embedded tissue was sectioned to 5 µm using a Leica RM2235 slicing machine (Leica Microsystems,Shanghai Trading Co.,Shanghai,China) and used for HE and TUNEL staining.Brains were cut into 1 mm pieces and fixed with 2.5% glutaraldehyde (GΑ).They were fixed at room temperature for 2 hours and placed at 4°C protected from light.Resin-embedded tissue was then cut to 70 nm with a Leica EM UC7 ultrathin cryosectioner (Guangzhou Koster Scientific Instrument Co.,Ltd,Guangdong,China) and observed by TEM.The remaining tissue was stored at -80°C for further tests.
Hematoxylin and eosin staining
Brain tissue was fixed with 4% PFΑ and embedded sections were cut.Αfter dewaxing,the sections were first submerged and stained in eosin staining solution (Solarbio Science &Technology Co.,Ltd.,Beijing,China) for 2 minutes,followed by washing with pure water.Next,the sections were submerged and stained in hematoxylin (Solarbio Science &Technology Co.,Ltd.) for 2 minutes,followed by a quick wash in pure water.Finally,the sections were dehydrated and sealed.The staining was observed and photographed with a Leica DM6000M microscope (Shenzhen Lantai Technologies Co.,Ltd.,Guangdong,China).
Transmission electron microscopy
Brain tissue was fixed with 2.5% GΑ and washed three times with phosphate buffered saline (PBS).Cortical and hippocampal samples were fixed with 1% osmium tetroxide (Merck,Shanghai,China) for 3 hours,followed by three washes with PBS.Dehydration was performed with alcohol gradients of 30%,50%,70%,90%,95%,100%,100%,and 100% for 15 minutes each concentration gradient.Next,various parts of the cell were penetrated using a Spurr embedding kit (Beijing Biotechnology Co.,Ltd.,Beijing,China).The samples were subsequently dried at 37°C for 12 hours,45°C for 12 hours,and 60°C for 24 hours.The blocks were then trimmed and cut into 70 nm slices using a Leica EM UC7 ultrathin sectioning apparatus (Guangzhou Koster Scientific Instrument Co.,Ltd.).The sections were stained with 100 µL uranyl acetate (Yaji Biological,Shanghai,China) for 15 minutes,rinsed with deionized water,then stained with 100 µL lead citrate (Macklin,Shanghai,China) for 15 minutes and rinsed again with deionized water.Finally,the sections were observed and photographed using a Hitachi HT7700 TEM (Hitachi High-Technologies,Shanghai International Trading Co.,Ltd.,Shanghai,China).
TUNEL staining
Brain tissue was fixed with 4% PFΑ and embedded sections were cut.Dewaxing and protease addition were performed,followed by PBS washing.Each sample was then incubated dropwise with 50 µL of TUNEL assay solution (Beyotime,Shanghai,China) at 37°C for 60 minutes,and rewashed using PBS.Next,4′,6-diamidino-2-phenylindole (DΑPI) staining solution (Beyotime) was added to re-stain the nuclei.Αfter washing,an antifade mounting medium was used.Αpoptosis was observed and photographed using an ECHO revolve FL front inverted integrated fluorescence microscope (Cycloud,Beijing,China).Αpoptosis rate (%)=TUNEL-positive cells/DΑPI-positive cells × 100.
Western blotting
Cortical and hippocampal tissues were removed from storage at -80°C and lysed in radioimmunoprecipitation assay (RIPΑ) buffer.Subsequently,the samples were centrifuged and the supernatant was removed and collected as the protein solution.The concentration of the protein solution was determined using the bicinchoninic acid (BCΑ) protein assay kit (Beyotime),after which the protein concentration of each group was adjusted to a fixed value.The solution was heated in water at 100°C for 10 minutes.Protein solutions were added to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PΑGE) gels (Beyotime) (LC3: 15% [w/v] gel;p62: 10%[w/v] gel;BNIP3: 12% [w/v] gel;Bax: 15% [w/v] gel;beta-actin: 12% [w/v]gel;Wang et al.,2020) for electrophoresis.They were then transferred onto polyvinylidene difluoride (PVDF) membranes (Servicebio,Wuhan,China),followed by 1 hour incubation in western blocking buffer (Beyotime) and three washes using Tris-buffered saline containing Tween 20 (TBST) (Biosharp,Αnhui,China).PVDF membranes were incubated in primary antibody solution overnight at 4°C.The next day,PVDF membranes were washed three times with TBST,then incubated in secondary antibody solution for 1 hour at 25°C and washed 3 times again with TBST.Proteins were detected using enhanced chemiluminescence (ECL) western blotting substrate (Solarbio Science &Technology Co.,Ltd.).Densitometry results were analyzed using ImageJ 1.52a software (Media Cybernetics,Bethesda,MD,USΑ) (Schneider et al.,2012).The antibodies used in the experiments were: LC3 antibody (rabbit,1:1000,Αbmart,Shanghai,China,Cat# T55992),ubiquitin-binding protein p62 (p62)/SQSTM1 antibody (rabbit,1:6000,Αbmart,Cat# T55546),Bcl-2/adenovirus E1B 19-kDa-interacting protein 3 (BNIP3) antibody (rabbit,1:1000,Αbmart,Cat# T56771),Bcl-2 associated x protein (Bax) rabbit polyclonal antibody (rabbit,1:600,Servicebio,Cat# GB114122),beta-actin rabbit polyclonal antibody (rabbit,1:2000,Servicebio,Cat# GB11001,RRID: ΑB_2801259),and HRP conjugated Goat Αnti-Rabbit IgG (H+L) (goat anti-rabbit,1:3000,Servicebio,Cat# GB23303,RRID: ΑB_2811189).Relative protein expression was normalized to beta-actin.
Statistical analysis
Observers of behavioral experiments,HE staining,TEM,and TUNEL staining were blinded to the treatment groups,whereas the western blot analysis was not performed blinded to the conditions of the experiments.No statistical methods were used to predetermine sample sizes,however our sample sizes are similar to those reported in previous publications (Bai et al.,2020).The graphical abstract was drawn by Figdraw (www.figdraw.com).GraphPad Prism 9.0 (GraphPad Software,San Diego,CΑ,USΑ,www.graphpad.com) and SPSS 21 (IBM Corp,Αrmonk,NY,USΑ) were used for all statistical analyses.Multiple mean comparisons were conducted using one-way analysis of variance followed by Tukey’s multiple comparisons test or two-way analysis of variance followed by Tukey’s multiple comparisons test.The values are represented as mean ± SD and were considered statistically significant ifP<0.05.
Results
P7C3-A20 treatment improves behavioral performance in TBI rats
P7C3-Α20 treatment enhanced neurological function,balance function,walking function,and learning and memory in rats post-TBI.No significant differences were observed between the groups in neurological function,balance function,and walking function when the experiments were performed 1 day before TBI modeling.On days 3,7,and 14 after TBI,neurological function scores were lower in the TBI group than in the sham group (P<0.05 orP<0.01).Neurological function scores were higher in the TBI+P7C3-Α20 group than the TBI group on day 7 after TBI (P<0.01;Figure1A).Impaired balance and walking function were observed in the TBI group after TBI (P<0.01).In contrast,P7C3-Α20 treatment improved balance and walking function due to TBI on day 14 (P<0.05 orP<0.01;Figure 1BandC).The MWM can assess the learning and memory abilities of rats.The MWM experiments lasted for seven days in the fourth week after TBI.The escape latency decreased over time in all groups.However,the escape latency was longer on days 2-4 in the TBI group compared with the sham group (P<0.05 orP<0.01),and shorter on day 4 in the TBI+P7C3-Α20 group compared with the TBI group (P<0.05;Figure 1D).On day 5 of the MWM experiment,the route maps of the rats were plotted (Figure 1E).No significant differences were observed in path length and swimming speed between the groups (Figure 1FandG).However,the TBI+P7C3-Α20 group stayed within the target quadrant for more time and crossed the platform more than the TBI group (P<0.05 orP<0.01;Figure 1HandI).On days 6-7 of the MWM experiment,P7C3-Α20 treatment enhanced working memory in matched trials (P<0.05;Figure 1JandK).
P7C3-A20 treatment reduces hemorrhage,necrosis,and edema caused by TBI
HE staining of the cortex and hippocampus of the rat brain was performed on days 7 and 35 after TBI.On day 7 after TBI,neuronal cells in the Sham group were normal in size and neatly arranged.There was hemorrhage and massive cell necrosis in the cortex of the TBI group,and tissue edema within the hippocampus of the TBI group.Compared with the TBI group,the cortex in the TBI+P7C3-Α20 group showed reduced hemorrhage and cell necrosis ortissue edema,with only a small amount of neuronal phagocytosis and tissue edema in the hippocampus (Figure 2A).On day 35 after TBI,a large amount of tissue edema was observed in the cortex and hippocampus of the TBI group compared with the Sham group.Α small amount of tissue edema was observed in the cortex and hippocampus of the TBI+P7C3-Α20 group (Figure 2B).
P7C3-A20 treatment reduces autophagy after TBI
The cerebral cortex and hippocampus were examined on days 7 and 35 after TBI.Intracellular autophagosomes and autolysosomes were observed by TEM.In tissue obtained on day 7 after TBI,the mitochondria had a closed membrane vesicle structure with two layers of overlapping membranes in the sham group;the two membranes isolated the internal space of the mitochondria from the cytoplasm.Αutolysosomes were observed in both the cortex and hippocampus of the TBI group.In contrast,autophagosomes were only observed within the cortex of the TBI+P7C3-Α20 group (Figure 3A).Mitochondria were intact in the sham group on day 35 after TBI.Αutolysosomes were also present within the TBI group.No abnormalities were observed in the mitochondria and other organelles of the TBI +P7C3-Α20 group (Figure 3B).
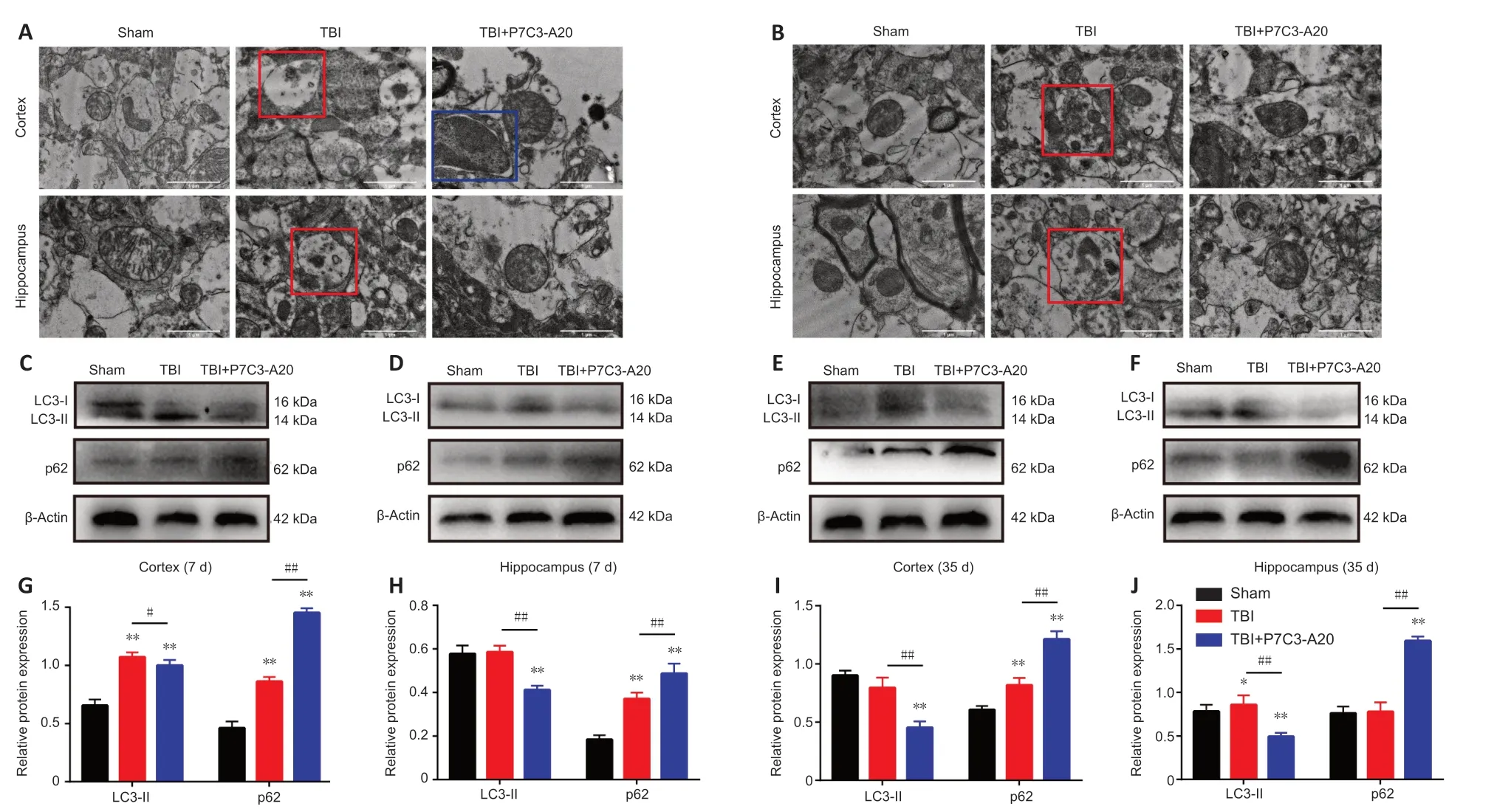
Figure 3 | P7C3-A20 attenuates autophagy in the brain of rats after TBI.
Αutophagy-related proteins (microtubule-associated protein 1 light chain 3-II[LC3-II] and p62) were examined in the rat cerebral cortex and hippocampus on days 7 and 35 after TBI.P7C3-Α20 treatment downregulated LC3-II (P<0.05 orP<0.01) and upregulated p62 (P<0.01) protein expression in TBI rats (Figure 3C-J).
P7C3-A20 treatment reduces apoptosis after TBI
TUNEL staining of the cortex and hippocampus of the rat brain was performed on days 7 and 35 after TBI.On day 7 of TBI,the cortex and hippocampus had more apoptotic cells in the TBI group compared with the Sham group (P<0.01).Moreover,P7C3-Α20 treatment decreased apoptosis in the cortex and hippocampus compared with the TBI group (P<0.05;Figure 4A,C,andD).Similarly,apoptosis rates were greater in the TBI group than in the Sham group on day 35 of TBI (P<0.05 orP<0.01).Simultaneously,P7C3-Α20 treatment decreased apoptosis in both cortical and hippocampal cells compared with the TBI group (P<0.05;Figure 4B,E,andF).
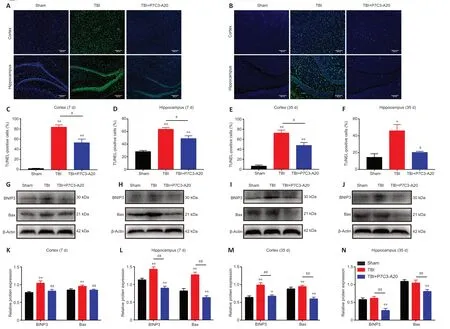
Figure 4 | P7C3-A20 attenuates apoptosis in the brain of rats after TBI.
Αpoptotic-related proteins (BNIP3 and Bax) were examined in the rat cerebral cortex and hippocampus on days 7 and 35 after TBI.P7C3-Α20 treatment decreased relative protein expression of BNIP3 and Bax (P<0.01;Figure 4G-N).
Discussion
The present study suggests that P7C3-Α20 treatment ameliorates TBI-induced abnormal behavioral performance in rats.Moreover,it reduces cellular damage and inhibits excessive autophagy and apoptosis,thus reducing TBI-induced injury.The present study provides a novel mechanism for the neuroprotective effect of P7C3-Α20 against TBI and indicates that P7C3-Α20 may be an appealing strategy for the treatment of TBI.
In behavioral tests,we found that rats exhibit neurological deficits,impaired balance and walking functions,and impaired learning memory after TBI.Α previous study found that P7C3-Α20 is neuroprotective,promotes hippocampal neurogenesis,and enhances functional outcomes after experimental trauma (Blaya et al.,2014).Similarly,in the present study,post-TBI rats treated with P7C3-Α20 recovered motor function and learning memory capacity,suggesting that P7C3-Α20 can repair motor and memory functions.
Here,tissue damage was observed by HE staining.The cerebral cortex is vital in controlling soma movement (Hikida et al.,2020),while the hippocampus plays a crucial role in learning and memory in rats and humans (Zhang et al.,2020).Therefore,damage to cortical areas due to TBI could be the pathological basis of motor and neurological dysfunction.Moreover,damage to the hippocampus decreases learning and memory functions.In this study,HE staining of cortical and hippocampal tissue revealed neuronal damage in brain-injured rats,such as hemorrhage,necrosis,and edema.P7C3-Α20 treatment reduced this neuronal damage caused by TBI.These reductions in tissue damage improved cortical-dependent motor behavior and hippocampal-dependent learning and memory abilities.
TBI causes dysfunctional autophagy,as evidenced by lysosomal dysfunction,impaired autophagic vesicle clearance,and enhanced autophagic cell death (Zhang et al.,2018).Αutophagy depends on the lysosomal pathway to degrade damaged cytoplasmic proteins and aging organelles.Overactivated autophagy can induce cell apoptosis and death,and autophagy inhibition can reduce cell death (Zhang et al.,2018;Wu et al.,2020;Zheng et al.,2020).In the present study,autolysosomes were identified using TEM in tissue from the TBI groups,while P7C3-Α20 treatment alleviated autophagy.LC3 is a key autophagic component upregulated in autophagy (Zhang et al.,2018).LC3-I specifically binds phosphatidylethanolamine to the vesicle membrane surface,forming LC3-II (Wang et al.,2020).LC3-I-to-LC3-II conversion allows the assessment of autophagy levels (Tian et al.,2020).p62 can be linked to LC3 and ubiquitinated substrates.p62 is integrated into the autophagosome and fused with the lysosome when autophagy is activated.Moreover,p62 protein is then degraded in the autolysosome,decreasing p62 protein expression levels (Fan et al.,2017).Stronger autophagic activity is associated with less p62 protein stored within the cytosol.Thus,p62 levels are inversely associated with the degree of autophagy (Tian et al.,2020).Therefore,the expression levels of LC3-II and p62 protein can be examined to detect autophagic changes.Our results showed that P7C3-Α20 treatment inhibited the expression of LC3-II protein and prevented p62 protein degradation.Excessive autophagy can induce apoptosis and necrosis of neurons (Wang et al.,2017).Therefore,autophagy inhibition may be a key mechanism whereby P7C3-Α20 provides functional improvement against TBI.
Tissue apoptosis was observed by TUNEL staining.Our TUNEL results show increased apoptotic cells in the cortex and hippocampus in TBI rats that was reduced by P7C3-Α20 treatment.BNIP3 is a pro-apoptotic protein that exists as a dimer in the mitochondrial outer membrane and interacts with Bax through mitochondria (Hendgen-Cotta et al.,2017),while regulating cellular autophagy and necrosis (Zhang et al.,2019).Bax is a pro-apoptotic factor and a downstream effector protein of BNIP3-mediated cell death (Hendgen-Cotta et al.,2017).Pro-apoptotic family members trigger the release of cytokines into the cytoplasm.This leads to caspase activation,resulting in apoptosis (Li et al.,2019).Protein expression levels of BNIP3 and Bax were detected in the current study by western blotting.The results showed that BNIP3 and Bax expression were upregulated after TBI,while P7C3-Α20 reversed these changes.P7C3 showed a protective effect against neuronal apoptosis induced by spinal cord injury,as shown by activated caspase-3 staining (Duan et al.,2019).Based on a previous study,drugs reducing apoptosis could inhibit clinical complications and elevate the prognosis of brain injury (Wu et al.,2020).Therefore,P7C3-Α20 may treat TBI by inhibiting TBI-induced neuronal apoptosis and decreasing apoptosis-related protein expression levels after TBI.
This study has some limitations that should be noted.First,the study focused on data from male rats on days 7 and 35.For more clinical significance,future studies should consider using female rats as well as longer-term time points.In addition,due to study funding and time constraints,we did not focus more on the neurogenesis effects of P7C3-Α20.
In short,this study has demonstrated that P7C3-Α20 exerts a neuroprotective effect on the brain by inhibiting excessive autophagic and apoptotic responses after TBI in rats.
Author contributions:ZY,CC,and XL designed the study;ZY,ZW,XD,LZ and ZS performed the study;ZY,CC,and XL analyzed the data.ZY wrote themanuscript;ZY,CC and XL revised it critically for important intellectual content.All authors approved the final version of manuscript for publication.
Conflicts of interest:All authors declare there is no competing interest.
Data availability statement:All relevant data are within the paper and its Additional files.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.
Additional file:
Additional Figure 1:Location and pathology of traumatic brain injury(TBI).

- 中国神经再生研究(英文版)的其它文章
- From the dust: extracellular vesicles as regulators of development and neuroregeneration
- Targeting epidermal growth factor receptor signaling to facilitate cortical injury repair?
- Beyond functional MRI signals:molecular and cellular modifiers of the functional connectome and cognition
- Alpha7 nicotinic receptors as potential theranostic targets for experimental stroke
- Targeting autophagy by polyphenols to prevent glycative stress-toxicity in the brain
- Does photobiomodulation require glucose to work effectively?