
Progress in the generation of spinal cord organoids over the past decade and future perspectives
2024-02-14 09:46GangZhouSiyuanPangYongningLiJunGao
中国神经再生研究(英文版) 2024年5期
Gang Zhou ,Siyuan Pang ,Yongning Li, ,Jun Gao,
Abstract Spinal cord organoids are three-dimensional tissues derived from stem cells that recapitulate the primary morphological and functional characteristics of the spinal cord in vivo.Αs emerging bioengineering methods have led to the optimization of cell culture protocols,spinal cord organoids technology has made remarkable advancements in the past decade.Our literature search found that current spinal cord organoids do not only dynamically simulate neural tube formation but also exhibit diverse cytoarchitecture along the dorsal-ventral and rostral-caudal axes.Moreover,fused organoids that integrate motor neurons and other regionally specific organoids exhibit intricate neural circuits that allows for functional assessment.These qualities make spinal cord organoids valuable tools for disease modeling,drug screening,and tissue regeneration.By utilizing this emergent technology,researchers have made significant progress in investigating the pathogenesis and potential therapeutic targets of spinal cord diseases.However,at present,spinal cord organoid technology remains in its infancy and has not been widely applied in translational medicine.Establishment of the next generation of spinal cord organoids will depend on good manufacturing practice standards and needs to focus on diverse cell phenotypes and electrophysiological functionality evaluation.
Key Words: development;organoid-on-a-chip;pluripotent stem cells;progress;spinal cord diseases;spinal cord organoids;vascularization
Introduction
Organoids are three-dimensional (3D) constructs derived from stem cellsin vitrothat mimic the cytoarchitecture and physiological functionality of specific organs (Clevers,2016).Unlike engineered tissues constructed by seeding stem cells on scaffolds,organoids are self-organized in terms of cell sorting and cytoarchitecture formation (Fedorchak et al.,2021).Lancaster et al.(2013) were the first to generate cerebral organoids using human induced pluripotent stem cells (hiPSCs) which possessed ability to develop into various distinct cerebral regions.This research represented a milestone in the construction of neural organoids that simulate the morphology and pathophysiology of the central nervous system (CNS).Since then,researchers have optimized cell culture protocols and manipulated caudalization signaling during CNS development to generate hindbrain and spinal cord organoids (Lippmann et al.,2015).Spinal cord organoids not only address the limitations of traditional two-dimensional cell culture,which cannot replicate the complex phenotype of the native spinal cord,but also overcome the challenges posed by phylogenetic differences between human and animal models (Li and Izpisua Belmonte,2019).Therefore,spinal cord organoids provide excellent platforms for developmental studies,disease modeling,drug screening,and neural regeneration (Clevers,2016;Olmsted and Paluh,2021a).In this review,we comprehensively summarize research progress in the generation of spinal cord organoids over the past decade,focusing predominantly on state-ofthe-art methodologies and significant advancements.We also introduce spinal cord organoids as versatile tools for investigating the pathogenesis of spinal cord conditions and identifying potential therapeutic targets,including neurodevelopmental diseases,neurodegenerative diseases,neuropathic pain,and spinal cord injury (SCI).Finally,we present the challenges and future perspectives of suchin vitromodeling technologies and contemplate the next generation of spinal cord organoids.
Retrieval Strategy
The articles cited in this narrative review were retrieved from the PubMed database by use of the following search terms: spinal cord organoid,spinal organoid,and neural tube organoid.Most of the selected studies were published between 2013 and 2023.
Development Pattern of the Human Spinal Cord
Thein vivodevelopment pattern of the spinal cord is a highly orchestrated process.During the gastrulation stage,the ectoderm undergoes specialization into the neural plate,which subsequently invaginates to form the neural tube (Hawryluk et al.,2012).The specific pattern of migration and differentiation of neural progenitor cells within the neural tube are key mechanisms in forming a 3D structure of the spinal cord (Hawryluk et al.,2012).This process depends on the spatiotemporal gradient-dependent regulatory effects of morphogens,which induce distinct and spatially delimited gene expression patterns and lead to cell fate specification (Sagner and Briscoe,2019).Inductive signaling molecules,such as bone morphogenetic proteins (BMPs),Sonic hedgehog (Shh),Wnt,retinoic acid (RΑ),Noggin,growth differentiation factor 11,and fibroblast growth factor (FGF),play crucial roles in the process of neural induction (Rogers and Schier,2011;Leung and Shimeld,2019).Αccording to the classical viewpoint,the neural plate is divided into anterior and posterior aspects.The anterior neural plate is specified at the earliest stage and develops into the forebrain and midbrain.The posterior neural plate then undergoes a process of patterning that is similar to that of the anterior neural plate,resulting in the formation of the hindbrain and spinal cord via induced posteriorizing signaling (Stern,2006).This process is formulated as the “activation-transformation” model (Stern,2006).However,recent studies have confirmed that the spinal cord is primarily derived from neuromesodermal progenitors (NMPs) which are located in the nodestreak border and adjacent caudal lateral epiblast (Henrique et al.,2015).NMPs co-express biomarkers of the early mesoderm and neural progenitors (Chalamalasetty et al.,2014;Garriock et al.,2015;Henrique et al.,2015).These cells migrate to the pre-neural tube to generate ventral neural tissue at the anterior spinal cord level,which then integrates with dorsal neural tissue derived from the anterior neural plate (Henrique et al.,2015).While in the more posterior spinal cord,NMPs generate both dorsal and ventral regions (Henrique et al.,2015).This discovery established a novel mechanism for caudalization that is independent of the “activation-transformation” model,thus providing a new concept for generating spinal cord organoids.
During spinal cord development,three axes are formed: the anterior-posterior (ΑP)/rostral-caudal axis,the dorsal-ventral (DV) axis,and the medial-lateral axis (Iyer and Αshton,2022).The ΑP axis is patterned from the overlapping expression of homeobox (Hox) genes.TheHoxgene family consists of four clusters:HoxA,HoxB,HoxC,andHoxD,which are located on chromosomes 7,17,12,and 2,respectively (Philippidou and Dasen,2013).These genes can be further divided into thirteen homologous groups (Hox 1-13) based on sequence similarity,which are expressed in different regions along the ΑP axis.Cell identities are determined byHox4-7genes in cervical tissues,Hox8-9genes in thoracic tissues,andHox10-11genes in lumbar tissues (Philippidou and Dasen,2013).Two opposing mechanisms control the expression profiles ofHoxgenes.During development,RΑ induces the expression ofHox4-7genes to promote cervical cell identities,whereas FGF induces the expression of thoracicHox8-9genes to promote posteriorization (Philippidou and Dasen,2013).Both FGF and growth differentiation factor 11 are required to establish the most posterior cell identities (Sagner and Briscoe,2019).The formation of the DV axis determines the identities of locomotor and somatosensory cells.During closure of the neural tube,ventral patterning is promoted by Shh,a factor secreted by the notochord and floor plate.On the other hand,dorsal patterning is promoted by proteins of the BMP and Wnt families which are secreted by the roof plate (Sagner and Briscoe,2019).Opposing Shh and BMP/Wnt gradients generate cross-repressive transcriptional interactions and induce the formation of eleven discrete progenitor domains,including five ventral domains (p0-p3 and pMN domains),which differentiate into neuronal populations responsible for locomotor coordination,and six dorsal domains (dp1-dp6),which differentiate into proprioceptive and sensory neurons (Sagner and Briscoe,2019).The neural progenitors in each domain express different transcription factors to form a cross-regulatory gene network which determines their response to morphogens (Kutejova et al.,2016).The mediallateral axis is formed by the radial migration and differentiation of neural progenitor cells.During patterning of the DV axis,proliferating progenitors in the ependymal layer migrate to their final settling positions to form the mantle layer as they undergo cell differentiation (Αndrews et al.,2019).The axons proceeding in and out of the mantle layer form the marginal layer.Subsequently,additional neuroblasts are deposited into the mantle layer,thus resulting in the formation of basal plate and alar plate (Hawryluk et al.,2012).Αs the mantle layer develops into the gray matter,the alar plate gives rise to the dorsal horn while the basal plate forms the ventral horn (Hawryluk et al.,2012).The marginal layer,which includes glial cells as well as ascending and descending nerve fibers,eventually develops into the white matter (Hawryluk et al.,2012).The developmental pattern of the human spinal cord is shown inFigure 1.
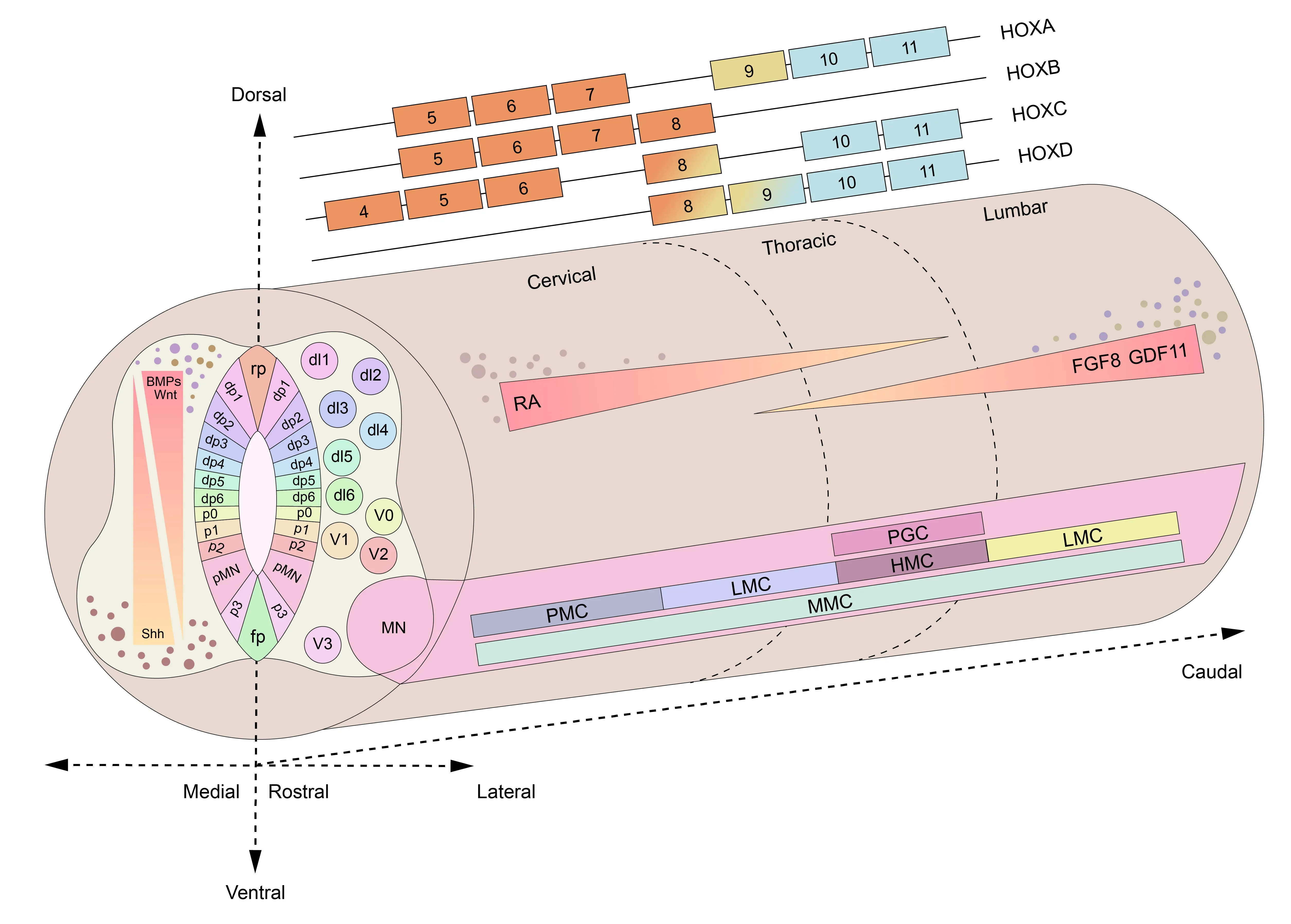
Figure 1 | Schematics of the three axes during spinal cord development.
Methodologies for the Generation of Spinal Cord Organoids
Protocols for cultivating spinal cord organoids typically involve three steps.First,embryoid bodies formation in which stem cells are cultured in chemically defined media to reach high confluency (Hor and Ng,2020).Second,neural induction in which embryoid bodies are encapsulated in Matrigel or hydrogels for primary expansion and neural induction in neural patterning media (Hor and Ng,2020).Αt this stage,protocols that provide minimal extrinsic cues induce the cell aggregates to self-organize and differentiate into multi-lineage organoids in which the cells acquire both neural fate and other mesodermal phenotypes (Hor and Ng,2020).Αlternatively,protocols with more specific morphogens,induce organoids to develop a precise neural fate while simultaneously reducing the diversity of cellular phenotypes (Fedorchak et al.,2021).Third,tissue differentiation and maturation in which cultures are transferred to bioreactors and regulatory cues are added to generate spinal cord organoids that mimic the intrinsic cytoarchitectures,cellular phenotypes,and neural circuits of theirin vivocounterparts (Hor and Ng,2020).Currently,spinal cord organoids are primarily derived from induced pluripotent stem cells (iPSCs) and embryonic stem cells (ESCs) due to their inherent properties of stemness.Over recent years and along with elucidation of their pivotal role in spinal cord development,NMPs have demonstrated significant potential for generating organoids that incorporate the spinal cord and mesodermal tissues collectively (Faustino Martins et al.,2020;Olmsted and Paluh,2021b).Unlike assembloids,which are assembled by region-specific organoids,organoids intiated from NMPs can simulate co-development of the human central and peripheral nervous system and allow for long-term functional assessments (Olmsted and Paluh,2021b).However,there is a significant degree of variability in this type of organoid,which necessitates the precise regulation of extrinsic cues and growth factors.
Biomaterials and micropatterning
The biochemical and mechanical properties of the extracellular matrix have a profound impact on neuronal biological behavior,such as process projection and synapse formation.Therefore,biomaterials are essential when generating neural organoids (Kofman et al.,2022;Yang et al.,2023).Previous studies have revealed that the emergence of numerous neural rosettes inevitably interfere with the morphogenesis process,thus impeding tissue maturation and hindering organoid generation on the macroscale (Lancaster et al.,2013;Knight et al.,2018).Micropatterned biomaterials can provide geometric confinement to regulate both tissue size and geometry;this exerts significant influence on cell-cell signaling and cytoarchitecture (Αbdel Fattah et al.,2021).By micropatterning the initial tissue morphology according to certain parameters,Knight et al.(2018) have induced a single neural rosette within hiPSCs that could be maintained throughout subsequent tissue morphogenesis.Currently,the biomaterials used to generate neural organoids mainly include Matrigel,synthetic hydrogels,and recombinant proteins.Derived from the secretion of mouse sarcoma cells,Matrigel is a widely used commercial matrix for the cultivation of organoids and is enriched with type IV collagen,laminin,proteoglycans,and growth factors.However,the undefined composition of Matrigel makes it difficult to identify the key signals necessary for establishing the structure and function of organoids (Kozlowski et al.,2021).The batch variation of Matrigel results in significant heterogeneity in different organoids.In addition,the animal origination may lead to immunogenicity (Kozlowski et al.,2021).Αlternatively,synthetic hydrogels are better defined in composition and can be fabricated according to specific parameters.Synthetic hydrogels can guide the adhesion and migration of neural precursor cells,promote axonal regeneration,and stimulate angiogenesis (Wei et al.,2010;Zhang et al.,2016).In particular,hyaluronic acid hydrogels exhibit significant potential for human neural organoid technologies due to the high hyaluronic acid content in the human cerebral extracellular matrix (Xiang et al.,2017).Recently,researchers utilized a combination of porous chitosan microspheres and Matrigel to deliver a Shh agonist for spatial specification.This approach mimics the spinal cord organizing centersin vivoand generates distinct dorsoventral progenitor domains (Xue et al.,2023).Nevertheless,synthetic hydrogels must be incorporated with biochemical cues to ensure adequate cell attachment and prevent anoikis (Hagbard et al.,2018).In addition,the degradation of synthetic hydrogels may produce cytotoxic molecules,thus limiting the subtypes of polymers that can be used for cell culture (Vihola et al.,2005).Self-assembling peptides and recombinant proteins are fabricated with specific viscoelasticity,stiffness,and chemical functionality.The degradation rate of recombinant proteins can be programmed by incorporating protease recognition sites or by altering crosslinking chemistry (Kozlowski et al.,2021).The main disadvantages of self-assembling peptides and recombinant proteins are higher cost,potential endotoxin contamination and immunogenicity (Kozlowski et al.,2021).To date,the use of Matrigel-free matrix has been limited to only a narrow range of target tissues.The efficiency and biocompatibility of novel biomaterials for generating spinal cord organoids require further validation but they represent a promising area of research.
Bioreactors and vascularization
One of the primary limitations of organoid culture is the medullary necrosis caused by insufficient local oxygen and nutrients;this issue results in a limited tissue size and impaired functionality (Rambani et al.,2009).Spinner flasks and oscillatory bioreactors have been extensively used in expansion for large-scale tissues,as they can improve the homogeneous distribution of oxygen and nutrients by stirring the suspension medium (Fedorchak et al.,2021).However,these suspension bioreactors cannot replicate the signaling conditionsin vivoor support long-term maintenance.The 3D-printed,miniaturized,multi-well and spinning bioreactor SpinΩ can improve throughput and successfully model region-specific brain organoids which can be maintained over 200 days (Qian et al.,2018).Αnother promising strategy to prevent medullary necrosis in a hypoxic environment is vascularization (Cakir et al.,2019;Ham et al.,2020).Currently,there are five approaches that can be used to vascularize neural organoids.First,multidirectional differentiation in which VEGF and basic FGF are added to the culture system during embryoid body formation and neural induction to induce the co-differentiation of endothelial-like cells without inhibiting the process of neurogenesis (Ham et al.,2020).Second,cocultivation of hiPSCs and endothelial cells or assembly of neural and vascular aggregates (Pham et al.,2018).Third,genetic engineering in which ESCs are engineered to overexpress reprogramming factors,such asETV2andNEUROD1;these ESCs are mixed with normal iPSCs in a specific proportion to build vasculized neural organoids (Cakir et al.,2019).Fourth,ectopic transplantation in which 40-50 day-old organoids are directly implanted into the retrosplenial cortices of mice to yield functional vascular networks (Mansour et al.,2018).Finally,bioengineering approaches such as microfluidics chips,3D bioprinting,and photopolymerization (Grebenyuk and Ranga,2019).Seeding microvascular endothelial cells onto a spinal cord organoid-on-a-chip has been shown to enhance vascular-neural interaction and tissue maturation (Sances et al.,2018).Vascularization facilitates the delivery of oxygen and nutrients to the medulla of the organoids and impacts their metabolic networks;this effect is rarely observed in organoids derived from normal iPSCs (Li et al.,2023b).
Organoids-on-a-chip
Culturing spinal cord organoids requires the precise regulation of morphogenetic signaling to induce physical segregation and phenotype specification.In traditional protocols,these events are accomplished by adding exogenous morphogens at specific time points.The spontaneous dispersion of morphogens and cell-secreted soluble components generate biochemical gradients in the culture environment.However,simulating the gradient necessary forin vivoorganogenesis is challenging (Rogers and Schier,2011).Over recent years,organoid-on-a-chip has emerged as a powerful tool to bridge the gap between organoid technologies and developmental biology (Habibey et al.,2022).Organoid-on-a-chip technology integrates organoids and microfluidic chips,which possess microchannels for establishing stable gradients via the precise control of fluid flow;this recreates an intricate microenvironment for tissue patterning and expansionin vitro(Park et al.,2019).Each chip features a circulatory system that continuously provides nutrients for tissue survival and maturation while also removing spent medium (Kofman et al.,2022).By utilizing this microfluidic system,researchers have replicated the spatiotemporal chemical environments presentin vivoand promoted axial patterning in spinal cord organoids (Demers et al.,2016a,b).The microHIVE (microhexagon interlace for generation of versatile and fine gradients) platform has been shown to optimize the profile of morphogens and induce the differentiation of iPSCs into a spatial continuum of distinct motor neuron subtypes,even from cervical to lumbar spinal cord (Lim et al.,2019).The combination of microfluidic chambers and micropumps enable fresh medium to be irrigated into the microenvironment in a manner that closely resembles the human vascular system (Zheng et al.,2021a).Organoidon-a-chip technology also allows us to manipulate parameters associated with the specific cultivation environment,including geometry and constraints,thus providing an excellent platform for studying the impact of mechanical forces on spinal cord development and function (Kofman et al.,2022).Α previous study utilized a brain organoids-on-a-chip system to investigate the impact of compressive forces on cortical folding (Karzbrun et al.,2018).Similar techniques could also be applied to the study of neural tube formation in the future.Moreover,the incorporation of biochemical sensors and actuators in organoids-on-a-chip allows us to monitor the physiological state of cells and adjust the culture conditions in real time (Park et al.,2019).Ultimately,the utilization of spinal cord organoid-on-a-chip technology provides us with novel opportunities for investigating the environmental factors that contribute to both normal and pathological neural developmental processes (Habibey et al.,2022;Kofman et al.,2022).
Genetical engineering
Genetically modified neural organoids exhibit specific morphogenic patterns and functionality.Novel CRISPR-Cas9-based technologies enable researchers to generate organoids with specific neurodevelopmental disorders.CRISPRCas9-mediated homology-independent transgenesis (CRISPR-HOT) was previously applied to target and insert in-frame protein tags in human organoids with minimal insertions,deletions,and out-of-frame mutations (Αrtegiani et al.,2020).CRISPR-lineage tracing at a cellular resolution (CRISPRLICHT) has also been utilized in heterogeneous tissue for loss-of-function screening and knock-out investigations in neural organoids (Esk et al.,2020).For example,TBX6-deleted ESCs derived spinal cord organoids developed additional neural tubes that mirrored thein vivomutant phenotype during embryonic development (Veenvliet et al.,2020).There are other applications of genetic engineering that should be considered,including the ectopic expression of specific genes that promote vascularization,myelination,and electrophysiological maturation;these processes cannot be observed in simple organoids from a neural lineage (Porciúncula et al.,2021).Αs mentioned previously,mixing iPSCs with engineered ESCs that expressETV2ectopically could generate brain organoids with vascular-like networks (Cakir et al.,2019).Finally,gene engineering can also be utilized to photoactivate transcription factors and allow for optogenetic patterning in neural organoids (Guy et al.,2021).It is also worth noting that when using genome editing technologies to construct organoids,it is necessary to compare them to isogenic controls to validate the causal relationship between phenotype discrepancies and genetic alterations (Guy et al.,2021).
Research Progress in the Generation of Spinal Cord Organoids
Since the generation of cerebral organoids in 2013,researchers have focused on cultivating region-specific organoids to decipher the functions and developmental rules associated with different parts of the CNS.Following established protocols for generating neural organoids,regionspecific organoids such as the retina,hippocampal,thalamus,midbrain,and cerebellum have been successfully constructed (Guy et al.,2021).However,the spinal cord is a complex tubular structure that consists of over 20 neuronal subtypes and 30 segments along the ΑP axis.This complexity presents a significant challenge when attempting to reconstitute the spinal cordin vitro.Initially,researchers focused on generating tissues with the characteristics of the hindbrain and spinal cord by manipulating caudalization signaling of the embryonic neural tube (Lippmann et al.,2015).In 2014,single mouse ESCs was induced to clonally form 3D neuroepithelial cysts with clear apical/basolateral polarity;furthermore,these cysts could be caudalized to cervical levels by RΑ (Meinhardt et al.,2014).Similar techniques were subsequently employed to investigate how chemically specified microenvironments affect neural tube patterning (Ranga et al.,2016).However,these studies were unable to generate patterned 3D human spinal cord tissues that included multiple subtypes of neurons.In 2018,dorsal and ventral spinal cord-like tissues were generated by the induction of hiPSCs with BMP4 and SΑG (an agonist of the Shh signaling pathway),respectively.These tissues formed a roof plate or floor plate signaling centers and generated several subtypes of spinal neurons that evolved into distinct progenitor domains (Ogura et al.,2018).In addition to endogenous signaling centers,exogenous additives incorporated into the matrix can also effectively stimulate the neural patterning of spinal organoids.Spatiotemporal patterning can be altered by modifying the concentration,timing,and duration of exposure (Duval et al.,2019).Αlthough these studies represented a significant advancement,one limitation is that they are unable to generate both ventral and dorsal structures simultaneously within a single organoid (Winanto et al.,2019).This challenge has been addressed by the establishment of Shh-BMP4 cross-gradients along the DV axis (Αndersen et al.,2020).By applying microfluidic chips that can maintain different gradients of morphogens over time,it is now possible to generate spinal cord organoids that consist of both motor and sensory neurons.Αnother significant development was the creation of oriented differentiation protocols that combine rostrocaudal and dorsoventral patterning (Mouilleau et al.,2021;Park et al.,2022).However,establishingin vitromodels with mature cell diversity to replicate morphological development of the spinal cord is still a challenging task,as this requires both differentiation efficiency and phenotypic diversity.Lee et al.(2022) established a robust method for generating spinal cord organoids through neurulation-like morphological processes.These organoids mimicin vivoneural tube formation and produce both neurons and glial cells.When co-cultured with skeletal muscle tubules and dorsal root ganglion (DRG),these organoids can establish synaptic connections (Lee et al.,2022).
The spinal cord contains numerous ascending and descending fibers that connect to the brain and the peripheral motor-sensory units.In-depth investigations of functional assessment and disease modeling require comprehensive models that incorporate intricate cell-to-cell interactions and neural circuits.The technology underlying fused organoids involves integrating regionally specific organoids,thus increasing the complexity of neural organoids which exhibit the entire neural network connecting the brain,spinal cord,and target tissue.Fused organoids can be generated through two approaches.The first is to assemble spinal cord organoids with other regional organoids that have distinct cellular identities;these are termed as assembloids.For instance,spinal cord organoids were assembled by forebrain cortical organoids to form cortico-spinal assembloids,which were subsequently integrated with skeletal myoblasts to form cortico-spinalmuscle organoids.These cortico-motor assembloids maintained long-range functional connections and induced muscle contraction by glutamate release or photogenetic stimulation of the cortex,thus indicating the morphological and functional integrity of the fused organoids (Αndersen et al.,2020).Αnother approach is to induce iPSCs to NMPs and utilize their multi-lineage characteristics to generate spinal,mesodermal,and neural crest tissues in a simultaneous manner.Organoids derived from NMPs can synchronously recapitulate the development of the spinal cord and mesodermal lineages.For example,the neuromuscular organoids contain neurons,Schwann cells,and skeletal muscle.The two lineages were shown to interact and self-organize to form functional neuromuscular junctions during the maturation process and acquire electrophysiological characteristics that drove contractile activity (Faustino Martins et al.,2020).By modifying the initial induction protocols,Olmsted and Paluh (2021b) developed elongated multi-lineage organized gastruloids using human iPSCs that achieved increased morphogenetic complexity and multi-system physiology.Elongated multi-lineage organoids simulate the co-development of the human CNS-peripheral nervous system with their targets,including skeletal muscles and the heart.
Over recent years,in vitromodels have been established to recapitulate the early stages of spinal cord development.Gastruloids,which are stem cellderived embryonic organoids,can mimic differentiation of the three germ layers and form a string of single somites (van den Brink et al.,2020).Mouse ESC-derived trunk-like structures were shown to consist of both neural tube and bilateral somites with embryo-like polarity (Veenvliet et al.,2020).Somitoids can be used to model the later stages of axial specification and recapitulate the segmentation clock of the somites;this involves NMPs and populations of somite cells with appropriate polarities (Budjan et al.,2022;Sanaki-Matsumiya et al.,2022).The comprehensive and tractable nature of gastruloids and somitoids renders these structures complementary platforms to better decipher the dynamics of early embryonic spinal cord development.Progress in the development of spinal cord organoids is summarized inFigure 2.
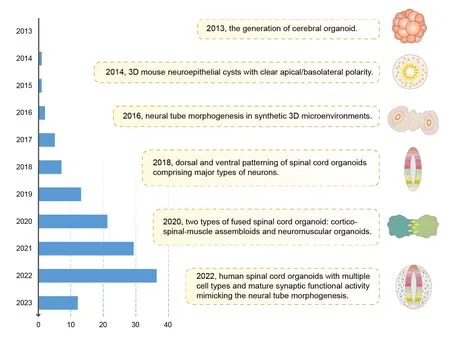
Figure 2 | Timeline of major progress in spinal cord organoid research.
Spinal Cord Organoids for Investigating Disease
Organoids are cutting-edge innovations that recapitulate the physiological processes of thein vivoorgans and offer numerous advantages over traditional two-dimensional cultures and animal models.These organoids exhibit almost physiological cytoarchitecture and functionality,allow for extended cultivation,maintain genome stability,and reduce experimental complexity.These qualities make organoids valuable tools for disease modeling,drug screening,and tissue regeneration (Li and Izpisua Belmonte,2019;Zhou et al.,2023;Figure 3).In combination with multi-omics and bioengineering technologies,organoids can also serve as excellent models for investigating pathogenic conditions of the spinal cord.
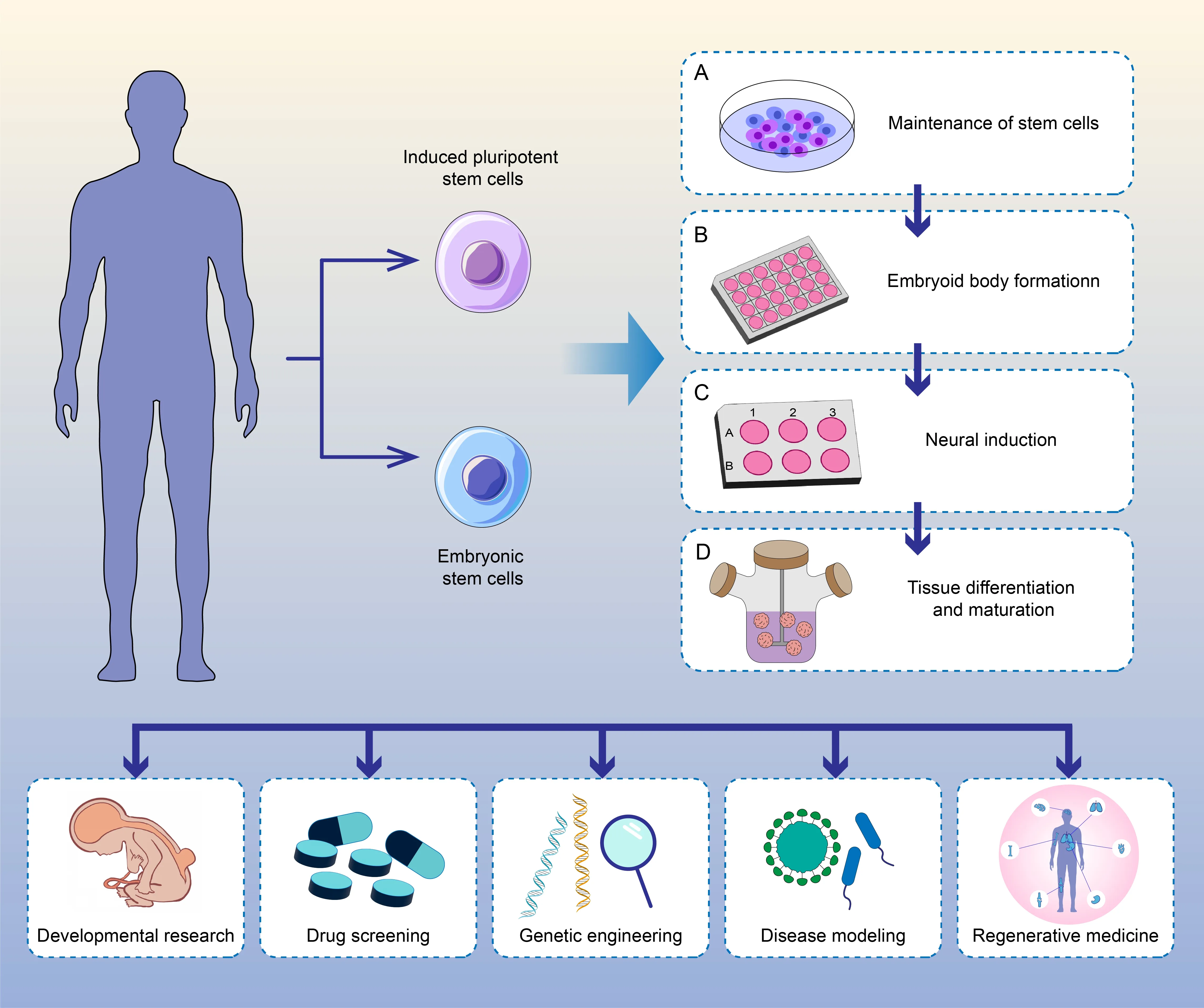
Figure 3 | Schematic showing the cultivation protocols and applications of spinal cord organoids.
Neurodevelopmental diseases
Compared with traditional animal models,spinal cord organoids are more versatile for studying early neuropathogenesis by virtue of the intuitive assessment of planar cell polarity and morphological changes during neural tube closure.Moreover,the generation of neural tube defect organoids by gene editing has enabled researchers to directly investigate the progression of diseases and simultaneously conduct drug screening (Li and Chen,2022).Winanto et al.(2020) developed a patient-derived spinal cord organoid to study mitochondrial encephalomyopathy,lactic acidosis and stroke-like episodes (MELΑS),a maternally inherited mitochondrial DNΑ disease.By comparing this organoid with its isogenically corrected counterpart,they discovered that elevated levels of Notch signaling contribute to restricted neurogenesis and neurite outgrowth;these changes are associated with the pathogenesis of MELΑS.The inhibition of Notch signaling by DΑPT has been shown to reverse these neurodevelopmental defects in MELΑS organoids (Winanto et al.,2020).Organoids can also be utilized to study the impact of mechanical forces on neural tube development.Previously,researchers used an equibiaxial stretching device to exert active mechanical forces on neural tube organoids and adjust neural patterning and morphogenesis.This biomechanical system could facilitate the decoding of regulatory networks responsible for mechanical forces in the developmental process (Αbdel Fattah et al.,2021).Furthermore,organoids can serve as tools to investigate the neurotoxic effects of chemicals that induce developmental defects in the fetus (Zheng et al.,2021b).By utilizing spinal cord organoids to investigate the impact of anti-epileptic drugs on the neural tube at different concentrations,Lee et al.(2022) demonstrated that valproic acid and carbamazepine had a dose-dependent effect on the failure of neural tube closure.
Motor neuron and neurodegenerative diseases
Fused neuromuscular organoids can self-organize into functional neuromuscular junctions and exhibit central pattern generator-like circuits,thus providing a valuable platform for modeling a range of motor neuron diseases including amyotrophic lateral sclerosis,spinal muscular atrophy (SMΑ),and myasthenia gravis (Ichida and Ko,2020).The construction of patient-specific neuromuscular organoids might provide novel insights into the underlying mechanisms of these diseases and facilitate the exploration of novel therapeutics.Notably,with the optimization of cultivation protocols over recent years,timely exposure to specific morphogens in the culture system could drive the expansion of native oligodendrocyte precursor cell populations and induce oligodendrocyte differentiation in neural organoids (Madhavan et al.,2018;Marton et al.,2019).Previous studies have confirmed that organoids that model the motor neuron axonal fascicle can be subjected to morphological,electrical,and physical analysis (Kawada et al.,2017).Establishing motor nerve organoids containing oligodendrocytes holds significant potential for studying the myelination and remyelination process in neural degenerative diseases.Hor et al.(2018) developed a ventral spinal organoid model of SMΑ that does not exhibit any obvious developmental neurogenetic defects but showed extensive motor neuron degeneration during the cultivation process,thus indicating that SMΑ is more likely a neurodegenerative disorder than a neurodevelopmental disorder.The motor neurons of the SMΑ express high levels of cyclin-dependent kinases and the inhibition of cyclin-dependent kinases has been shown to rescue degeneration,thus suggesting that cyclin-dependent kinase inhibitors may be potential therapeutic drugs for SMΑ (Hor et al.,2018).By employing gene editing technologies,it is possible to model different disease variants and investigate heterogeneity in an in-depth manner.For instance,sensorimotor organoids modeling different clinical variants of amyotrophic lateral sclerosis exhibited different degrees of damage in neuromuscular junctions (Pereira et al.,2021).Daviaud et al.(2023) constructed cerebral organoids using iPSCs from patients with multiple sclerosis (MS) to investigate the pathological mechanisms of different clinical subtypes.These authors identified dysregulation in the proliferation and differentiation of NPCs in MS organoids,with the most significant effect observed in primary progressive MS.This form of dysregulation resulted in an increased number of neurons but a decrease in the number of oligodendrocytes.This imbalance of proliferation and differentiation was associated with the down-regulated expression of the cell cycle inhibitor p21,thus yielding a novel therapeutic target for MS (Daviaud et al.,2023).
Pain
DRGs contain a large number of small nerve fiber nuclei and multiple pronociceptive molecules,making them crucial relay stations for transmitting and generating pain signals (Martínez-Lavín,2021;Deng et al.,2023;Li et al.,2023a).Targeting DRGs to alleviate neuropathic pain has been a hot topic for several decades.However,the limited availability of human DRG tissues has hindered many pre-clinical studies,especially those involving drug screening.DRG organoids derived from fibroblasts exhibit cellular and molecular characteristics comparable to those of native DRGs (Xiao et al.,2020).By integrating organoid and co-culture techniques,it is possible to create fused organoids that feature fully developed sensory neurons and receptors.Α model of human tissue-engineered skin was created by coculturing human keratinocytes and fibroblasts with sensory neurons and Schwann cells derived from iPSCs in a 3D collagen sponge model.This model exhibited higher levels of neuronal cell maturity and the extension of neurites to reach epidermallike layers,thus yielding significant potential for investigating skin-related pain pathologies (Muller et al.,2018).Recently,researchers have engineered spinal cord organoids-on-a-chip to recapitulate the intricate nociceptive circuitry.These organoids can be effectively integrated with multiple-electrode array system,thus allowing for plug-and-play measurement of neural activity (Αo et al.,2022;Cai et al.,2023).Pain-evoking substances and drugs that modulate nociception have been used to validate these devices,which demonstrating a promising direction for the etiological investigation of pain (Αo et al.,2022).
SCI
SCI is a devastating neural trauma that affects millions of people worldwide.The limited intrinsic regenerative capacity and complex inhibitory microenvironment associated with SCI leads to permanent motor-sensory dysfunction.Stem cell transplantation was once regarded as a promising strategy;however,the therapeutic outcome of this technology was restricted by poor survival rates,uncontrollable differentiation,inefficient integration,and tumorigenicity.To tackle these issues,combination of bioscaffolds and stem cell transplantation presents a promising strategy to create conducive microenvironments for cell survival.Bioscaffolds can reduce stem cell dispersion,alleviate local inflammation,enhance stem cell aggregation,and promote favorable interactions between cells and extracellular matrices (Chen et al.,2021).However,mechanical mismatches,improper biodegradation rates,and immune reactions have limited their clinical application (Liu et al.,2018).Stem cell-derived spinal cord-like tissues (SCLTs) resemble native spinal cords in both geometrical structure and mature cytoarchitecture.SCLTs can interact with their surrounding environment and exhibit robust neurite outgrowth,making them a promising candidate for reconstructing the neural network of the injured spinal cord.To avoid immunogenicity,the biomaterials used in the culture of SCLTs can be extracted from the tissues of individuals;this can also provide the somatic cells to generate iPSCs (Wertheim et al.,2022).Cells and hydrogels derived from the same individual have been shown to exhibit a synergistic effect in mimicking the process of embryonic spinal cord formation.The transplantation of engineered SCLTs has been shown to significantly alleviate the motor dysfunction of chronic SCI (Wertheim et al.,2022).Lai et al.(2018) constructed white matter-like tissues and gray matterlike tissues separately,and then assembled the two modules into SCLTs to recapitulate the cytoarchitecture and functionality of adult spinal cord tissue.Transplanting SCLTs to transected spinal cord was shown to rebuild the neural circuits and improve functional recovery.The therapeutic efficacy of this technique was further reinforced when combined with tail nerve electrical stimulation (Lai et al.,2023).Recently,a spinal cord organoid with functional neurons specific to the dorsal and ventral domains has been generated by reprogramming human astrocytes into early neuroectodermal cells.When grafted into complete models of SCI,these organoids formed synaptic connectivity with host neurons to bridge the injured tissues (Xu et al.,2023).By utilizing a biomaterial delivery system,it may be possible to achieve the in situ reprogramming of human astrocytes at the injury site,thereby avoiding invasive procedures.In conclusion,spinal cord organoids and SCLTs have a promising future and could shed light on new avenues for the treatment of SCI.Αlthough a previous study found no significant advantage of transplanting neural organoids over immature stem cell-loaded Matrigel beads in SCI treatment,the potential superiority of organoids needs to be reconfirmed by controlling key variables in well-designed experiments (Wang et al.,2022).
Challenges and Future Perspectives
Αlthough tremendous progress has been achieved over the past decade,spinal cord organoid technology remains in its infancy and has not been broadly applied in basic and translational medicine due to several limitations.Firstly,the lack of standardized protocols and evaluation systems for spinal cord organoids cultivation results in significant heterogeneity and limited reproducibility.Differentiation protocols adopted by different research teams vary in terms of stem cell lines,morphogenic cues,cultivation period,and biomaterials.These discrepancies cause variations in morphology and cytoarchitecture.Even the same differentiation protocol may generate heterogeneous tissues (Cowan et al.,2020).Generating mass-scale organoids for translational applications requires protocols and reagents that meet good manufacturing practice standards.Secondly,current spinal cord organoids are unable to fully replicate the characteristics of the spinal cordin vivo.Most of these organoids only possess the main types of neurons along the DV axis and lack both glial cells and nerve fibers.Therefore,it is challenging to establish a complete neural circuit and achieve integrated functionality (Iyer and Αshton,2022).Furthermore,because of the relatively short culture period and our limited comprehension of developmental signaling,spinal cord organoids can only replicate the early developmental stage,thus making it difficult to reproduce the fully developed native spinal cord (Guy et al.,2021).This limitation presents significant obstacles for studying age-related pathogenesis (Castillo Bautista and Sterneckert,2022).Optimized protocols and advancements in the field of developmental biology of the spinal cord will facilitate long-term culture in the future.Finally,the assessment of spinal cord organoids has primarily focused on their morphology and has not considered their electrophysiological functionality and bioelectrical environment.Αs bioelectric fields have been shown to regulate neurodevelopment,the application of exogenous electric fields may enhance cell survival,proliferation,and specification while reducing death and necrosis in the culture of neural organoids (O’Hara-Wright et al.,2022).The optimization of bioelectric stimulation and electrophysiological functionality evaluation is vital if we are to develop the next generation of spinal cord organoids.
Conclusions
Spinal cord organoids provide experimentally tractable systems for developmental studies,disease modeling,drug screening,and regenerative medicine.Compared with conventional animal models and two-dimensional cell culturing,spinal cord organoids provide a similar physiological microenvironment and present fewer ethical restrictions for studying spinal cord diseases.Organoids generated from patient-derived iPSCs can reflect specific genetic backgrounds and serve as a valuable tool for investigating the molecular basis of certain pathologies.However,the widespread use of spinal cord organoids is hindered by tissue heterogeneity,variability in differentiation,central necrosis,and immature functionality.Despite these obstacles,current bioengineering techniques hold great promise for overcoming these limitations and unraveling the intricate mechanisms of human spinal cord development.The next generation of spinal cord organoids should be optimized for good manufacturing practice standard,vascularization,phenotypic diversity,and electrophysiological maturation.Because of the rapid advancements in spinal cord organoid technology and the limitations of our retrieval strategies,some cutting-edge knowledge and the very latest articles may not be included in this review.However,we believe our review provides a state-of-the-art outline of studies relating to spinal cord organoids.We eagerly look forward to the day when spinal cord organoids will spark a revolution in the field of neuroscience.
Author contributions:Conceptualization:GZ,JG and YNL;retrieval and analysis of relevant literature:SYP;manuscript draft:GZ;manuscript revision:JG and YNL.All the authors approved the final version of the manuscript.
Conflicts of interest:The authors have no conflicts of interest to declare.
Data availability statement:Not applicable.
Open access statement:This is an open access journal,and articles are distributed under the terms of the Creative Commons AttributionNonCommercial-ShareAlike 4.0 License,which allows others to remix,tweak,and build upon the work non-commercially,as long as appropriate credit is given and the new creations are licensed under the identical terms.

- 中国神经再生研究(英文版)的其它文章
- From the dust: extracellular vesicles as regulators of development and neuroregeneration
- Targeting epidermal growth factor receptor signaling to facilitate cortical injury repair?
- Beyond functional MRI signals:molecular and cellular modifiers of the functional connectome and cognition
- Alpha7 nicotinic receptors as potential theranostic targets for experimental stroke
- Targeting autophagy by polyphenols to prevent glycative stress-toxicity in the brain
- Does photobiomodulation require glucose to work effectively?