
连云港南部近岸海域游泳生物群落结构调查
2024-01-24 09:54祝超文张虎胡海生贲成恺袁健美葛跃浩
水产养殖 2024年1期
祝超文,张虎*,胡海生,贲成恺,袁健美,葛跃浩
(1.江苏省海洋水产研究所,江苏 南通 226001;2.国家海洋局南通海洋环境监测中心站,江苏 南通 226001)
连云港海域位于海州湾渔场中部[1-2],海域内水质优良,具有丰富的生物资源和较高的生产力[3-4]。近年来,掠夺式的捕捞方式和涉海工程建设导致的环境污染问题,造成该海域内多数游泳生物的栖息地环境被严重破坏。鱼类、甲壳类和头足类等游泳生物是渔业资源的重要组成部分[5]。结合多个类群,分析游泳生物的群落结构组成及其变化,是渔业生态学研究的重要内容,也是渔业资源科学管理和合理利用的参考依据[6-8]。
冗余理论是解释生物群落稳定性的理论之一,目前多用于分析大型底栖动物群落的稳定性,对于游泳生物群落稳定性的研究较少[9]。在生态学中,冗余度是指一个物种和成分的失效,不会造成系统功能的失效[10]。群落特征可以由许多种类交替决定,群落中或多或少存在部分的种类冗余[11-12]。群落稳定性与生物多样性有关,食物网越复杂,群落结构越稳定,对外来干扰的抵御能力也越强,可更好地抵御外来干扰和维持其功能相对稳定性[13]。目前国内学者针对海州湾海域渔业资源做过许多相关研究,其中关于游泳生物群落的结构及其变化的研究,多集中在种类组成、资源量、多样性指数、不同捕捞方式、不同营养级和生物量谱的时空变化等[14-18]。张芮等[19]在2011 年、2013—2016 年春、秋2 季,研究了海州湾及其邻近海域鱼类群落结构冗余度,发现决定调查海域的鱼类群落结构特征的冗余度较低,当主要种类受损时,群落结构会发生较大变化,鱼类群落结构稳定性较低。现于2016—2020年春季,对连云港南部近岸海域游泳生物群落结构进行调查,运用多元统计分析和结构冗余度分析方法,分析不同年份间游泳生物群落结构变化,结合广义可加模型(GAM),解释造成游泳生物群落差异的主要影响因素,以期为深入研究连云港南部近岸海域游泳生物资源可持续发展,以及该海域的资源养护提供基础资料。
1 材料与方法
1.1 调查方法
于2016—2020 年春季(5 月),在江苏省连云港市南部近岸海域开展5 个航次渔业资源调查。根据项目调查和实际海况,设9 个固定站点,站位号1—9,见图1。采用底拖网调查方式,调查船主机功率为202 kW;网具主尺度125.3 m×59.0 m,网囊长6.1 m、网孔径20 mm,基本浮沉力配备90 kg;网板为钢质矩形1.6 m×1.0 m。作业期间,平均渔船拖速3.0 kn,站位拖曳时间设定1 h,在白天进行作业调查,同时测定水深、温度和盐度等指标。
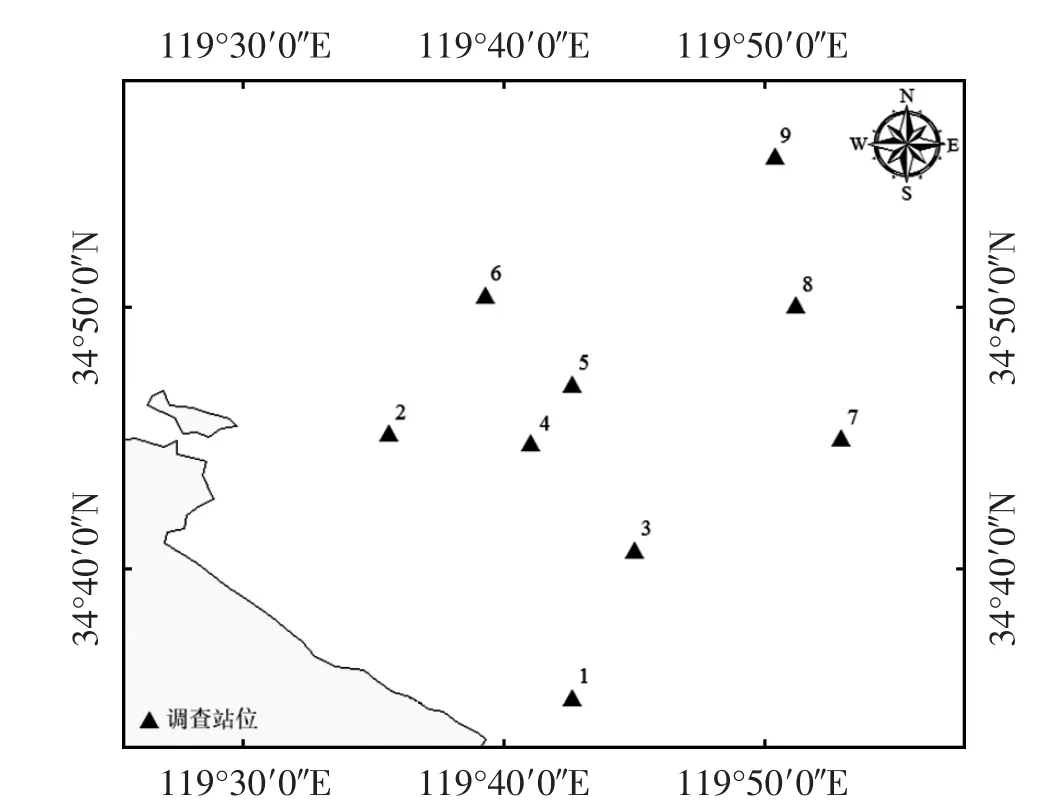
图1 连云港南部近岸海域调查站位
1.2 指标计算
1.2.1 渔业资源密度
渔业资源密度计算采用扫海面积法[20],按照《建设项目对海洋生物资源影响评价技术规程》(SC/T 9110—2007),各调查站位资源密度(质量和尾数)的计算公式为:
式中:Di——第i站的渔业资源密度,ind./km2或kg/km2;
Ci——第i站平均每小时拖网渔获量,ind./h 或kg/h;
ai——第i站位每小时网具取样面积,km2/h;
q——网具捕获率。参考《海洋渔业资源调查规范》(SC/T 9403—2012),底层鱼类、虾蟹类、头足类q取0.5,近底层鱼类取0.4,中上层鱼类取0.3。
1.2.2 生物多样性
游泳生物多样性指数,根据Shannon-wiener 指数(H')[21]、丰富度指数(d)[22]、均匀度指数(J)[23]计算,公式如下:
式中:pi——第i种的数量,ind.;
N——测站所有种类总数量,ind.;
S——出现的种类数。
1.2.3 优势种
游泳生物优势种,依据Pinkas 等[24]的相对重要性指数(IRI)计算,公式如下:
式中:N——某一种游泳生物的个数占总数的百分比,%;
W——某一种游泳生物的质量占总质量的百分比,%;
F——某一种出现的站次数占调查总站数的百分比,%。
采用传统划分标准定义IRI:IRI<10 时,为少见种,100>IRI≥10 时,为常见种,1 000>IRI≥100 时,为重要种,IRI≥1 000 时,为优势种。
1.2.4 聚类分析和多维尺度分析
为了分析群落结构年变化,运用聚类分析的方法,比较不同年份不同站位群落的相似性。对不同年份春季游泳生物单位网次渔获数量,进行平方根转换,根据平方根转换的游泳生物单位网次渔获数量,计算各个站位的游泳生物种类组成Bray-Curtis 相似性系数矩阵,运用聚类分析(CLUSTER)和(指多维尺度分析)MDS 比较不同年份间的相似性。应用相似性分析(SIMPER)研究造成组间群落组成相似的典型种和造成年间差异的分歧种[25-26]。
1.2.5 结构冗余度分析
应用PRIMER 软件包中的BVSTEP 程序,实现筛选响应单元的过程。对2016—2020 年春季调查数据,进行标准化处理,运用单网次游泳生物数量进行计算。响应单元一般定义为:满足p>0.95且包含最少种类数的子种类组合。响应单元的确定,采用类似于多元逐步回归分析的“向前选择/向后剔除”方法,逐步搜索计算,直到Spearman 秩相关系数p>0.95 时为止。当筛选出第一个响应单元后,排除这些种类,然后重复上述过程,搜索是否还有满足要求的响应单元,直到不能满足p>0.95 为止。用最后筛选的响应单元的个数,来衡量群落结构冗余度水平[27]。
1.2.6 GAM 构建
GAM 能拟合响应变量和解释变量之间的非线性关系,各环境因子经Pearson 相关性分析后,初步选择的因子包括:年份、温度(SST)、盐度(SSS)、深度(DEP)、溶解氧(DO)和pH 值[28]。以各站位资源质量密度为响应变量,以相关因子作为解释变量,利用GAM 模型对连云港南部近岸海域资源量与相关环境因子进行回归分析[29],其表达式为:
式中:Y——响应变量;
a——拟合模型中的截距;
xi——第i个解释变量;
ε——误差项;
si——样条平滑函数。
冗余度和多样性的计算,在Primer 5.0 中进行,GAM 通过R 语言软件中的“mgcv”包实现,站位图采用Arcmap 10.2 软件绘制。
2 结果与分析
2.1 物种组成
调查海域2016—2020 年5 个航次共捕获各种游泳生物85 种,其中鱼类54 种、虾类16 种、蟹类10种、头足类5 种;游泳生物物种数分别为52、55、42、48 和39 种。该海域5 年间游泳生物种类数量呈现下降趋势。鱼类在各类游泳生物种类组成中所占比重较高,其次是虾类,蟹类和头足类比重相对较低;主要种类减少类群为鱼类和蟹类,鱼类主要减少品种为许氏平(Sebastes schlegelii)等;蟹类主要减少品种为狭颚绒螯蟹(Eriocheir leptognathus)等,见图2。
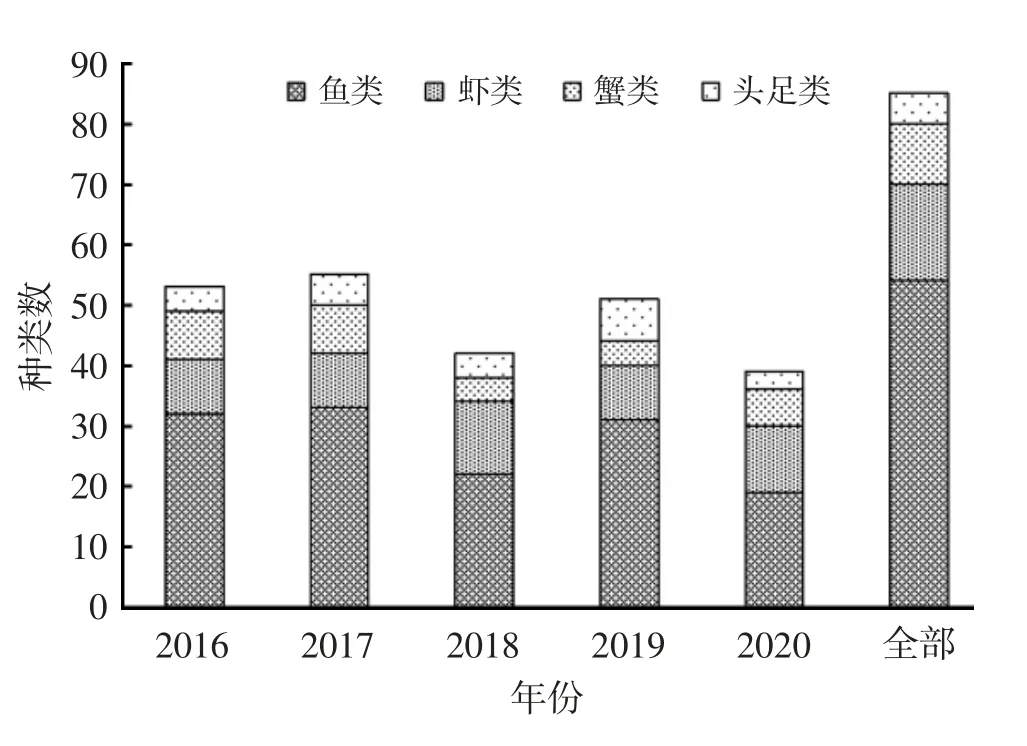
图2 2016—2020 年春季连云港南部近岸海域游泳生物物种组成
2.2 游泳生物资源密度年变化
根据扫海面积法计算结果分析,除2017 年数据波动较大外,其余年份资源数量密度和质量密度呈上升趋势;2017 年的资源数量密度和质量密度最高。在各类群资源密度方面,鱼类尾数密度和质量密度占比相对较高,为游泳生物群落的主要类群,其次为虾类和蟹类,头足类资源尾数密度相对较低。其中,2017 年赤鼻棱鳀(Thrissa kammalensis)、2018 年口虾蛄(Oratosquilla oratoria)和2020 年三疣梭子蟹(Portunus trituberculatus)资源尾数密度和质量密度,占当年总资源尾数密度和质量密度的比例较高,单品种资源数量的暴发,导致个别年份游泳生物资源尾数密度和质量密度,出现较为显著的差异(表1)。
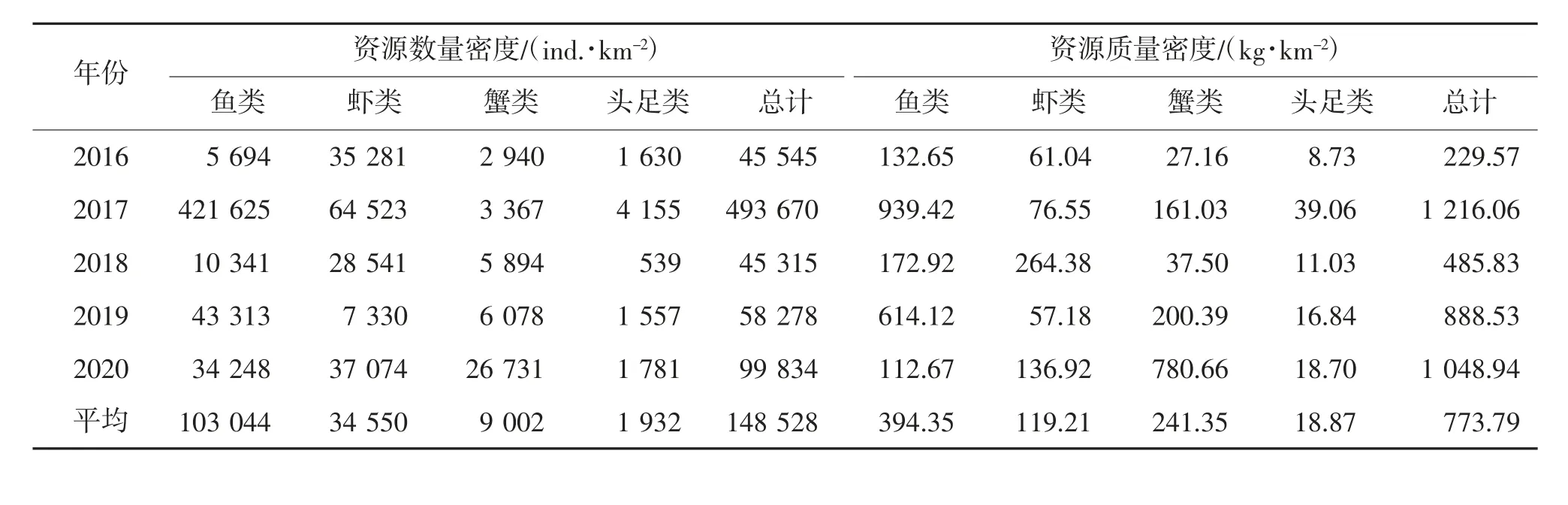
表1 调查站位游泳生物各种类资源密度
2.3 资源优势种
2016—2020 年春季连云港南部近岸海域游泳生物资源优势种及其年间变化见表2。由表2 可见,2016—2020 年连云港南部近岸海域春季优势种主要有9 种。其中2017 年优势度最高的赤鼻棱鳀资源数量密度,占当年总数量密度的81%;2018 年春季主要优势种口虾蛄资源数量密度占总数量密度的44%;2020 年春季优势度最高的三疣梭子蟹资源数量密度,占当年总数量密度的23%。口虾蛄和三疣梭子蟹为5 年间的主要优势种,戴氏赤虾(Metapenaeopsis dalei)的优势度在逐年下降,尖海龙(Syngnathus acus)和日本鼓虾(Alpheus japonicus)的优势度呈上升趋势。
2.4 生物多样性指数
2016—2020 年春季调查海域多样性指数分布见表3。由表3 可见,调查海域2016—2020 年H'为0.19~3.95;d为0.59~3.56;J为0.05~0.91。总体上,2016—2020 年连云港南部近岸海域生物H'呈下降趋势,与物种数变化趋势相似。
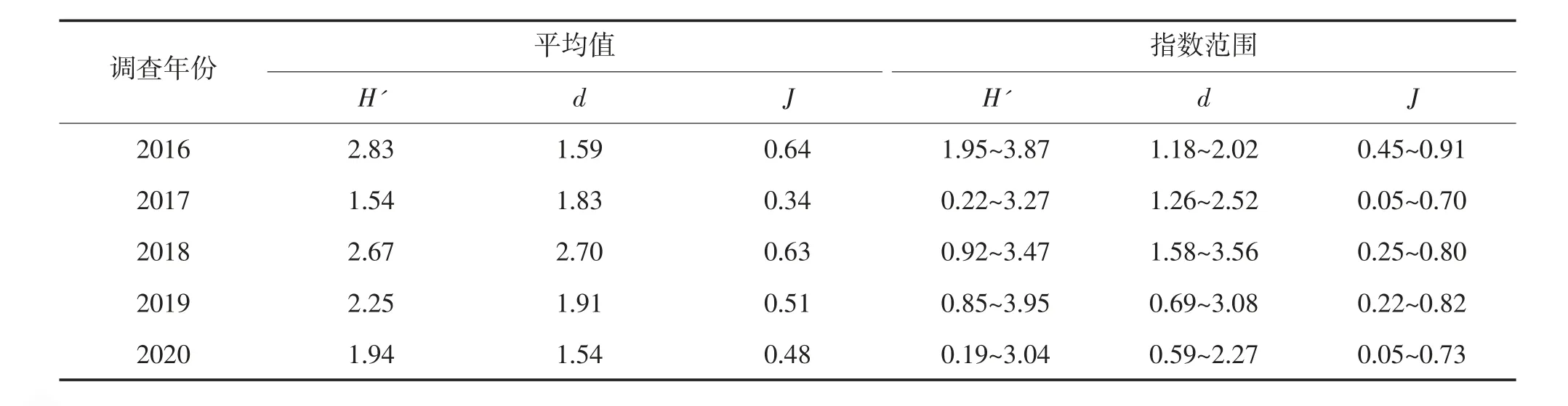
表3 2016—2020 年春季连云港南部近岸海域游泳生物H'、d 和J
2.5 游泳生物群落结构年变化
聚类分析结果表明,2016—2020 年调查海域游泳生物群落可分为2 个站位组(图3)。站位组1 包含11 个站位,站位组2 包含34 个站位。从站位年份上来看,站位组1 主要为2017 年调查站位,站位组2 为其余年份的站位。表4 列出了调查海域春季游泳生物群落2 个站位组的组间典型种和分歧种。站位组1 的典型种包括赤鼻棱鳀和日本鳀,对组间平均相似性的贡献达到80.70%。站位组2 的典型种为戴氏赤虾、口虾蛄、双斑蟳(Charybdis bimaculata)、火枪乌贼(Loligo beka)和尖海龙,对组间相似性的贡献达到75.56%。2 组间的分歧种主要为戴氏赤虾、口虾蛄、尖海龙、赤鼻棱鳀、日本鳀和康氏侧带小公鱼,对组间相异性贡献率达到了53.20%。

表4 连云港南部近岸海域春季游泳生物群落各组间典型种和分歧种
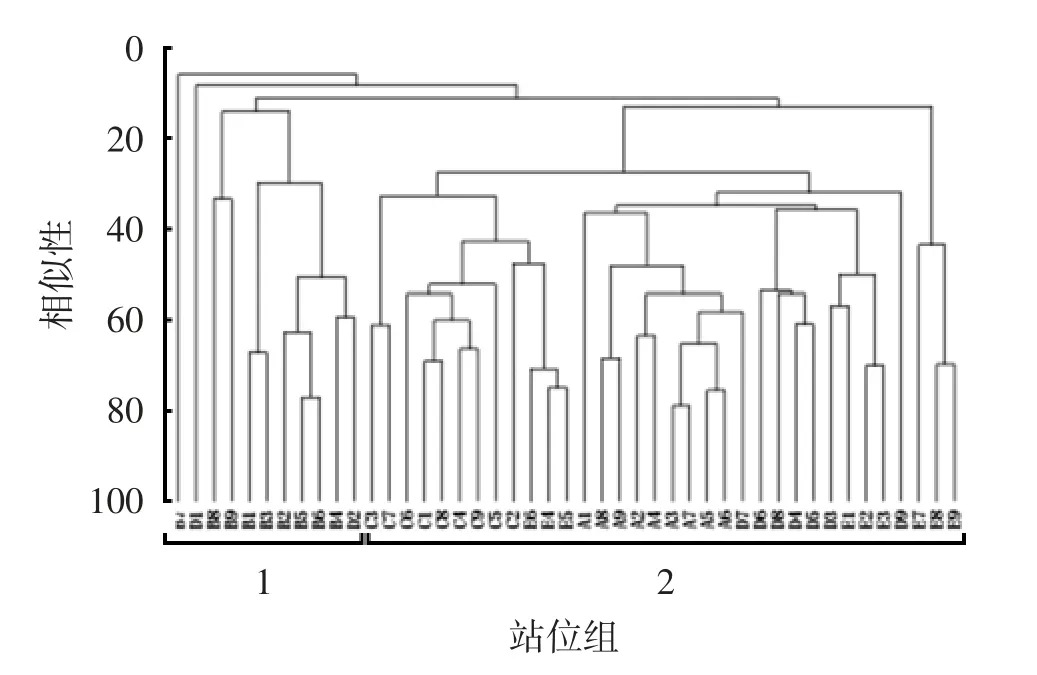
图3 2016—2020 年调查海域游泳生物群落结构变化的聚类分析
2.6 结构冗余度
2016 年春季连云港南部近岸海域游泳生物群落多维尺度分析(MDS)图见图4(a)(b)(c)(d)。由图4可见,连云港南部近岸海域2016 年春季游泳生物群落响应单元1 包括6 个种类,响应单元1 对应的MDS 图如图4(b),与4(a)的Spearman 秩相关系数p=0.955;响应单元2 包括5 个种类,该响应单元的MDS 图为4(c),与图4(a)的匹配相关系数p=0.952;响应单元3 包括12 个种类,该响应单元的MDS 图为4(d),与图4(a)的匹配相关系数p=0.893。因此,可以筛选出2 个相关系数p>0.950 的响应单元,2016年调查海域游泳生物群落结构冗余度水平较低。同样的方法处理得到,2017 年春季筛选出1 个相关系数p>0.950 的响应单元;2018 年春季筛选出1 个相关系数p>0.950 的响应单元;2019 年春季筛选出2个相关系数p>0.950 的响应单元;2020 年春季筛选出1 个相关系数p>0.950 的响应单元。调查海域2016—2020 年春季游泳生物群落结构冗余度,除2016 和2019 年有2 个响应单元外,其余仅有1 个响应单元,5 年间,调查海域游泳生物群落结构的冗余度水平都较低。
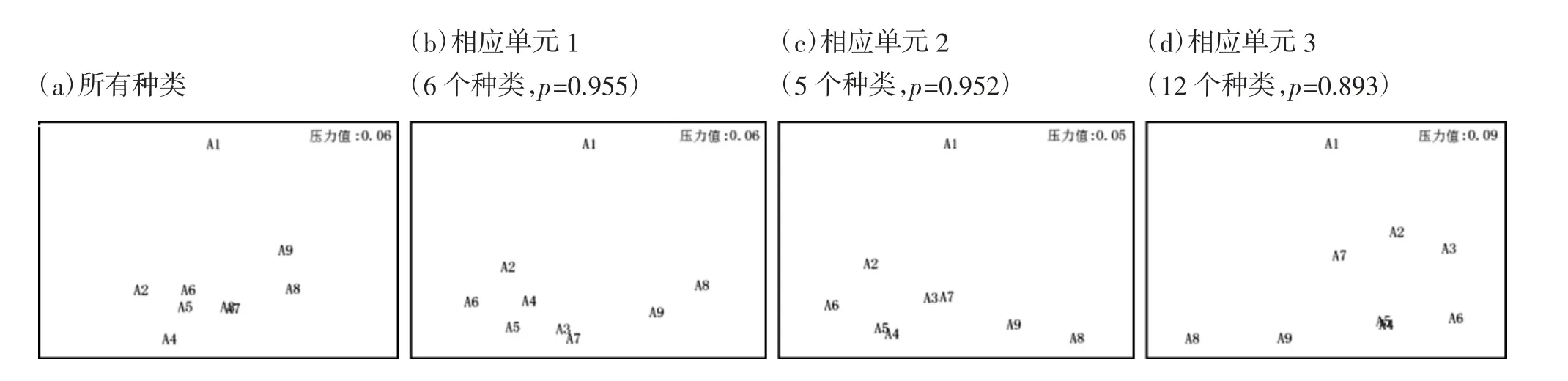
图4 2016 年春季连云港南部近岸海域游泳生物群落MDS
2.7 不同因子对调查海域资源量的影响
通过GAM 模型,拟合2016—2020 年各站位资源质量密度与各影响因子之间的关系。采用AIC 准则进行因子的筛选,根据赤池信息准则(AIC)最小原则,获得最优GAM 模型(表5、6),模型最终的表达式为:
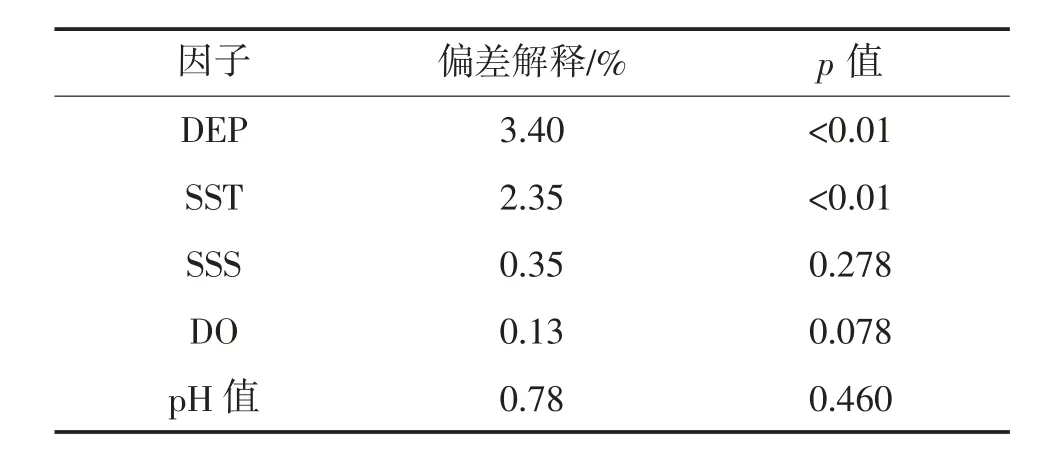
表5 游泳生物资源质量密度的环境单因子广义可加模型的显著性变量的检验
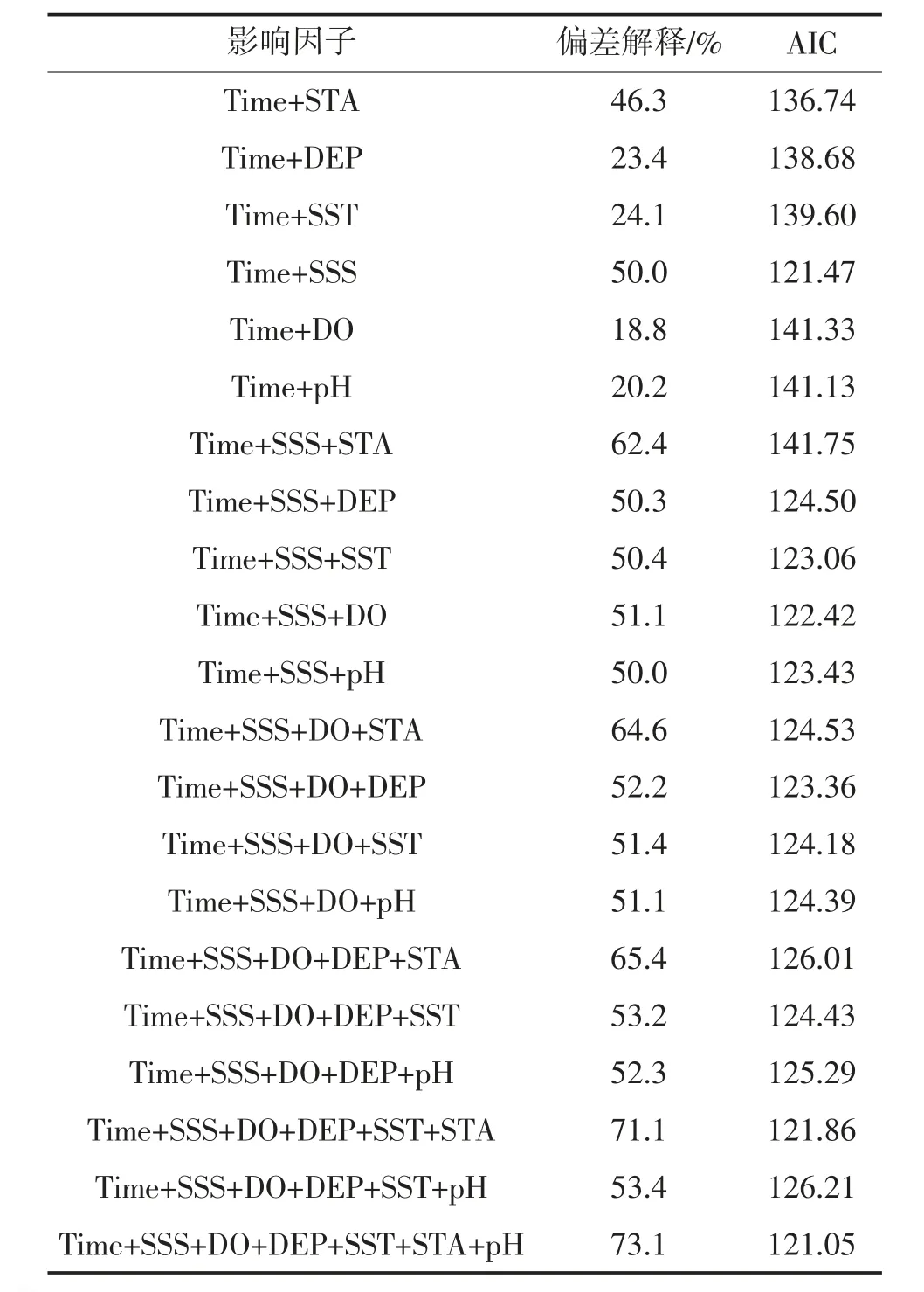
表6 GAM 模型变量筛选及影响因子的参数
CUPE 为单位捕捞努力量渔获量;Time 为年际因子;STA 为站位。模型包含5 个环境因子,模型AIC 值为121.05,可解释73.1%的总偏差,深度和温度与资源质量密度存在极显著关系(p<0.05)。
不同因子对连云港南部近岸海域资源量的影响见图5(a)(b)。由图5 可见,DEP 为3.0~3.8 m 时,其资源质量密度随着海水深度的增加而减少;当DEP 达到13 m 时,其资源质量密度随水深而小幅度增加。温度效应图显示,为12.8~18.0 ℃,其资源质量密度随着温度的上升而增加;当SST>18 ℃时,资源质量密度随着温度的升高而减少。
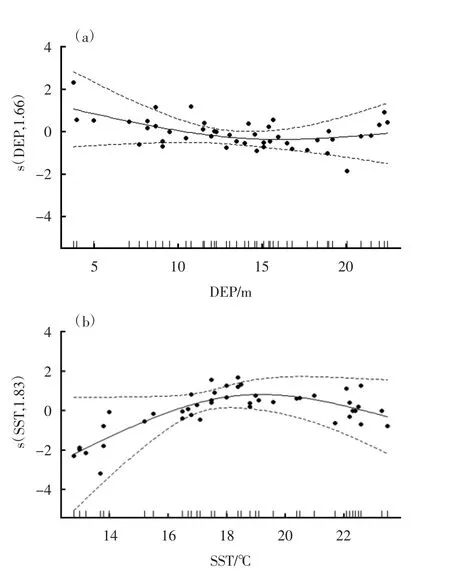
图5 不同因子对连云港南部近岸海域资源量的影响
3 讨论
调查海域位于海州湾中南部,是暖温带南部和北温带的过渡地带,具有海洋性季风气候的特点,是黄海海域重要的渔场之一[1],也是多种经济鱼类的产卵场和索饵场[30]。文献[14-15]研究表明,在21 世纪初对黄海北部连云港海域春季调查中,共发现游泳生物60 种;本次调查中,从2016 年春季共捕获游泳生物种类数52 种,到2020 年春季只捕获到游泳生物39 种,2017 和2018 年春季捕获到的游泳生物种类有小幅度波动,但整体呈下降趋势,其中虾类、蟹类和鱼类变化的幅度较大。研究[31-32]表明,当人为因素对某一时期或某一区域的群落干扰较大时,部分抗干扰能力较差的种类资源密度会下降或者消失。高强度的捕捞、环境污染以及栖息地被破坏等人为和自然因素的干扰,可能是导致连云港南部近岸海域自然群落种类数量减少的重要原因之一。
结合GAM 模型结果分析,温度和水深对调查海域资源质量密度的影响较小,而年间的变化是影响资源质量密度的主要因素。聚类分析中,2 站位组环境因素主要差异在于水深和温度,站位组1 的平均深度和温度要高于站位组2。2 站位组游泳生物群落的典型种和分歧种,基本上为戴氏赤虾、口虾蛄、三疣梭子蟹、赤鼻棱鳀、日本鳀和尖海龙等对环境变化干扰适应性较强的品种,当这些品种面对外界干扰时,适应性较强的品种多为r-选择物种,是所谓的“机会主义者”。本调查中,底层种类例如三疣梭子蟹和口虾蛄等,可能因为对海底生境变化适用性强,出现了资源密度增加的现象。游泳生物种类中的赤鼻棱鳀和小黄鱼等短距离洄游性鱼类,对于摄食环境和温度的变化敏感性,高于洄游品种,温度升高,有利于典型种中赤鼻棱鳀(暖水性)和小黄鱼(暖温性)生长[33],当外界环境不适宜时,其会向适宜的海域移动。戴氏赤虾适宜生存温度为13~25 ℃[34],连云港近岸春季水温适宜其生存繁殖。口虾蛄和三疣梭子蟹等属于广温广盐品种,在黄海各海域均有分布,对外界干扰抵抗力较强。所以这2 个种类种群数量,在生存环境适宜的年份产生较大的波动,对年际间相异性贡献率也最高。因此,连云港南部近岸海域游泳生物群落结构的差异,主要是由少数种类数量分布变化造成的。
游泳生物群落通常与生物多样性联系在一起,食物网越复杂,对抗外来干扰的能力越强。食物网发生不稳定,是由于某物种减少或灭绝,没有替代的物种而引起的。食物网内的物种和种群个体间都存在一定的营养级结构,是群落能够抵抗干扰和维持相对稳定的原因[35]。本调查中,2016、2018 和2019年的生物多样性和均匀度水平较2017 和2020 年高。在群落中多样性水平高的情况下,物种分布较均匀,群落结构就更加稳定。
本调查中,该海域2016—2020 年春季游泳生物群落结构冗余度都偏低,除了2016 和2019 年有2 个ρ>0.95 的响应单元外,其他年份均只有1 个响应单元,表明决定游泳生物群落结构特征的可替代种类较少。当主要游泳生物种类受损时,群落结构会发生较大的变化,群落结构变得不稳定。不同海域不同生物群落或者相同海域不同时空变化的结构冗余度水平,都具有一定的变化。张芮等[19]对2011 年、2013—2016 年春季和秋季海州湾鱼类群落研究表明,各季节均存在1 个响应单元,与本调查游泳生物群落结构冗余度相近;1985—2002 年,南黄海冬季鱼类群落结构存在2 个响应单元,冗余度水平都不高[36-37]。也有研究发现,大型底栖动物群落冗余度水平通常远高于海洋游泳生物的群落冗余度水平。例如,Morlaix 海湾大型底栖动物群落存在5 个响应单元;Northumberland 海岸大型底栖动物群落存在3~4 个响应单元[38];群落结构冗余度水平越高,营养级中相近种类的生态位越分化,对空间资源的利用更趋向于互补,结构冗余度水平的提高,可以增加群落结构的稳定性和抵抗外来干扰的能力[34]。今后,应加大对近岸海域的叶绿素a、悬浮物和重金属元素等环境因子的监测,并应用多物种的空间模型,为解决资源衰退问题提供数据支撑,保障渔业资源的可持续发展。
猜你喜欢
纺织报告(2022年8期)2022-08-25
今日农业(2021年8期)2021-11-28
水上消防(2021年3期)2021-08-21
今日农业(2020年23期)2020-12-15
今日农业(2020年22期)2020-12-14
人大建设(2019年5期)2019-10-08
山西建筑(2017年29期)2017-11-15
黑龙江交通科技(2017年7期)2017-09-20
黑龙江交通科技(2017年10期)2017-03-01
黑龙江交通科技(2016年11期)2016-03-11
