
补播苜蓿对退化草地植物群落结构和生产力的影响途径
2024-02-02 06:06郭美琪郭童天徐民乐张英俊
草地学报 2024年1期
郭美琪 郭童天 徐民乐 张英俊

doi:10.11733/j.issn.1007-0435.2024.01.005
引用格式:
郭美琪, 郭童天, 徐民乐,等.补播苜蓿对退化草地植物群落结构和生产力的影响途径[J].草地学报,2024,32(1):46-53
GUO Mei-qi, GUO Tong-tian, XU Min-le,et al.Mechanism of Reseeding Alfalfa Improving Productivity and Plant Community Structure of Degraded Grassland[J].Acta Agrestia Sinica,2024,32(1):46-53
收稿日期:2023-07-17;修回日期:2023-08-31
基金项目:中国科学院战略性先导科技专项(XDA26000000)子课题:呼伦贝尔家庭牧场草畜平衡技术体系研究(XDA26010302);国家自然科学基金重大项目(32192462)和国家牧草产业技术体系项目(CARS-34)资助
作者简介:
郭美琪(1997-),女,满族,辽宁丹东人,博士研究生,主要从事草地生态与管理方面研究,E-mail:B20213241026@cau.edu.cn;*通信作者Author for correspondence,E-mail:zhangyj@cau.edu.cn
摘要:近年来,紫花苜蓿(Medicago sativa L.)和黄花苜蓿(Medicago falcata L.)被广泛选作补播材料,用于修复退化草地。尽管研究表明二者的植物学特征存在差异,但鲜有人关注它们是否会通过不同的方式影响退化草地的生产力。为了探究二者对退化草地的修复效果是否存在差异,并确定影响补播地生产力的限制因素,本研究选取黄花苜蓿和紫花苜蓿作为补播种对退化草甸草原进行了补播,并在隔年对草地植被、土壤理化性质和微生物量进行监测。结果表明,两种苜蓿都可以显著增加退化草地生产力并改善土壤健康状况。然而,紫花苜蓿通过增加土壤铵态氮改变植被群落结构,其补播样地生产力也由土壤速效养分影响;黄花苜蓿通过增加土壤速效磷含量改变植被群落结构,其补播样地生产力由植被群落结构和土壤微生物量共同影响。初步结果表明,黄花苜蓿更适宜在养分贫瘠的退化草地进行补播。此外在选择紫花苜蓿补播时,可以适当施肥以提高补播成功率。
关键词:补播;黄花苜蓿;紫花苜蓿;植物群落结构;生产力
中图分类号:S963.22+3.3 文献标识码:A 文章編号:1007-0435(2024)01-0046-08
Mechanism of Reseeding Alfalfa Improving Productivity and Plant
Community Structure of Degraded Grassland
GUO Mei-qi1, GUO Tong-tian1, XU Min-le1, ZHANG Ying-jun1,2*
(1.College of Grassland Science and Technology,China Agricultural University,Beijing 100193,China;
2.Grassland Management and Utilization Laboratory,Ministry of Agriculture and Rural Affair,Beijing 100193,China)
Abstract:In recent years,Medicago sativa L. and Medicago falcata L. have been widely chosen as reseeding materials for the restoration of degraded grasslands. Although studies have shown differences in their botanical characteristics,little attention has been given to whether they would have differential effects on the restoration of degraded grasslands. In order to investigate whether there are differences in the productivity of M. sativa L. and M. falcata L. on degraded grasslands,and to determine the limiting factors for successful reseeding,this study selected M. sativa L. and M. falcata L. as reseeding species and conducted a monitoring of vegetation,soil physicochemical properties,and microbial biomass in the following year. The results showed that both alfalfa species significantly increased productivity and improved soil health of degraded grasslands. However,M. sativa L. altered the vegetation community structure by increasing soil ammonium nitrogen,and the productivity of the reseeding plots was influenced by soil available nutrients. M. falcata L. altered the vegetation community structure by increasing soil available phosphorus,and the productivity of the reseeding plots was influenced by both vegetation community structure and soil microbial biomass. Preliminary results suggested that M. falcata L. was more suitable for reseeding in nutrient-poor degraded grasslands. Additionally,when selecting M. falcata L. for reseeding,appropriate fertilization should be applied to improve the success rate of reseeding.
Key words:Reseeding; Medicago falcata L.; Medicago sativa L.;Plant community structure;Productivity
豆科植物作為天然草地的重要组分之一,可以通过共生固氮和氮转移减少草地对无机氮肥的依赖[1-2]。因此豆科牧草成为近年来退化植被补播修复的首选材料之一[3-6]。其中,苜蓿属植物具有产量高且蛋白丰富的特点,被广泛选择作为补播种[7-9]。如Zhou等[9]的研究发现补播黄花苜蓿(Medicago falcata L.)和紫花苜蓿(Medicago sativa L.)都能显著提高草地生产力,但黄花苜蓿补播草地拥有更好的持久性。这表明即使是同属豆科植物,其对退化草地的修复效果也存在差异。但造成这种差异的原因,很少有人关注。
根系是植物与土壤环境进行物质能量交换的主要桥梁,其形态与分布反映了植物对土壤的利用状况,直接影响植株的生长状况[10]。已有研究表明,虽然同为苜蓿属牧草,但黄花苜蓿和紫花苜蓿的根系形态和分布存在显著差异。紫花苜蓿多为轴根型牧草,而黄花苜蓿根系较为多样化,主要为侧根型,轴根型和根蘖型[11],这种独特的根系构型,导致黄花苜蓿更侧重侧根的生长[12-13]。还有研究表明,相较于紫花苜蓿,黄花苜蓿拥有更多的主茎节数,因此黄花苜蓿拥有更强的抗旱性和地方适应性[14]。但紫花苜蓿主根直径和入土深度较大,有利于大范围吸收土壤养分和水分,同时紫花苜蓿还拥有更高的分蘖数,从而使其拥有更高的产量[14-15]。因此补播黄花苜蓿和紫花苜蓿对退化草地的修复效果不同可能是因为植物学特征的差异导致的,尤其是根系差异。
根据前人研究,植物根系的差异会导致不同的土壤氮磷含量及形态[16],同时还可以通过根系分泌物和根凋落物质量影响根际微生物[17]。因此,紫花苜蓿和黄花苜蓿可能因为根系的差异导致其在在补播定植过程中对退化草地的土壤理化性质和微生物产生不同的影响。如Li等[18]发现,紫花苜蓿和羊草(Leymus chinensis)混播草地相较于黄花苜蓿和羊草混播草地拥有更高的土壤总氮和速效氮含量以及更高的生产力,但其他土壤参数是否存在差异作者并未关注。因此,有必要研究比较紫花苜蓿和黄花苜蓿对退化草地的土壤参数的影响是否存在差异,进而导致植被群落结构发生不同的变化,从而通过不同的途径影响退化草地生态系统功能。这项研究将帮助我们确定黄花苜蓿和紫花苜蓿补播成功的关键限制因素,提高补播成功率,为充实与修复退化植被相关的理论基础与实践提供依据。
1 材料与方法
1.1 研究区域概况
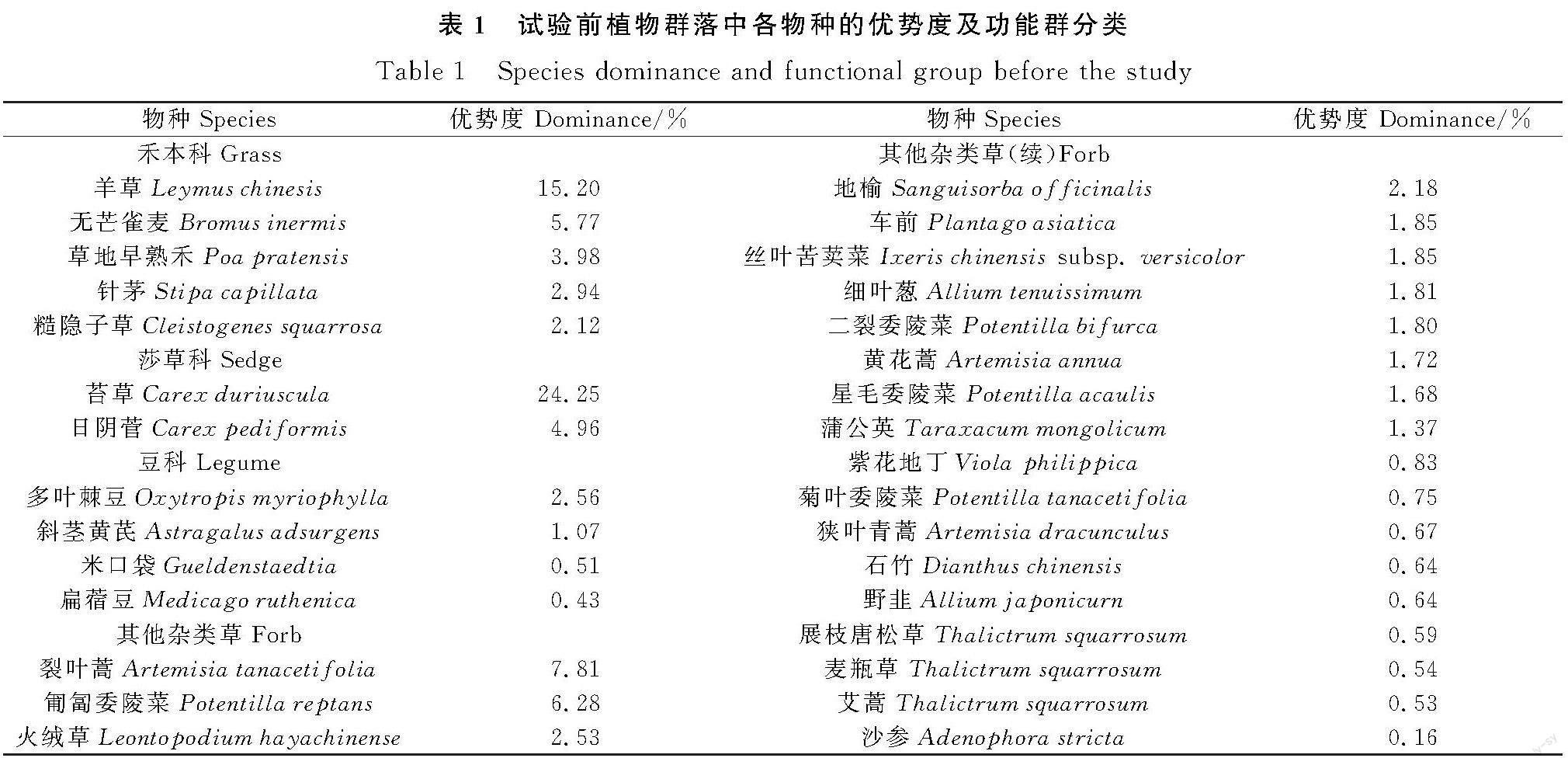
本试验地点位于内蒙古自治区呼伦贝尔市陈巴尔虎旗特尼河农牧场第九生产队(49°20′~ 49°26′N,119°55′~ 120°9′E,海拔628 ~ 649 m)。年均温-3℃~0℃,无霜期90~115 d,年平均降水量350~410 mm。试验样地为中轻度退化草甸草原,试验前作为打草场使用(每年8月打草),以羊草、无芒雀麦(Bromus inermis)为优势种,主要伴生植物有二裂委陵菜(Potentilla bifurca)、日阴菅(Carex pediformis)等,具体各物种优势度及功能群分类见表1。试验前样地土壤理化性质如下:pH值5.75,全碳含量为45.72 mg·g-1,速效氮含量77.77 mg·kg-1,速效磷6.79 mg·kg-1,速效钾296.06 mg·kg-1。
1.2 试验设计
试验于2020年6月开展,采用配置倒T型开沟器的免耕补播机进行播种,条播行距为20 cm,开沟深度为5 cm,补播后当年休牧。试验采用单因素随机区组设计,以黄花苜蓿(Mf)和紫花苜蓿(Ms)作为补播种,同时以未补播(Lc)的同期围封样地作为对照,每处理设置 4个重复,共12个小区,小区间隔为3 m,小区面积为100 m2(10 m×10 m)。其中种子播量 15 kg·hm-2,补播处理采用配置倒T型开沟器的免耕补播机进行播种,开沟深度为5 cm,行距设置为15 cm,播种同时施肥以促进苜蓿定植,其中磷酸二铵(P2O5≥46%)施用量为135 kg·hm-2,硫酸钾的施用量为30 kg·hm-2。未补播处理进行划破草皮同时予以施肥处理,以确保补播物种为唯一控制变量。
供试验使用的草种:紫花苜蓿品种名称为‘龙牧801,来源于黑龙江畜牧研究所,其特点为抗寒耐碱性较强,发芽率为90%;黄花苜蓿为呼伦贝尔当地采集草种,种子发芽率为92%。
1.3 植被监测与取样
本试验于2021年8月下旬生物量现存量最大时进行植被地上调查和土壤取样,在处理小区内随机放置3个0.5 m×0.5 m的样方进行植被调查,将样方内植物分种齐地刈割,带回实验室65℃烘至恒重,利用最大现存量估计地上净初级生产力(ANPP)。
苜蓿根瘤的采集与植被调查同期进行。于黄花苜蓿补播小区和紫花苜蓿小区植被调查样方附近随机选择3株苜蓿进行采样,每个处理共采集36株苜蓿,然后测定其有效根瘤数量。采根瘤样品时,使用铁锹垂直于地面向地下挖掘,连土撬起(0~20 cm),尽量将根系完整挖出,用蒸馏水小心清洗,对新鲜有效根瘤进行计数。
1.4 土壤样品采集及测定
在植被调查的样方中用土钻法取两钻0~20 cm表层土壤样品,混合均匀后带回实验室,过2 mm筛分离出植物根系。将筛下的土壤分成两份,一份室内遮阴风干保存,一份保存在-20℃的冰箱里,用于后续的土壤理化性质和微生物指标的测定。测定的土壤理化性质包括土壤含水量(Soil water content,SWC)、pH值、速效氮(Soil available nitrogen,AN)、速效磷(Soil available phosphorus,AP)、有机碳(Soil organic carbon,SOC)、总碳(Soil total carbon,STC)、全氮(Soil total nitrogen,STN),微生物指标包括微生物量碳(Microbial biomass carbon,MBC)和微生物量氮(Microbial biomass nitrogen,MBN)。
其中采用铝盒烘干法测定土壤含水量;采用Sartorius普及型pH计PB-10(德国)测定土壤pH值;采用元素分析仪(Vario Macro,elementar,德国)测定土壤全氮以及有机碳含量;采用流动分析仪(AutoAnalyser 3,Seal Analytical,Norderstedt,德国)测定土壤铵态氮、硝态氮含量;采用Olsen的方法测定土壤速效磷含量[20]。MBC和MBN采用氯仿熏蒸浸提法,浸提后使用TOC-VCPH有机碳分析仪(Vario TOC,德国)测定[21]。
1.5 统计分析
统计分析与图表绘制均使用R 4.0.3完成。植被物种多样性指标通过vegan包进行计算,包括物种丰富度和香农多样性(Shannon Wiener)指数。数据分析前使用shapiro.test函数(stats)进行正态性检验。多样性指数计算公式如下:
Shannon Wiener指数:H=-∑Si=1PilnPi
式中:Pi是物种i的相对生物量,S为植物群落的物种丰富度,即各水平下的物种数总和。
植被群落组成使用ape包采用主坐标分析量化[19]。所有指标使用aov函数运行单因素方差分析,对处理效应進行分析,事后多重比较采用Duncan方法(duncan.test函数,agricolae),使用ggplot2和ggsci等程序包进行绘图。
2 结果与分析
2.1 补播苜蓿对草地生产力、群落组成以及根系结瘤数的影响
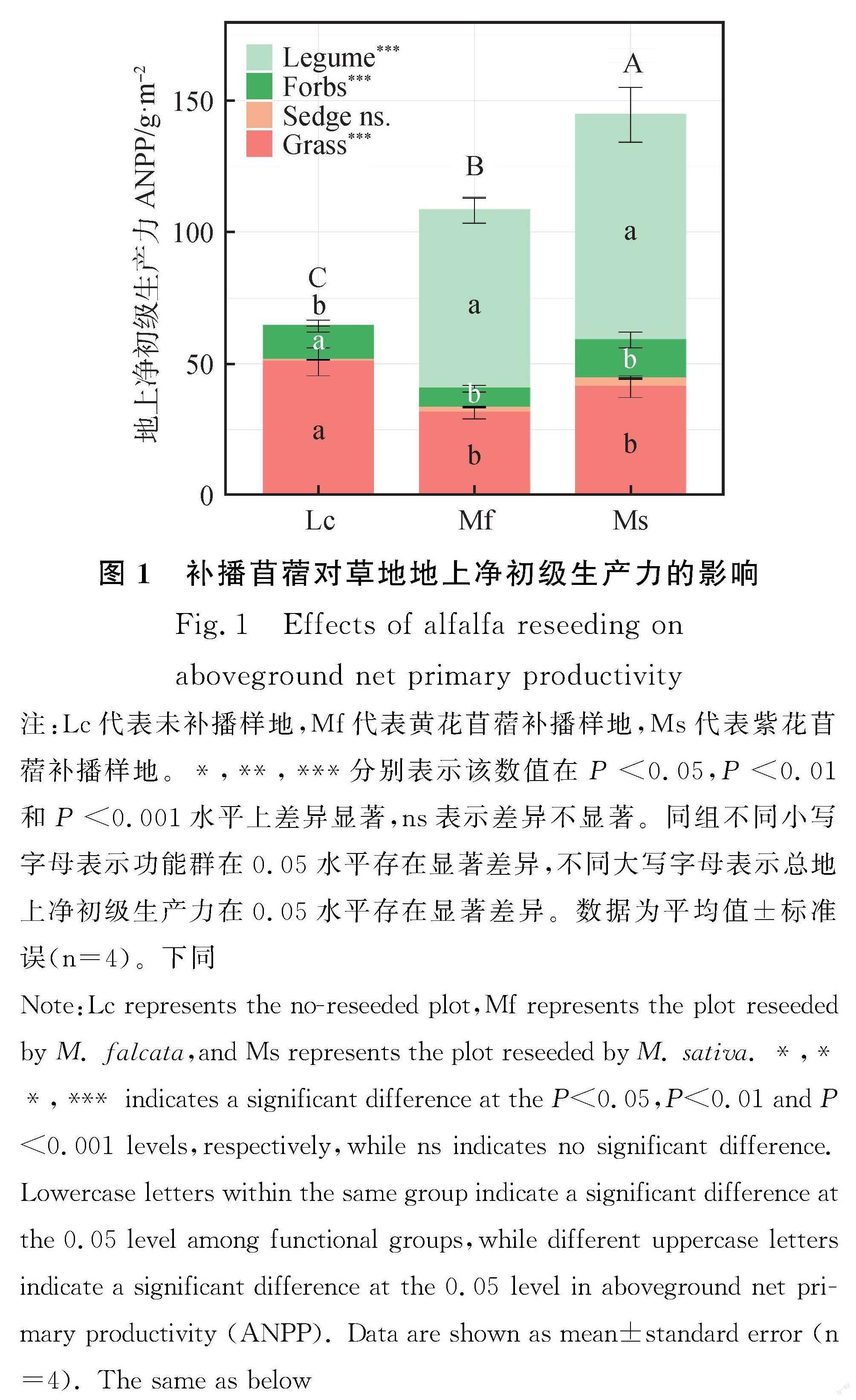
与未补播的同期围封样地相比,补播紫花苜蓿和黄花苜蓿均显著增加草地的ANPP(图1,P<0.001),黄花苜蓿补播草地的ANPP提高68.3%,紫花苜蓿补播草地的ANPP提高了124.7%。补播草地生产力提升主要通过增加豆科牧草生产力来实现(P<0.001)。
相较于未补播处理,两种补播处理均显著增加了植物物种丰富度(图2a,P < 0.001)和香农多样性指数(图2b,P < 0.001)。补播黄花苜蓿和紫花苜蓿均改变了植被群落组成,增加了豆科功能群的优势度的同时降低了禾草和杂类草的优势度,但莎草的优势度并未发生显著变化(图3a)。基于Bray-Curtis距离的主坐标分析(PCoA)结果显示,补播黄花苜蓿、紫花苜蓿和未补播样地的植被群落结构发生显著分离,表明补播苜蓿显著改变了退化草地的植被群落结构(图3b,P = 0.001)。
除此之外,我们还对黄花苜蓿补播草地和紫花苜蓿补播草地苜蓿根系有效结瘤数进行了测定,结果表明紫花苜蓿根瘤数显著高于黄花苜蓿(图4,P<0.001)。
2.2 补播苜蓿对草地土壤理化性质和微生物量碳氮的影响
补播苜蓿显著增加了土壤pH值,也显著改变了土壤速效养分含量:土壤硝态氮(NO-3-N)和AN含量在两种补播处理下均显著增加,铵态氮(NH+4-N)含量只在补播紫花苜蓿处理下显著增加;AP含量只在补播黄花苜蓿后显著增加。但补播苜蓿一年后土壤水分含量,总氮,总碳,有机碳含量并未发生显著变化(表1)。此外,对于土壤微生物而言,我们发现补播苜蓿均显著增加了土壤MBN和MBC的积累(图5)。
2.3 植物群落结构与土壤环境因子的关系
土壤环境因子与植被群落组成之间的典型分析(CCA)结果显示,植物群落的变化主要由土壤pH值,NO-3-N,NH+4-N,AP以及MBC和MBN几个土壤环境因子解释(图6,表1)。CCA1轴和CCA2轴中的环境因子组合对植物群落变化的解释率为86.16%,补播紫花苜蓿通过增加土壤NH+4-N含量从而导致植被群落结构发生变化,补播黄花苜蓿则主要通过增加土壤AP含量改变植被群落结构。
2.4 植物地上净初级生产力的影响因子
通过多元回归分析探究植被组成(PC2)、土壤生物因子及非生物因子对草地ANPP的相对贡献,结果表明:紫花苜蓿补播草地的ANPP主要受土壤速效养分含量影响(图7a),黄花苜蓿补播草地的ANPP主要由植被群落结构和土壤生物因子共同影响(图7b)。
3 讨论
本研究结果发现,补播黄花苜蓿和紫花苜蓿都能显著提高退化草地的生产性能,这与前人研究结果一致,补播苜蓿不仅可以通过空间或时间生态位互补来增强土壤资源的开发利用[20-21],而且还通过根分泌物、菌根网络等途径将氮素转移到邻近植物,从而加强物种间的促进作用[22],促进了生产力的积累的同时,还增加了植物α多样性。
本研究中,无论补播黄花苜蓿还是紫花苜蓿,土壤速效养分和土壤微生物量都得到显著改善,这与前人研究结果一致,豆科牧草加拥有较高质量的凋落物和根系分泌物,不仅可以增草地生态系统氮输入,提升土壤养分的矿化效率和养分有效性[22-23],还可以为微生物提供易降解的碳源,增加群落中微生物生物量和酶活性[24],从而改善生态系统的土壤健康状况。但值得注意的是,与黄花苜蓿补播草地相比,紫花苜蓿补播草地拥有更高的速效氮含量和微生物量,以及更低的速效磷含量。这可能是因为紫花苜蓿拥有更高的根系生物量和主根直径,可以更好的利用土壤养分和水分来促进植物地上部的生长[25-26],增加凋落物的质量与数量,从而促进土壤氮循环。与此同时,我们的研究结果显示,在补播次年,紫花苜蓿固氮能力显著高于同期补播的黄花苜蓿,这可能也是导致紫花苜蓿补播草地可利用氮养分含量更高的原因。此外,生物固氮过程往往会造成土壤磷的高消耗[9,27],这可能就是紫花苜蓿同时拥有更低土壤氮含量的原因。因此,在补播苜蓿时需要伴随磷肥添加来补偿土壤磷的消耗,尤其是紫花苜蓿。
紫花苜蓿补播样地群落结构改变的原因主要是显著增加的土壤NH+4-N含量,而黄花苜蓿补播样地群落结构的改变主要由增加的土壤AP含量影响。这表明即便是同属豆科植物,在补播修复后也会导致群部落结构朝不同方向演替。此外,与我们的假设一致,补播草地和紫花苜蓿补播草地生产力的主要影响因子也不同,紫花苜蓿补播样地生产力主要受土壤速效养分影响,而黄花苜蓿补播样地生产力主要受植物群落结构和土壤微生物量影响。根据前人研究,根系生物量的积累量决定了植物的生长能力[25],紫花苜蓿拥有更高的根系生物量,表明其利用土壤养分和水分的能力更强[26];同时,紫花苜蓿拥有更粗的主根,有利于其大范围吸收土壤养分和水分[15]。因此对于紫花苜蓿补播地而言,提供充足的养分和水分有利于紫花苜蓿的生长,是提高草地生产力的关键。黄花苜蓿根系长且纤细,侧根多,有利于其吸收土壤深层的水分的同时,能充分的利用天然降水[28]。因此,相较于紫花苜蓿,黄花苜蓿拥有更强的抗逆性,养分对其限制较小,更适宜在土壤养分贫瘠的退化草地进行补播。我们的研究结果显示黄花苜蓿补播样地拥有更高的速效磷含量。根据前人研究结果,丛枝菌根真菌(AMF)和植物根系可通过协作促进中等易分解态有机磷和原生矿物态磷向易分解态磷转换,增加土壤可利用磷含量[29-30],所以黄花苜蓿可能更依赖于与AMF的共生。此外植被群落结构也是黄花苜蓿样地的草地生产力的重要影响因素,因此黄花苜蓿补播样地生产力的提升可能主要通过改善土壤微生物,促进植被群落与微生物的共生关系来实现。但如果想得到进一步的验证,还需要对黄花苜蓿和紫花苜蓿的根际微生物进行具体的测定。
4 结论
补播紫花苜蓿和黄花苜蓿都能提高草地生产力并改善土壤健康状况,达到修复退化草地的目的。但补播紫花苜蓿和黄花苜蓿分别通过增加土壤铵态氮含量和速效磷含量导致植被群落结构发生不同的变化,从而导致生产力的影响途径不同:紫花苜蓿补播样地生产力主要由土壤速效养分影响,黄花苜蓿补播样地生产力主要由植物群落结构和微生物量影响。
参考文献
[1]RASMUSSEN J,GYLFADOTTIR T,LOGES R,et al. Spatial and temporal variation in N transfer in grass-white clover mixtures at three Northern European field sites [J]. Soil Biology & Biochemistry,2013(57):654-662
[2]ABALOS D,DE DEYN G B,PHILIPPOT L,et al. Manipulating plant community composition to steer efficient N-cycling in intensively managed grasslands [J]. Journal of Applied Ecology,2021,58(1):167-180
[3]ZHOU J Q,ZHANG F G,HUO Y Q,et al. Following legume establishment,microbial and chemical associations facilitate improved productivity in degraded grasslands [J]. Plant and Soil,2019,443(1-2):273-292
[4]岳麗楠,师尚礼,祁娟,等. 免耕补播对北方退化草地生产力及营养品质的影响[J]. 草地学报,2021,29(11):2583-2590
[5]孙伟,刘玉玲,王德平,等. 补播羊草和黄花苜蓿对退化草甸植物群落特征的影响[J]. 草地学报,2021,29(8):1809-1817
[6]ZHANG Y,ZHOU J,YANG G,et al. Theory and application of no-tillage reseeding in degraded grasslands [J]. Chinese Science Bulletin,2020,65(16):154615-154655
[7]李强,丛山,赵成振,等. 播期和播前刈割对羊草草甸中紫花苜蓿建植的影响[J]. 草业学报,2022,31(11):94-104
[8]阎子盟,陈倩,张玉娟,等. 河北坝上天然割草地补播豆科牧草早期建植的研究[J]. 草地学报,2016,24(1):178-183
[9]ZHOU J,WILSON G W T,COBB A B,et al. Phosphorus and mowing improve native alfalfa establishment,facilitating restoration of grassland productivity and diversity [J]. Land Degradation & Development,2019,30(6):647-657
[10]李鹏,李占斌,澹台湛. 黄土高原退耕草地植被根系动态分布特征[J]. 应用生态学报,2005 (5):849-853
[11]王红梅. 不同地理来源野生黄花苜蓿形态学、解剖学比较研究[D]. 呼和浩特:内蒙古农业大学,2009:16
[12]徐大伟,徐丽君,辛晓平,等. 呼伦贝尔地区不同多年生牧草根系形态性状及分布研究[J]. 草地学报,2017,25(1):55-60
[13]南丽丽,师尚礼,张建华. 不同根型苜蓿根系发育能力研究[J]. 草业学报,2014,23(2):117-124
[14]白晨晨,孙赛赛,王双双,等. 安徽野生黄花苜蓿与紫花苜蓿产量性状、核型及遗传纯合度比较分析[J]. 草原与草坪,2020,40(3):42-46,56
[15]王富贵,于林清,田自华,等. 18个苜蓿品种根系特征及与地上生物量关系的研究[J]. 中国草地学报,2011,33(4):51-57
[16]CREME A,RUMPEL C,GASTAL F,et al. Effects of grasses and a legume grown in monoculture or mixture on soil organic matter and phosphorus forms [J]. Plant and Soil,2016,402(1-2):117-128
[17]SPITZER C M,LINDAHL B,WARDLE D A,et al. Root trait-microbial relationships across tundra plant species [J]. New Phytol,2021,229(3):1508-1520
[18]LI Q,SONG Y T,LI G D,et al. Grass-legume mixtures impact soil N,species recruitment,and productivity in temperate steppe grassland [J]. Plant and Soil,2015,394(1-2):271-285
[19]PARADIS E,CLAUDE J,STRIMMER K J B. APE:analyses of phylogenetics and evolution in R language [J].National Library of Medicine 2004,20(2):289-290
[20]MUELLER K E,TILMAN D,FORNARA D A,et al. Root depth distribution and the diversity-productivity relationship in a long-term grassland experiment [J]. Ecology,2013,94(4):787-793
[21]HANSEN W,WOLLNY J,OTTE A,et al. Invasive legume affects species and functional composition of mountain meadow plant communities [J]. Biological Invasions,2021,23(1):281-296
[22]REAY M K,PEARS K A,KUHL A,et al. Mechanisms of nitrogen transfer in a model clover-ryegrass pasture:a 15 N-tracer approach [J]. Plant and Soil,2022,480(1-2):369-389
[23]WEI X,REICH P B,HOBBIE S E. Legumes regulate grassland soil N cycling and its response to variation in species diversity and N supply but not CO2 [J]. Glob Chang Biol,2019,25(7):2396-2409
[24]SEKARAN U,LOYA J R,ABAGANDURA G O,et al. Intercropping of kura clover (Trifolium ambiguum M. Bieb) with prairie cordgrass (Spartina pectinata link.) enhanced soil biochemical activities and microbial community structure [J]. Applied Soil Ecology,2020(147):10
[25]吴新卫,韩清芳,贾志宽. 不同苜蓿品种根颈和根系形态学特性比较及根系发育能力[J]. 西北农业学报,2007(2):80-86
[26]白文明,左强,黄元仿,等. 乌兰布和沙区紫花苜蓿根系生长及吸水规律的研究[J]. 植物生态学报,2001(1):35-41
[27]ROSCHER C,THEIN S,WEIGELT A,et al. N2 fixation and performance of 12 legume species in a 6-year grassland biodiversity experiment [J]. Plant and Soil,2010,341(1-2):333-248
[28]王月勝. 不同苜蓿品种根系特征及其抗寒性关系的研究[D]. 兰州:甘肃农业大学,2008:20-30
[29]谢欢,张秋芳,陈廷廷,等. 氮添加促进丛枝菌根真菌和根系协作维持土壤磷有效性[J]. 植物生态学报,2022,46(7):811-822
[30]ZHANG H,SHI L,WEN D,et al. Soil potential labile but not occluded phosphorus forms increase with forest succession [J]. Biology and Fertility of Soils,2015,52(1):41-51
(责任编辑 刘婷婷)
猜你喜欢
草地学报(2022年11期)2022-12-01
草食家畜(2021年5期)2021-03-27
草原与草业(2020年1期)2020-04-09
草地学报(2017年6期)2017-03-24
江苏农业科学(2016年8期)2017-02-15
红土地(2016年3期)2017-01-15
现代经济信息(2016年7期)2016-05-19
天津农业科学(2016年4期)2016-04-20
海峡姐妹(2015年9期)2015-02-27
杭州科技(2014年3期)2014-02-27
