
干旱胁迫对伊犁绢蒿幼苗生长及叶片解剖结构的影响
2024-02-02 15:57韩喆张永强张浩浩罗文浩陈爱萍
草地学报 2024年1期
韩喆 张永强 张浩浩 罗文浩 陈爱萍


doi:10.11733/j.issn.1007-0435.2024.01.011
引用格式:
韓 喆, 张永强, 张浩浩,等.干旱胁迫对伊犁绢蒿幼苗生长及叶片解剖结构的影响[J].草地学报,2024,32(1):105-112
HAN Zhe, ZHANG Yong-qiang, ZHANG Hao-hao,et al.Effects of Drought Stress on Growth and Leaf Anatomical Structure of Seriphidium transiliense seedlings[J].Acta Agrestia Sinica,2024,32(1):105-112
摘要:为探究干旱胁迫对伊犁绢蒿(Seriphidium transiliense)幼苗生长及叶片解剖结构的影响,本研究采用控水实验,对培养45 d后的幼苗进行持续干旱胁迫处理,测定幼苗的株高、根长、地上、地下生物量以及叶片解剖结构。结果表明:随着干旱胁迫程度的加剧,幼苗地上部分生长逐渐减缓,地下部分生长和根冠比则持续增加。当土壤相对含水量降至6.38%时,株高和地上生物量较对照分别降低了73.83%和73.05%(P<0.01),而根长、地下生物量和根冠比较对照分别增加了6.09%,67.92%和527.49%(P<0.01)。随着干旱胁迫程度的加剧,叶片厚度、上下表皮细胞厚度、维管束直径均呈先增后降趋势,而上下角质层厚度则表现出逐渐增加的趋势,栅栏组织呈逐渐下降趋势。综上所述,伊犁绢蒿幼苗通过调节地上部分和地下部分的生长以及叶片解剖结构的变化来应对干旱胁迫。
关键词:干旱胁迫;伊犁绢蒿幼苗;生长;叶片解剖结构
中图分类号:S548 文献标识码:A 文章编号:1007-0435(2024)01-0105-08
Effects of Drought Stress on Growth and Leaf Anatomical Structure of
Seriphidium transiliense seedlings
HAN Zhe1,2,3, ZHANG Yong-qiang1,2,3, ZHANG Hao-hao1,2,3, LUO Wen-hao1,2,3, CHEN Ai-ping1,2,3*
(1. College of Grassland Science, Xinjiang Agricultural University, Urumqi, Xinjiang 830052 China; 2. Ministry of Education
Key Laboratory for Western Arid Region Grassland Resources and Ecology, Urumqi, Xinjiang 830052 China; 3. Xinjiang Key
Laboratory of Grassland Resources and Ecology, Urumqi, Xinjiang 830052, China)
Abstract:In order to explore the effects of drought stress on growth and leaf anatomical structure of Seriphidium transiliense seedlings,the 45 d old seedlings were treated with persistent drought stress using the potting method. The plant height,root length,aboveground and underground biomass and leaf anatomical structure were measured. The results showed that with the increased of drought stress,the aboveground growth of seedlings gradually slowed down,while underground growth and the root to shoot ratio continuously increased. When the relative soil water content decreased to 6.38%,plant height and above ground biomass decreased by 73.83% and 73.05%,respectively,compared with control (P<0.01),while root length,underground biomass,and the root to shoot ratio increased by 6.09%,67.92%,and 527.49%,respectively,compared with control (P<0.01). With the increased drought stress,leaf thickness,upper and lower epidermis cell thickness and vascular bundle diameter first increased and then decreased,upper and lower cuticle thickness increased continuously,and the palisade tissue decreased continuously. In summary,S.transiliense seedlings responded to drought stress by regulating the relative growth of aboveground and underground as well as changing leaf anatomical structure.
Key words:Drought stress;Seriphidium transiliense seedlings;Growth;Leaf anatomical structure
收稿日期:2023-08-23;修回日期:2023-11-09
基金项目:干旱胁迫下伊犁绢蒿幼苗形态、生理响应及分子机制(2021D01A94)资助
作者简介:
韩喆(1996-),女,汉族,吉林农安人,硕士研究生,主要从事饲草学研究,E-mail:810659960@qq.com;*通信作者Author for correspondence,E-mail:xjauchenaiping@sina. com
受全球气候与环境变化影响,持续高温及重大干旱等极端气候事件的发生频率和发展强度都表现出逐年上升趋势[1]。目前全球干旱和半干旱地区的面积约占陆地面积的三分之一,干旱已经成为全球范围内影响植物生长、发育和繁殖等生命活动的主要环境因素。新疆地处我国西北干旱荒漠区,通过对1961—2018年气象观测西北地区年降水量呈现增加的趋势[2],但由于近50年以来增温幅度高达0.34℃·10 a-1,是全球平均增温幅度的近三倍[3],导致蒸发量远大于降水量,对于分布在该地区的植物将面临干旱胁迫的情况更为严峻。因此,从生长、生理及形态结构角度探讨荒漠植物对干旱胁迫的响应、适应机制可以为预测荒漠植物生态系统对全球气候变化的响应提供理论基础,同时也为荒漠植物的保护及更新等工作提供一定的参考[4]。
在面对干旱胁迫时,植物通过改变自身形态结构特征和各个器官的生长速度来抵御逆境[5]。荒漠区恶劣的生态环境及常年的水分亏缺使荒漠植物形成了独特的生存策略[6]。生物量分配是植物面对逆境时维持生长的一种策略[7]。植物的地上部分和地下部分的生长速度、根冠比等形态指标是衡量植物在干旱胁迫下抗旱能力的关键指标[8]。研究表明,荒漠植物会通过减少对地上部分生物量的分配,更多的投入给地下根系,以形成强大的根系系统提高吸收能力[9]。如小叶锦鸡儿(Caragana microphylla)在极端干旱环境下为了更好的利用土壤养分会把大部分生物量分配于地下生物量[10],水分胁迫使得涩荠(Malcolmia africana)和角果藜(Ceratocarpus arenarius)的地下生物量显著增加[11],当土壤含水量逐渐降低时,新疆大叶苜蓿(Medicago sativa.Xinjiang Daye)通过将更多的同化物分配到根系以促进根系生长,进而从土壤中吸取更多的水分提高其抗旱性[12]。
叶片是进行光合作用合成有机物的主要器官,也是暴露在空气中面积最大的器官。叶片解剖結构具有很强的可塑性,使其适应环境的变化[13]。胡杨(Populus euphratica)叶片厚度和栅栏组织厚度随干旱胁迫的加剧表现为不断增加的趋势,狭叶锦鸡儿(Caragana stenophylla Pojark)通过增大表皮细胞壁和角质层厚度,提高叶片储水能力,对于提高水分调节能力具有重要意义[14-15]。
伊犁绢蒿(Seriphidium transiliense)为菊科(Compositae)绢蒿属[Seriphidium(Bess.)Poljak]多年生半灌木植物,是新疆荒漠、荒漠草原主要的建群种和优势种之一[16]。该物种具有明显的抗旱性、抗寒性、耐瘠薄、抗盐碱的特点[17],能涵养水源、防风固沙。本研究以伊犁绢蒿为研究对象,通过盆栽控水法研究持续干旱胁迫下伊犁绢蒿幼苗株高、根系长度、地上、地下部分生物量以及叶片厚度、上下表皮厚度、上下角质层厚度、维管束直径以及栅栏组织的变化规律,探究干旱胁迫对伊犁绢蒿幼苗生长及叶片解剖结构的影响,为伊犁绢蒿在荒漠地区生态环境的保护及和荒漠化草地的恢复等提供理论参考。
1 材料与方法
1.1 试验材料
伊犁绢蒿种子采自新疆昌吉市三工滩的荒漠草地,地理位置43°50′9″ N,87°11′21″ E,海拔994 m。
1.2 试验方法
试验于2022年9月在新疆农业大学人工气候室中开展。培养条件为(24±3)℃下光照培养16 h,(16±3)℃下黑暗培养8 h,相对湿度设定为(30±5)%,光照强度设定为400 μmol·m-2·s-1。将伊犁绢蒿种子播种在装有500 g蛭石的花盆中(上口直径18 cm×下底直径14 cm×高18 cm),每盆播15粒种子,每2 d用蒸馏水补充蒸散的水分,每7 d用1/2浓度的Hoagland营养液浇灌1次,待幼苗长至20 d时,每盆保留生长一致的幼苗5株。
待幼苗长至45 d时进行干旱胁迫。试验材料分成两组:一组土壤相对含水量(Soil relative water content,SRWC)为田间持水量的80%~85%(CK),另一组采用持续干旱胁迫的方法。对同一发育时期的对照组和干旱组每5 d进行一次取样,对照组和干旱组各取5盆,3次重复。每次取样均测定处理组和对照组的株高、根系长度、地上部分和地下部分生物量,同时取叶片(自上往下数第3~4片),用FAA固定液存储,用于测定解剖结构。
1.3 测定指标与方法
生长指标测定:用直尺测量株高和根长,之后将地上植株和地下根系分离,放入75℃烘箱中烘干48 h,称重。每盆有5株幼苗,每次取样5盆,共25株,3次重复。通过地下部分与地上部分干重比值计算根冠比,计算公式为:根冠比=地下部分干重/地上部分干重。
叶片解剖结构:采用石蜡切片法。于Nikon-E100 光学显微镜下观察并拍照。用NISElements D 4.10.00 软件对叶片厚度、上下表皮厚度、栅栏组织、上下角质层厚度、维管束直径等进行测量。每个处理观察5张切片,每张切片观察3个叶片中部视野,3次重复。
1.4 数据分析
用SPSS 19.0 进行数据统计分析,各试验指标用单因素方差分析(ANOVA)进行显著性检验,用Origin 2021 软件作图。
2 结果与分析
2.1 干旱胁迫对伊犁绢蒿幼苗叶片生长的影响
由图1和图2可知,处理组的SRWC随着干旱时间的加长而不断的减少,在干旱处理第40 d时,SRWC仅有6.38%。当SRWC在60%~85%时,叶片生长状态未受到明显影响,当SRWC在35%~55%時,基部叶片开始黄化,叶片下垂,上部叶片颜色由绿色转为灰绿色,而当SRWC小于10%~25%时,基部叶片干枯死亡,中部叶片黄化,顶端叶片生长缓慢,且叶片小型化,当SRWC低于10%时,幼苗顶端叶片呈灰绿色且叶片卷曲下垂,其余叶片发黄干枯。干旱胁迫时间与SRWC的关系见图1。
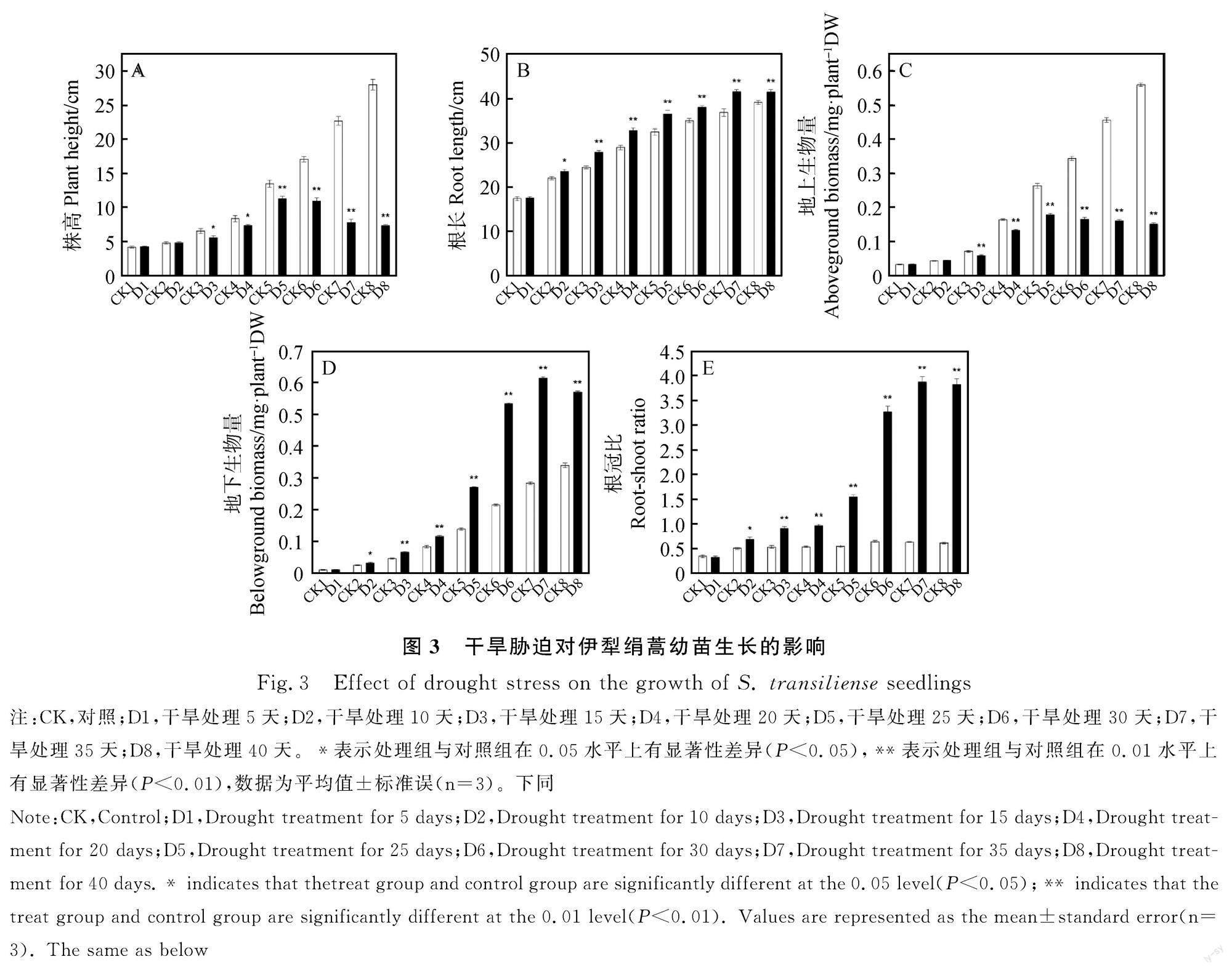
2.2 干旱胁迫对伊犁绢蒿幼苗生长的影响
干旱胁迫对伊犁绢蒿幼苗的生长存在显著影响(图3)。随着干旱胁迫程度的加剧,株高和地上生物量呈先增加后降低的趋势,当SRWC≤53.5%时,处理组与对照组均呈现显著差异,当SRWC为21.94%时,株高和地上生物量都达到最大值,当SRWC为6.38%时,与对照相比分别降低了73.83%和73.05%;随着干旱胁迫程度的加剧,根长、地下生物量和根冠比呈逐渐增加的趋势,当SRWC为≤69.64%时,处理组与对照组均呈显著差异(P<0.05),当SRWC为11.37%时,根长、地下生物量和根冠比达到最大值,当SRWC为6.38%时,与对照相比,根长、地下生物量和根冠分别增加了6.09%,67.92%和527.49%(P<0.01)。
2.3 干旱胁迫对伊犁绢蒿幼苗叶片解剖结构的影响
由图4可知,通过光学显微镜观察,伊犁绢蒿的叶片上下解剖结构相似,其栅栏组织由2~3层柱状细胞组成,无海绵组织,为等面叶叶片类型。上下表皮由一层不规则椭圆状细胞组成。能观察到上下表皮之上覆盖有角质层,上、下表皮处均有气孔分布、无规律。叶片中含有叶脉维管束多束,大小叶脉错综分布呈网状,每一个维管束外侧紧密围绕一层较大的薄壁细胞形成“花环结构”。
2.3.1 干旱胁迫下伊犁绢蒿幼苗叶片厚度和表皮厚度的变化 随着干旱胁迫程度的加剧,叶片厚度呈现先升高后降低的趋势(图5),当SRWC在11.37%~69.64%之间,处理组比对照组呈现显著性差异(P<0.01),当SRWC为21.94%时,叶片厚度达到最大值707.07 μm,比对照组增加了27.27%。随着干旱胁迫程度的加剧,叶片的上下表皮厚度均呈现先增加后降低的趋势,当SRWC在21.94%~53.5%之间,处理组显著高于对照组(P<0.01),上表皮厚度分别比对照组增加了47.71%,68.16%和48.84%,而当SRWC为6.38%时,处理组显著低于对照组,比对照组降低了26.66%;下表皮厚度与上表皮变化基本一致,当SRWC为21.94%~53.5%之间,处理组显著高于对照组,分别比对照组增加了56.23%,61.78%和41.21%,当SRWC为35.08%时,上下表皮厚度均达到最大值。
2.3.2 干旱胁迫下伊犁绢蒿幼苗叶片角质层的变化 随着干旱胁迫程度的加剧,叶片的上、下角质层厚度都呈现逐渐增加的趋势(图6),当SRWC为≤53.5%时,处理组上、下角质层厚度与对照组相比均呈显著性差异,当SRWC为6.38%时,上、下角质层厚度均达到最大值,分别比对照组增加了118.82%和127.39%。
2.3.3 干旱胁迫下伊犁绢蒿栅栏组织及维管束的变化 随着干旱胁迫程度的加剧,伊犁绢蒿的叶肉结构也有所变化(图7)。对照条件下栅栏组织的细胞呈长柱形,且细胞排列整齐、紧密,而处理条件下的栅栏组织厚度呈逐渐降低趋势,当SRWC为≤35.08%时,处理组与对照组出现显著性差异(P<0.01),SRWC为6.38%时,达到最小值,处理组比对照组显著降低了58.86%。
主脉维管束直径随着干旱胁迫程度的加剧呈先增加后减小的趋势,当SRWC≤53.5%时,处理组与对照组出现显著性差异(P<0.05),在SRWC为16.8%时达到最大值,比对照组显著增加了34.32%。
3 讨论
3.1 干旱胁迫对伊犁绢蒿幼苗生长的调控
植物的形态结构能反应植物在应对干旱胁迫下的生长变化,也是作为评价植物抗旱性的重要依据[4]。干旱胁迫下,植物的生长发育会受到抑制,植物在受到胁迫过程中会采取调整资源配置的方式来应对胁迫带来的影响,一些植物会减少地上部分的养分和水分供给,而向地下部分投入更多[18]。烤烟(Nicotiana tabacum L.)的生长发育受干旱胁迫的影响,株高下降[19];饲料油菜(Brassica napus L.)的地上部分干物质积累量、生物量在干旱胁迫的影响下逐渐减小[20];黑麦草(Secale cereale L.)的地上生物量和地下生物量总体呈下降趋势[21]。紫花苜蓿(Medicago sativa L.)在面临干旱胁迫时,株高、地上生物量会降低,而根长和根冠比会增加[22]。本研究中,干旱胁迫对伊犁绢蒿幼苗的生长产生了显著影响,随着干旱胁迫的加剧,幼苗的株高和地上生物量呈先升高后降低的趋势,根长和地下生物量以及根冠比整体呈显著增加趋势,这可能是因为严重胁迫下抑制了地上部分的生长,且根系是植物生存和生长过程中对土壤水分和营养物质吸收的主要器官,幼苗需要通过增加根长和土壤的接触面积吸收更多的水分和养分来应对胁迫。
3.2 干旱胁迫对伊犁绢蒿叶片解剖结构的影响
在干旱胁迫下,叶片作为植物的生长发育中直接与空气接触暴露面积最大,且具有蒸腾作用和光合作用的器官[23],会受到严重的伤害。由于叶片是植物体内储水的主要器官,植物在长期干旱胁迫下,为保证自身的正常生长,减少水分的流失,植物叶片的外观形态和微观结构会发生相应程度的改变[24],这些结构的改变在一定程度上可以反映植物的生理适应性,常用来作为植物是否具有抗旱性的重要依据。
不同程度的干旱胁迫下叶片结构的响应有所不同。菊科蒿属植物会通过增加叶片厚度、角质层厚度来适应环境[25]。研究表明,较厚的叶片和表皮細胞能够减少植物体内的水分散失,具有一定的贮水能力,可以增强抗旱性[26]。针茅叶片在干旱胁迫下叶片总厚度和角质层厚度显著增加[27]。本研究中,幼苗叶片厚度和上下表皮厚度随着干旱胁迫的加剧都呈现先增加后减少的趋势,究其原因可能是幼苗通过表皮细胞的不断增厚来储存水分,但随着胁迫程度的加剧,影响了植物体内的水分代谢,叶片失水严重进而导致叶片细胞的水分减少,使叶片细胞的生长和分裂受到抑制,也可能是幼苗应对干旱胁迫的适应性变化,这与吴丽君等[28]研究结果一致。
角质层是植物叶片的主要保护组织,干旱胁迫使叶片表皮的气孔关闭,植物水分主要通过表皮角质层蒸散,发达的角质层能防止植物体内水分过份蒸腾[29]。水分胁迫的增加导致叶片和角质层厚度的显著增加。这与Kofidis等[30]的研究一致,即在干燥条件下生长的植物通常比在潮湿条件下生长的植物具有更厚的叶片和角质层。文冠果(Xanthoceras sorbifolia Bunge)在长期适应干旱中具有较厚的角质层和细胞壁[31];不同生境酸枣(Ziziphus jujuba var. spinosas)随干旱程度加剧,叶片厚度和叶片角质层厚度增加[32]。本研究中,随着干旱胁迫的加剧,上下表皮角质层厚度显著增加,说明伊犁绢蒿叶片通过增加角质层厚度,降低蒸腾,从而增强抗旱性。
叶肉是位于叶片上下表皮之间最发达的组织,不同植物在应对干旱时会采取不同策略,演化成不同的叶肉类型[33]。本研究中,伊犁绢蒿的叶肉为全栅型,栅栏组织发达,能很大程度的减少暴露在空气中的面积,保存叶片水分。旱生植物高度发达的栅栏组织可以防止强烈光照对叶肉细胞的灼伤,有效的利用衍射光进行光合作用[34]。干旱胁迫下,油茶(Camellia oleifera)组织结构发生改变,栅栏组织厚度呈减小趋势,从而减少叶片水分流失增强了抗旱性[35]。本研究中伊犁绢蒿幼苗上、下表皮均有排列紧密的栅栏组织,在干旱胁迫的影响下,叶片栅栏组织的厚度显著降低,细胞逐渐变小,细胞间隙变小,排列逐渐紧密,有利于对光能的利用。
发达的维管系统可提高植物水分和养分的运输效率,减缓干旱对叶片造成的萎蔫损伤[36]。于海燕等[37]研究结果表明,在干旱胁迫下,文冠果的叶脉维管系统发达。本研究中,伊犁绢蒿幼苗主脉维管束直径呈先增加后减小的趋势,在重度胁迫下减小,究其原因可能是幼苗通过维管束直径高效运输水分,而重度胁迫下抑制了植物的维管系统发育。
4 结论
综上所述,随着干旱胁迫程度的加剧,伊犁绢蒿幼苗的株高和地上生物量先增加后降低,而根长、地下生物量和根冠比增加;叶片厚度、上下表皮厚度和维管束直径先增加后降低,栅栏组织逐渐降低,叶片角质层厚度增加。表明培养45 d的伊犁绢蒿幼苗在面对干旱胁迫时能及时调整分配策略,提高其对干旱胁迫的适应能力。
参考文献
[1]尹晗,李耀辉.我国西南干旱研究最新进展综述[J].干旱气象,2013,31(1):182-193
[2]王澄海,张晟宁,李课臣,等.1961~2018年西北地区降水的变化特征[J].大气科学,2021,45(4):713-724
[3]李明,孙洪泉,苏志诚.中国西北气候干湿变化研究进展[J].地理研究,2021,40(4):1180-1194
[4]陈爱萍,隋晓青,王玉祥,等.干旱胁迫及复水对伊犁绢蒿幼苗生长及生理特性的影响[J].草地学报,2020,28(5):1216-1225
[5]ANJUM S A,ASHRAF U,ZOHAIB A,et al. Growth and developmental responses of crop plants under drought stress:A review[J].Zemdirbyste Agriculture,2017,104(3):267-276
[6]王方琳,柴成武,赵鹏,等.3种荒漠植物光合及叶绿素荧光对干旱胁迫的响应及抗旱性评价[J].西北植物学报,2021,41(10):1755-1765
[7]HOOVER,DAVID L,KORIAKIN,et al. Comparing water-related plant functional traits among dominant grasses of the Colorado Plateau:Implications for drought resistance[J].Plant and Soil,2019,441(1-2):207-218
[8]陈静,宫文龙,张尚雄,等.紫花苜蓿与蓝花苜蓿抗旱性比较研究[J].中国草地学报,2020,42(2):32-40
[9]席璐璐,缑倩倩,王国华,等.荒漠绿洲过渡带一年生草本植物对干旱胁迫的响应[J].生态学,2021,41(13):5425-5434
[10]牛存洋,阿拉木萨,宗芹,等.科尔沁沙地小叶锦鸡儿地上-地下生物量分配格局[J].生态学杂志,2013,32(8):1980-1986
[11]程军回,张元明.水分胁迫下荒漠地区2种草本植物生物量分配策略[J].干旱区研究,2012,29(3):432-439
[12]罗永忠,李广.土壤水分胁迫对新疆大叶苜蓿的生长及生物量的影响[J].草业学报,2014,23(4):213-219
[13]吴建慧,郭瑶,赵倩竹,等.干旱胁迫对绢毛委陵菜叶片解剖结构和生理指标的影响[J].草业科学,2012,29(8):1229-1234
[14]李端,司建华,张小由,等.胡杨对干旱胁迫的生态适应[J].中国沙漠,2020,40(2):17-23
[15]王海超,王春光,贺晓,等.狭叶锦鸡儿叶片解剖结构对干旱胁迫下荒漠草原退化梯度的响应[J].生态环境学报,2016,25(5):744-751
[16]李莉,董志国,贾纳提,等.干旱胁迫对伊犁绢蒿生理生化指标的影响[J].草原与草坪,2014,34(6):82-85
[17]张晶,崔国盈,张勇娟,等.伊犁绢蒿种质资源保护与利用[J].草食家畜,2016(1):51-56
[18]谢志玉,张文辉.干旱和复水对文冠果生长及生理生态特性的影响[J].应用生态学报,2018,29(6):1759-1767
[19]庄晔,葛嘉雪,汪孝国,等.干旱胁迫后复水对烤烟生长及其生理特性的影响[J].中国烟草学报,2022,28(4):48-58
[20]王悦,高山,程凯乐,等.干旱胁迫对不同品种饲料油菜生长发育、干物质积累及产量的影响[J].江蘇农业科学,2023,51(10):155-163
[21]杨丹,张舒芸,田新会,等.不同黑麦品种(品系)对持续干旱胁迫的形态适应性研究[J].草地学报,2020,28(1):122-129
[22]ZHANG C M,SHI S L,WANG B W,et al. Physiological and biochemical changes in different drought-tolerant alfalfa (Medicago sativa L.) varieties under PEG-induced drought stress[J].Acta Physiologiae Plantarum,2018,40(2):25
[23]赵雪,张秀珍,牟洪香,等.文冠果幼苗叶片解剖结构和光合作用对干旱胁迫的响应[J].北方园艺,2017(13):38-44
[24]宋鹏,丁彦芬,朱贵珍,等.6种卫矛属植物叶片解剖结构与抗旱性评价[J].河南农业大学学报,2019,53(4):574-580
[25]孙萌,尚忠海,沈植国,等.植物对干旱胁迫响应的研究进展[J].河南林业科技,2019,39(4):1-3,44
[26]宋捷,田青.4种园林植物叶片的解剖结构及抗旱性[J].兰州大学学报(自然科学版),2022,58(2):262-269
[27]BOUGHALLEB,FAYCAL,NEFFATI,et al. Anatomical adaptations of the desert species Stipa lagascae against drought stress[J].Biologia:Casopis Slovenskej Akademie Vied,2015,70(8):1042-1052
[28]吴丽君,李志辉,杨模华,等.赤皮青冈幼苗叶片解剖结构对干早胁迫的响应[J].应用生态学报,2015,26(12):3619-3626
[29]岑湘涛,沈伟,牛俊乐,等.基于植物叶片解剖结构的抗逆性评价研究进展[J].北方园艺,2021(18):140-147
[30]KOFIDIS G,BOSABALIDIS A M,CHARTZOULAKIS K. Leaf anatomical alterations induced by drought stress in two avocado cultivars[J].Journal of Biological Research,2004,(1):115-120
[31]马小芬,王兴芳,李强,等.不同种源地文冠果叶片解剖结构比较及抗旱性分析[J].干旱区资源与环境,2013,27(6):92-96
[32]朱广龙,魏学智.酸枣叶片结构可塑性对自然梯度干旱生境的适应特征[J].生态学报,2016,36(19):6178-6187
[33]马红英,吕小旭,计雅男,等.17种锦鸡儿属植物叶片解剖结构及抗旱性分析[J].水土保持研究,2020,27(1):8
[34]崔秀萍,刘果厚,张瑞麟.浑善达克沙地不同生境下黄柳叶片解剖结构的比较[J].生态学报,2006(6):1842-1847
[35]何小三,王玉娟,徐林初,等.干旱胁迫对不同油茶品种叶片解剖结构的影响[J].中南林业科技大学学报,2020,40(10):1-17
[36]崔晓晗,叶丽红,贾文龙,等.不同土壤相对含水量下沙棘叶片解剖结构研究[J].北方农业学报,2023,51(2):74-82
[37]于海燕,胡潇予,何春霞,等.文冠果不同种源叶片结构对水分胁迫的差异性响应[J].北京林业大学学报,2019,41(1):57-63
(责任编辑 刘婷婷)
猜你喜欢
小读者(2021年2期)2021-03-29
疯狂英语·新悦读(2019年11期)2019-12-18
华人时刊(2019年13期)2019-11-17
NBA特刊(2018年21期)2018-11-24
文苑(2018年22期)2018-11-19
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
山东农业科学(2016年11期)2016-12-17
