
疏勒河源区高寒草甸中型土壤动物群落特征及对土壤水分变化的响应
2024-01-18 10:26:54刘继亮赵文智王永珍冯怡琳林永一陈生云
冰川冻土 2023年6期
刘继亮, 赵文智, 王永珍, 冯怡琳, 林永一, 陈生云
(1. 中国科学院 西北生态环境资源研究院 临泽内陆河流域研究站,甘肃 兰州 730000;2. 中国科学院大学,北京 100049; 3. 宁夏大学,宁夏 银川 750021)
0 引言
高寒草甸是中国重要的草地类型,主要分布在青藏高原及其毗邻区域的中高海拔区,它是中国西部重要的生态安全屏障[1-2]。高寒草甸面临气候变化和人类活动干扰的双重威胁,生态系统结构和功能不断降低,严重威胁区域生态系统的安全与稳定,这在降水稀少的疏勒河源区表现得尤为明显[2-4]。放牧等人类活动导致高寒草甸植被盖度及多样性降低,植被退化会引起多年生和季节性冻土出现不同程度的退化,冻土退化强烈影响高寒草地生态系统的生态服务功能[5-7]。目前,围绕疏勒河源区冻土退化过程中水文、气候、土壤、植被及微生物多样性已经陆续开展了一系列研究,确定了高寒草甸地上及地下生态系统关键要素对全球变化的响应规律[2,7-10]。土壤动物是地下生态系统的关键组分,它与微生物组成的土壤微小食物网和碎屑食物网控制着高寒草甸凋落物分解和养分循环过程,通过地上和地下生态系统的互馈影响地上生态系统过程[11-13]。然而,我们对高寒草甸土壤动物物种组成、多样性及其功能的认识还很有限,不清楚高寒草甸退化过程中土壤动物的组成及多样性变化规律,更不清楚它们中一些关键土壤动物类群对高寒草甸退化的响应模式。
高寒草甸退化过程中植物多样性及初级生产力不断降低,土壤水分及养分含量逐步下降、土壤出现退化,从而威胁草地生态系统的安全与稳定。地表和土栖土壤动物多样性对高寒草甸退化的响应模式不同并存在地域差异[14]。地表节肢动物、土栖节肢动物和线虫及微生物对高寒草甸退化的敏感程度不同,适度的放牧可以提高高寒草甸土壤动物多样性,与植被初级生产力和多样性及土壤肥力对草地退化的响应相近,高寒草甸生态系统生物多样性及其功能对放牧强度的响应满足“中度干扰理论”[4,15-18]。此外,一些研究还发现土壤水分有效性影响高寒草甸植被初级生产力及多样性对放牧强度的响应,充足的土壤水分可以缓解牲畜践踏对高寒草甸植被生长的影响[19]。高寒草甸退化过程中土壤水分含量随之降低,地表节肢动物多样性会随之增加,土壤水分还改变了地表节肢动物对围封禁牧和自由放牧的响应模式[20]。土壤水分是高寒草甸地表和土栖土壤动物分布的重要限制因子,过高的土壤水分含量会抑制土壤节肢动物的活动,适度降低土壤水分含量有利于节肢动物的生存和繁殖,土壤水分在高寒草甸土壤生物多样性维持方面扮演重要的角色[21-23]。鉴于此,笔者以祁连山西段的疏勒河源区为研究区,解析土壤动物群落中参与凋落物分解的主要类群中型土壤动物多样性对该区高寒草甸退化的响应规律,确定高寒草甸退化过程中土壤水分对中型土壤动物群落特征的影响。
1 材料与方法
1.1 研究区概况
疏勒河源区位于祁连山国家公园南坡西段,青海省海西藏族自治州天峻县苏里乡境内,是中国西部高寒草甸的主要分布区,也是重要的生态安全屏障和水源涵养区。疏勒河源区属于多年冻土和季节性冻土区,海拔在3 800~4 100 m 之间;该区气候干旱,年降水量为321.6 mm,降水主要集中在5—9月,这5 个月降水量占全年降水量的88.5%。年均气温为-4.5 ℃,月最高气温为6.5 ℃出现在8月,月最低气温为-17.8 ℃出现在1 月。前期研究已经发现疏勒河源区高寒草甸植被盖度介于16.3%~88.7%之间,莎草科青藏苔草(Carex moorcroftii)、小苔草(C. parva)、高山蒿草(Kobresia pygmaea)、线叶蒿草(K. capillifolia)、藏蒿草(K.tibetica)和禾本科紫花针茅(Stipa purpurea)等是该区主要的植物种,气候变化引起的冻土退化和放牧强度改变高寒草甸植被盖度及物种多样性变化[9,24]。疏勒河源区藏野驴(Equus kiang)、藏狐(Vulpes ferrilata)、赤狐(V.vulpes)、普通鵟(Buteo buteo)、秃鹫(Aegypius monachus)、纵纹腹小鸮(Athene noctua)、棕颈雪雀(Pyrgilauda ruficollis)、旱獭(Marmota bobak)和鼠兔(Ochtonaspp.)等是该区常见的脊椎动物;家养食草动物主要为牦牛,牲畜养殖以自由放牧为主。高寒草甸土壤类型为栗钙土,低洼处沼泽化草甸土壤发育为潜育土,高寒草甸退化区土壤沙化现象明显,部分区域出现大范围黑土滩[1]。
1.2 试验设计、样品采集及处理
高寒草甸冻土退化引起沼泽化草甸向草甸、草原化草甸和沙化草甸演替,本文利用空间梯度代替时间序列的方法在疏勒河源区选取沼泽化草甸、草甸、草原化草甸和沙化草甸4 种典型的高寒草甸生境类型构成冻土退化及放牧扰动驱动的高寒区植被退化演替梯度[24]。每种高寒草甸生境依据微地形和植被变化及放牧强度变化等选择4 个地点,每个地点选择2 500~3 000 m2的范围内选择5 个1 m×1 m 的区域作为中型土壤动物样品采集区,每个土壤动物样品采集区采集1 个10 cm×10 cm×10 cm 土壤样品带回室内,利用改进干漏斗分离器分离中型土壤动物样品。高寒草甸土壤动物多样性存在明显的季节变异,夏季土壤动物数量和物种数较低,秋季土壤动物的数量和物种数较高[25]。2021年在6月中旬和10 月上旬采集0~10 cm 土壤样品,土壤动物样品连续分离120 h(5 d)。同时,在每个土壤动物样品采集区利用土钻采集0~10 cm 土壤样品带回室内使用烘干法测定土壤含水量。收集到的土壤动物样品保存在75%的酒精溶液中用于种属信息鉴定及数量统计。收集的中型土壤动物样品参照相关分类资料至科或亚目,部分动物样品鉴定至属种或形态种或直接测量获取其功能性状信息[26]。螨类样品中大量幼体无法鉴定至科,螨类、跳虫和昆虫在科或亚目的水平统计个体数量计算群落结构指数和进行相似性分析。
1.3 数据处理
2021 年6 月和10 月采集的中型土壤动物样品合并统计,统计中小型土壤动物群落中优势类群(相对多度>10%)、常见类群(相对多度在1%~10%之间)和稀有类群组成(相对多度<1%)。计算6 月和10月中型土壤动物群落密度(每平方米采集的动物个体数)、类群数(每平方米采集的动物类群数)和多样性指数(Shannon-Wiener index),统计分析使用Past 4.01 软件包。6 月和10 月中型土壤动物合并统计,计算中型土壤动物中螨类与跳虫密度的比值(A/C)和土壤动物QBS-ar 指数可以反映土壤动物群落结构及土壤质量变化,是在科或目的水平基于不同中型土壤动物栖居类型赋值计算的土壤动物群落指数,值越高说明土壤质量越好[23]。利用已有中型土壤动物EMI值赋值表计算4种高寒草甸中型土壤动物QBS-ar 指数[27]。采用二因素方差分析比较不同生境中型土壤动物群落和主要类群密度,利用单因素方差分析比较A/C 和QBS-ar指数差异,统计分析使用SPSS 21.0 软件包。在此基础上,利用曲线拟合分析中型土壤动物群落及QBS-ar 指数与表层土壤水分含量的关系,统计分析使用SPSS 21.0软件包。
利用非度量多维尺度(Non-metric multidimensional scaling, NMDS)和多元方差(Permutational multivariate analysis of variance, PERMANOVA)分析6月和10月4种高寒草甸生境中型土壤动物群落组成差异,然后利用相似性百分比(Similarity percentage analysis, SIMPER)分析确定4 种高寒草甸生境中型土壤动物群落的平均相异性和主要动物类群的贡献率,统计分析使用Past 4.01 软件包。NMDS 排序结果采用胁强系数(Stress)衡量排序分析结果的优劣,其中Stress<0.01,完全可信;0.01<Stress<0.05,可信的;0.05<Stress<0.1,基本可信;0.1<Stress<0.2,部分信息不可信;0.2<Stress<0.3,不可信[28]。
2 结果与分析
2.1 高寒草甸中型土壤动物物种组成及数量变化
2021 年6 月和10 月,在沼泽化草甸、草甸、草原化草甸和沙化草甸4 种生境利用干漏斗法共采集7 669 头31 类中型土壤动物,甲螨亚目、等节跳科和辐螨亚目是优势中型土壤动物类群;革螨亚目、长角跳科、球角跳科、长蝽科和蚧科是常见中型土壤动物类群;稀有中型土壤动物类群有23 个,它们的个体数占2.93%。沼泽化草甸生境捕获1 559 头17类中型土壤动物,甲螨亚目、等节跳科、辐螨亚目、革螨亚目、长角跳科、球角跳科和圆跳科是主要的中型土壤动物类群;草甸生境捕获到963 头13 类中型土壤动物,甲螨亚目、等节跳科、辐螨亚目、革螨亚目、长角跳科、球角跳科和蚧科是主要的中型土壤动物类群;草原化草甸生境捕获到4 278 头21 类中型土壤动物,甲螨亚目、节跳科、辐螨亚目、革螨亚目、长角跳科、球角跳科、长蝽科和蚧科是主要的中型土壤动物类群;沙化草甸生境捕获到869 头16类中小型土壤动物,甲螨亚目、等节跳科、辐螨亚目、革螨亚目、球角跳科、软蜱科、长蝽科和蚧科是主要的中型土壤动物类群。
2.2 高寒草甸中型土壤动物群落结构变化
2021年6月和10月4种高寒草甸生境类型中型土壤动物PERMANOVA 分析结果表明,采样期和生境类型对中型土壤动物群落组成均有显著影响(F=19.31,P<0.001;F=11.99,P<0.001),采样期和生境类型对高寒草甸又有显著的交互影响(F=4.89,P<0.001)。6 月沼泽化草甸、草甸、草原化草甸和沙化草甸4种生境中型土壤动物群落组成明显不同(F=7.89,P<0.001),4 种高寒草甸间中型土壤动物群落组成均明显不同[图1(a)]。SIMPER 分析结果表明,沼泽化草甸和草甸生境中型土壤动物群落的平均相异性为70.0%,甲螨亚目、等节跳科和辐螨亚目解释了2 种生境中型动物群落差异的75.4%;沼泽化草甸与草原化草甸和沙化草甸中型土壤动物的平均相异性为73.9%和78.7%,甲螨亚目、等节跳科和辐螨亚目解释了它们之间群落差异的60.4%和65.4%。草原化草甸和沙化草甸中型土壤动物的平均相异性为74.8%,甲螨亚目、等节跳科、辐螨亚目、革螨亚目和球角跳科解释了它们之间中型土壤动物群落差异的83.7%。10 月沼泽化草甸、草甸、草原化草甸和沙化草甸4种生境中型土壤动物群落组成明显不同(F=9.00,P<0.001),4种高寒草甸间中型土壤动物群落组成均明显不同
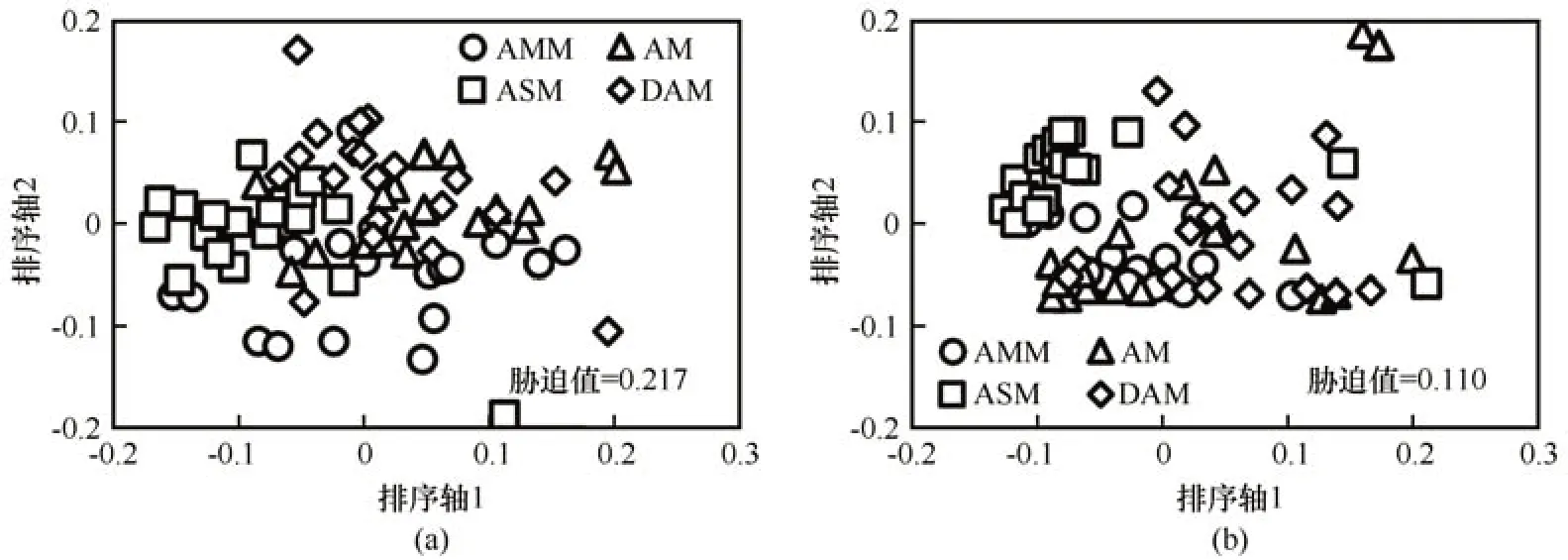
图1 疏勒河源区6月(a)和10月(b)沼泽化草甸(AMM)、草甸(AM)、草原化草甸(ASM)和沙化草甸(DAM)中型土壤动物群落组成NMDS排序图Fig. 1 Non-metric multidimensional scaling (NMDS) plots indicating 2-dimensional distances of soil mesofauna community of the alpine marsh meadow (AMM), alpine meadow (AM), alpine steppe meadow (ASM), and desertification alpine meadow (DAM) in June (a) and October (b) in the Shule River headwaters
[图1(b)]。SIMPER 分析结果表明,沼泽化草甸和草甸生境中型土壤动物群落的平均相异性为64.3%,甲螨亚目和等节跳科解释了2 种生境中型动物群落差异的74.3%;沼泽化草甸与草原化草甸和沙化草甸中型土壤动物的平均相异性为69.5%和68.2%,甲螨亚目和等节跳科解释了它们之间群落差异的66.6%和71.6%。草原化草甸和沙化草甸中型土壤动物的平均相异性为81.8%,甲螨亚目、等节跳科和革螨亚目解释了它们之间中型土壤动物群落差异的78.4%。
疏勒河源区2 个采样期和4 种高寒草甸生境中型土壤动物群落结构参数的二因素方差分析结果表明,采样期对中型土壤动物密度和多样性指数有显著影响,生境类型对中型土壤动物密度、类群丰富度和多样性指数均有显著影响,采样期和生境类型对中型土壤动物密度、类群丰富度和多样性指数的交互影响较小(表1)。6 月疏勒河源区高寒草甸中型土壤动物密度显著低于10月,而中型土壤动物多样性指数变化相反,6 月和10 月中型土壤动物类群丰富度相差较小(图2)。6 月草原化草甸中型土壤动物密度和类群丰富度均显著高于其他生境,10月草原化草甸中型土壤动物密度和类群丰富度显著高于沼泽化草甸,沼泽化草甸中型土壤动物的密度和类群丰富度又显著高于草甸和沙化草甸;6 月和10 月草原化草甸中型土壤动物多样性指数均显著高于其他生境(图2)。

表1 采样期和高寒草甸类型对中型土壤动物群落结构和主要类群密度影响的二因素方差分析Table 1 The results of two-way ANOVAs for sampling periods and habitat types affected community structure and dominant groups of soil mesofauna in the Shule River headwaters
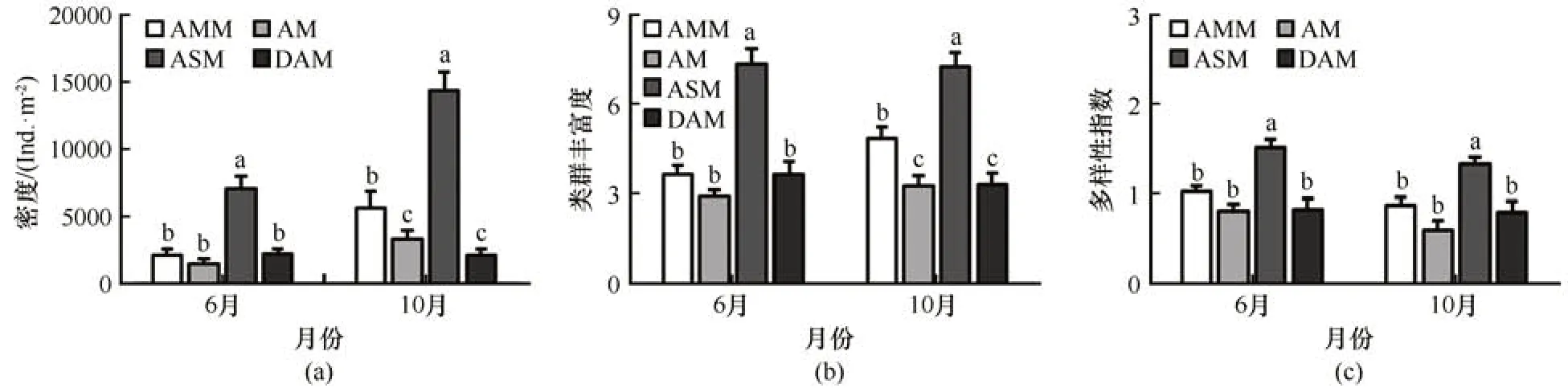
图2 6月和10月疏勒河源区沼泽化草甸(AMM)、草甸(AM)、草原化草甸(ASM)和沙化草甸(DAM)中型土壤动物密度(a)、类群丰富度(b)和多样性指数(c)Fig. 2 The density (a), groups richness (b), and Shannon-Wiener index (c) of soil mesofauna community in the alpine marsh meadow (AMM), alpine meadow (AM), alpine steppe meadow (ASM), and desertification alpine meadow (DAM)in June and October in the Shule River headwaters
中型土壤动物群落QBS-ar 指数能指示高寒草甸土壤质量变化,4 种高寒草甸生境中型土壤动物QBS-ar 指数存在显著差异(F=14.09,P<0.001)。中型土壤动物QBS-ar 指数变化趋势与中型土壤动物多样性指数一致,草原化草甸生境中型土壤动物QBS-ar 指数显著高于其他3 种生境,草原化草甸生境中型土壤动物QBS-ar指数是沼泽化草甸、草甸和沙化草甸生境的1.3 倍、1.6 倍和1.6 倍(图3)。此外,研究还发现沼泽化草甸中型土壤动物QBS-ar指数略高于草甸和沙化草甸生境(图3)。
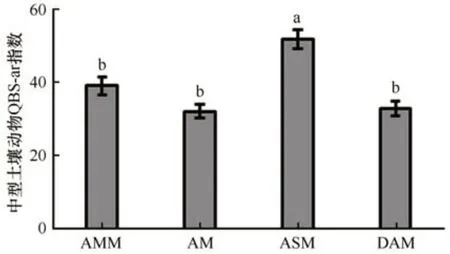
图3 疏勒河源区沼泽化草甸(AMM)、草甸(AM)、草原化草甸(ASM)和沙化草甸(DAM)中型土壤动物QBS-ar指数比较Fig. 3 The QBS-ar index of soil mesofauna community in the alpine marsh meadow (AMM), alpine meadow (AM),alpine steppe meadow (ASM), and desertification alpine meadow (DAM) in the Shule River headwaters
2.3 高寒草甸主要中型土壤动物类群密度变化
疏勒河源区蜱螨目和弹尾目密度的二因素方差分析结果表明,采样期对蜱螨目和弹尾目密度的影响较小,高寒草甸生境类型均对蜱螨目和弹尾目密度有显著影响,采样期和生境类型的交互对蜱螨目的密度也有显著影响(表1)。6 月草原化草甸蜱螨目的密度显著高于其他生境,10 月草原化草甸和沼泽化草甸蜱螨目的密度显著高于草甸和沙化草甸;6 月和10 月草原化草甸弹尾目的密度均显著高于其他生境(图4)。螨类和跳虫密度的比值(A/C)在4 种高寒草甸生境也存在明显变化(F=7.56,P<0.001),草原化草甸生境A/C 值显著低于其他草甸生境,沼泽化草甸、草甸和退化草甸A/C值是草原化草甸的7.5倍、12.0倍和7.3倍。
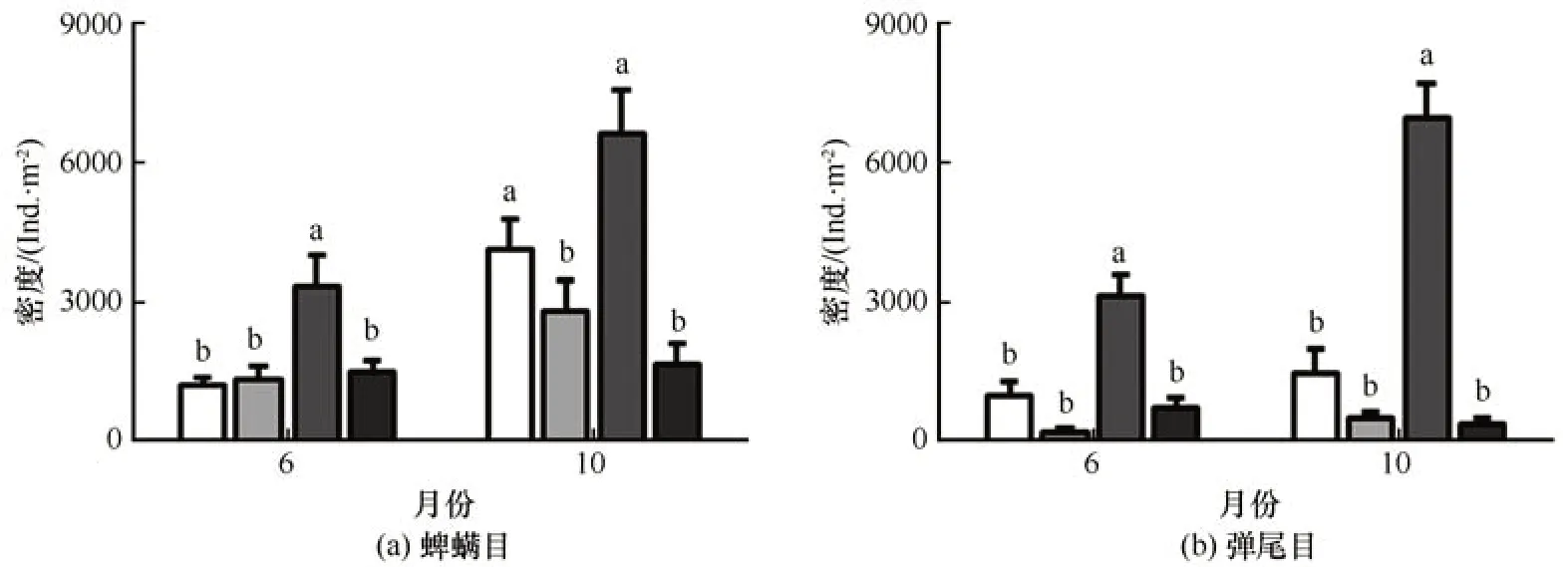
图4 6月和10月疏勒河源区沼泽化草甸(AMM)、草甸(AM)、草原化草甸(ASM)和沙化草甸(DAM)生境蜱螨目和弹尾目密度比较Fig. 4 The density of Acarina (a) and Collembola (b) of the alpine marsh meadow (AMM), alpine meadow (AM), alpine steppe meadow (ASM), and desertification alpine meadow (DAM) in June and October in the Shule River headwaters
2021 年6 月和10 月不同中型土壤动物对沼泽化草甸、草甸、草原化草甸和沙化草甸构成的退化梯度的响应模式不同。采样期对高寒草甸甲螨亚目、辐螨亚目、长角跳科、球角跳科和等节跳科的密度有显著影响(表1),10月甲螨亚目、长角跳科和等节跳科的密度均显著高于6 月,而辐螨亚目和球角跳科的密度与甲螨亚目等相反(图5)。高寒草甸生境类型对8 个主要的中型土壤动物密度有显著影响,而采样期和生境类型的交互仅对辐螨亚目、长角跳科、球角跳科和等节跳科的密度有显著影响(表1)。6 月和10 月革螨亚目、长蝽科和蚧科密度在4种高寒草甸变化趋势相近,6月草原化草甸革螨亚目、长蝽科和蚧科密度显著高于其他生境;10 月草原化草甸革螨亚目和蚧科密度显著高于其他生境,草原化草甸长蝽科密度显著高于草甸(图5)。6月草原化草甸甲螨亚目的密度显著高于草甸和沙化草甸,10 月草原化草甸和沼泽化草甸甲螨亚目的密度显著高于草甸和沙化草甸;6 月草原化草甸辐螨亚目的密度显著高于沼泽化草甸,10 月草原化草甸辐螨亚目的密度显著高于草甸和沙化草甸(图5)。4 种高寒草甸长角跳科和等节跳科密度变化趋势相近,6 月和10 月草原化草甸长角跳科和等节跳科密度显著高于其他草甸生境,6 月沼泽化草甸等节跳科密度又显著高于草甸和沙化草甸,10 月沼泽化草甸长角跳科密度又显著高于草甸和沙化草甸(图5)。4 种高寒草甸球角跳科密度变化与长角跳科和等节跳科不同,6 月沼泽化草甸和沙化草甸球角跳科密度显著高于草甸和沼泽化草甸(图5)。
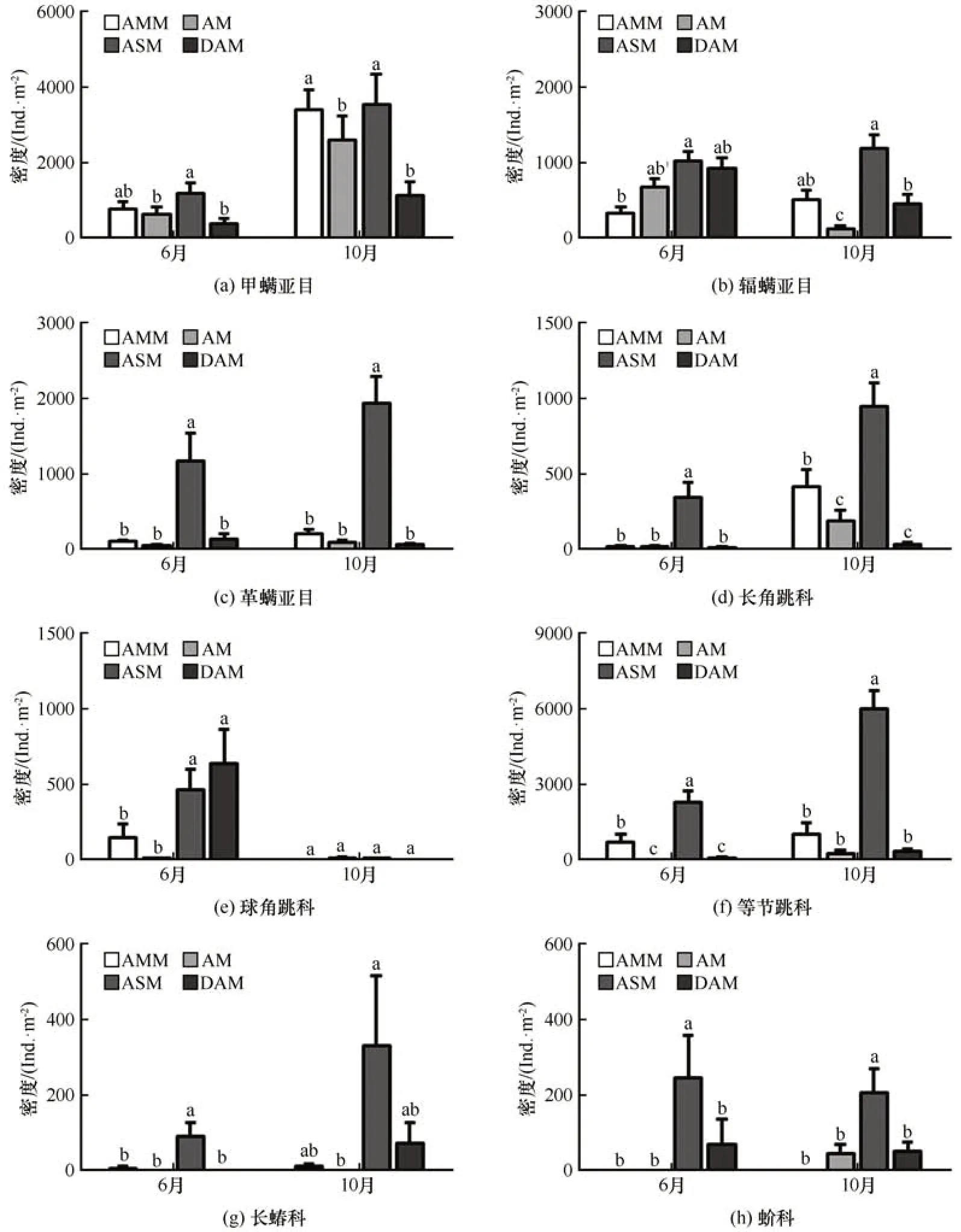
图5 6月和10月疏勒河源区沼泽化草甸(AMM)、草甸(AM)、草原化草甸(ASM)和沙化草甸(DAM)8个主要中型土壤动物密度比较Fig. 5 The density of eight dominant soil mesofauna in the alpine marsh meadow( AMM), alpine meadow( AM), alpine steppe meadow( ASM), and desertification alpine meadow( DAM) in June and October in the Shule River headwaters:Oribatida( a), Prostigmata( b), Mesostigmata( c), Entomobryidae( d), Hypogastruridae( e),Isotomidae( f), Lysaeidae( g), Coccoidea( h)
2.4 高寒草甸土壤水分变化对中型土壤动物分布的影响
高寒草甸土壤水分有效性变化不仅影响高寒草甸初级生产力,它还会直接或间接影响中型土壤动物的分布及多样性。高寒草甸表层土壤水分与中型土壤动物密度、类群丰富度和多样性指数均呈显著的二次曲线关系,它们随着高寒草甸土壤水分含量的增加先升后降(图6)。高寒草甸土壤水分含量接近35.7%和33.2%时,中型土壤动物密度和类群丰富度达到最大值分别为每平方米6 424.2 头和每平方米7.6 科或亚目。中型土壤动物QBS-ar 指数与土壤水分含量也呈显著的二次曲线关系,高寒草甸土壤水分含量接近35.7%时,中型土壤动物QBS-ar指数达到最大为42.7(图7)。

图6 疏勒河源区土壤水分与中型土壤动物密度(a)、类群数(b)和多样性指数(c)的关系Fig. 6 Relationships of the density (a), groups richness (b), and Shannon-Wiener index (c) of soil mesofauna community with soil water content in the Shule River headwaters

图7 疏勒河源区土壤水分与中型土壤动物QBS-ar指数的关系Fig. 7 Relationship of the QBS-ar index of soil mesofauna community with soil water content in the Shule River headwaters
3 讨论
疏勒河源区地处青藏高原的东北缘、祁连山的西段,降水稀少,中型土壤动物物种数(科)低于青藏高原东南缘[29],略高于祁连山高寒草地[30]。疏勒河源区高寒草甸中型土壤动物群落中甲螨亚目是优势类群这与青藏高原其他区域的研究结果一致,跳虫中的等节跳科是优势的中型土壤动物类群,这与孙彩彩等[30]的研究结果相近,而与周磊等[29]的在青藏高原东南缘的研究结果不同。疏勒河源区沼泽化草甸、草甸、草原化草甸和沙化草甸与放牧引起冻土退化驱动土壤及地下水位变化有关[1,20],4 种草甸生境构成的高寒草甸退化梯度强烈影响中型土壤动物的分布及多样性,这与本研究前期关于地表节肢动物多样性对围封响应研究结果相近[20]。高寒草甸中型土壤动物密度和类群丰富度存在明显的季节变异,它在秋季的密度增大与祁连山南坡前指数期调查结果相近[25]。高寒草甸退化过程中,6 月和10 月中型土壤动物的密度、类群数和多样性指数的变化趋势相近,草原化草甸中型土壤动物的密度及多样性指数与螨类和跳虫等主要类群的密度均显著高于其他生境,这与吴鹏飞等[31]、武崎等[14]和周磊等[29]关于中小型土壤动物对高寒草甸退化的响应研究结果一致。疏勒河源区草原化草甸中型土壤动物数量及多样性最高,草原化草甸生境螨类和跳虫的密度也显著高于其他生境,这说明该生境土壤水分、植被和土壤环境适于螨类和跳虫等中小型土壤动物生存和繁殖[15,29,32]。沼泽化草甸和草甸植被盖度较高,地下水位浅导致土壤水分含量高、土壤温度低,这不利于节肢动物的生存和繁殖;草原化草甸和沙化草甸植被和土壤水分含量降低、土壤温度高,地表和土栖节肢动物的数量和多样性指数明显增高[1,20]。此外,研究还发现疏勒河源区降水主要集中在夏季,秋季降水稀少,这也促进了一些土壤螨类和跳虫的繁殖,故秋季中型土壤动物的密度明显增加。疏勒河源区4种高寒草甸生境甲螨亚目的密度在沙化草甸生境的密度大幅下降;辐螨亚目的密度则在草原化草甸最高,而革螨亚目密度仅在草原化草甸大幅增加,这与周磊等[29]关于螨类研究结果略有不同,不同螨类生物学特征不同导致它们对高寒草甸退化的响应模式不同。疏勒河源区10 月甲螨亚目的密度明显高于6 月,6月和10 月沙化草甸螨类的密度低于其他高寒草甸生境,土壤水分变化对甲螨亚目的影响较大;6 月和10 月辐螨亚目和革螨亚目在草原化草甸的密度最大,这些螨类数量变化可能与其他动物数量的增加有关。跳虫群落中2个优势类群长角跳科和节跳科的密度在4种高寒草甸生境变化均在草原化草甸生境大幅增加,这与周磊等[29]关于跳虫密度变化的研究结果相近。此外,研究还发现长蝽科和蚧科两种植食性类群主要分布在草原化草甸生境,这可能与土壤水分及植被群落有关。青海省海北州海晏县西海镇一项牲畜种类对土壤节肢动物多样性影响的控制试验也发现,藏羊单独放牧条件下土壤节肢动物丰富度指数、多样性指数和类群数较高;放牧与不放牧均降低了土壤节肢动物优势类群螨总科的组成比例,增加了甲螨总科的组成比例[30]。高寒草甸土壤动物对生境退化的响应与中低海拔区草地和灌丛草地不同,这可能与土壤水分及温度的限制有关。针对中国北方草地和中亚的研究均表明,土壤动物多样性及其功能随着草地或灌木林地退化而减少[32-33]。总之,中等退化程度的高寒草甸中型土壤动物的密度及多样性均较高,与植被和土壤养分变化相近,说明适度放牧可以提高中型土壤动物的多样性,还会改变中型土壤动物的群落结构及功能。
螨类和跳虫等主要中型土壤动物对高寒草甸退化过程生物和非生物环境要素的响应不同,它影响了高寒草甸中小型土壤动物食物网结构及其功能变化。一项在青藏高原模拟放牧强度的控制试验结果表明,中度放牧条件增加了线虫的数量和微小食物网结构的复杂性;根系的数量和质量改变了植食性和真菌食性线虫的数量,说明资源的上行效应强烈影响了低营养级的线虫[18]。高寒草甸线虫、螨类和跳虫及大型土壤动物均随着高寒草甸的退化而增加,这与草甸初级生产力和多样性变化一致,说明高寒草甸生态系统适度放牧有利于维持地上和地下生态系统多样性及其生态功能[4,11,20,31]。本研究中还发现,高寒草甸中型土壤动物QBS-ar指数变化与中型土壤动物多样性指数变化相近,草原化草甸生境中型土壤动物QBS-ar 指数显著高于其他生境,这也表明草原化草甸生境土壤质量好于其他高寒草甸生境[1]。中型土壤动物QBS-ar 指数可以指示土壤质量变化,它在平原和山区土地利用变化研究中已经得到了较好地应用,准确反映土壤健康变化[27,34]。草原化草甸螨类和跳虫的A/C 比值显著低于沼泽化草甸、草甸和沙化草甸生境,高寒草甸A/C 值较高与尹文英[20]的研究结果相近,草原化草甸土壤水分降低提高了螨类和跳虫的密度,但螨类的密度增加快于跳虫,这也导致该生境A/C 值显著降低。青藏高原高寒草甸生态系统草原化草甸大型和中型土壤节肢动物多样性均高于沼泽化草甸,沙化草甸大型和中小型土壤动物多样性均降低,土壤环境变化影响大型和中型土壤动物的分布及多样性[29,32,35-36]。高寒草甸适度放牧有利于维持大型和中小型土壤动物多样性及其功能,这与土壤微生物和植被对围封及放牧的响应研究结果一致,适度放牧有利于维持高寒草甸土壤生态系统的健康与稳定[13-15]。
土壤水分是高寒草甸初级生产力的重要限制因子,高寒草甸退化土壤水分有效性也随之降低,它决定土壤动物多样性及关键种对放牧强度的响应模式[12,16]。青藏高原线虫和土壤节肢动物与降水、土壤有机碳含量和草本物种丰富度呈正相关,而与pH 和土壤温度的季节变化呈负相关;年平均温度和降水的季节变化对线虫和土壤节肢动物的影响较小[14]。一项关于高寒草甸地下水位控制的实验发现,不同小型土壤动物类群对实验处理的响应不同,螨类对地下水位下降处理的响应不敏感,跳虫和线虫数量对地下水位下降处理存在正响应,这也表明高寒草甸跳虫和线虫数量随着表层土壤水分降低而增加[18]。高寒草甸土壤水分含量较高会抑制土壤动物生存和繁殖,降低地下水位或适度放牧降低表层土壤水分都会促进土壤动物活动,而随着高寒草甸退化程度的加剧,土壤水分的匮缺又会抑制土壤动物的活动。王宇彤等[35]对青藏高原高寒草甸不同微生境(沟底平地、阴坡、阳坡和山顶)线虫分布的研究发现,阳坡线虫的丰度和多样性最高,这也表明高寒草甸过高和过低的土壤水分抑制了线虫等土壤动物的活动。Kooch等[32]在干旱区山地研究发现,生境退化过程中土壤水分、温度和肥力影响土壤生物活动,螨类和跳虫数量变化与蚯蚓和线虫及真菌相近,均随着退化程度的加剧而下降。笔者研究发现,疏勒河源区高寒草甸退化过程中,中型土壤动物密度、类群丰富度和多样性指数均与土壤水分含量呈显著的二次曲线关系,它们随着土壤水分含量的增加先增后降。青藏高原土壤动物多样性对高寒草甸退化的响应研究均发现,降水和土壤水分变化是影响大型和中小型土壤动物数量及多样性的主要环境要素[12,18,29,37],高寒草甸退化过程中适宜的土壤水分含量有利于维持土壤动物多样性及其功能。本研究还发现,疏勒河源区高寒草甸中小型土壤动物QBS-ar 指数与土壤水分变化也呈显著的二次曲线关系,这表明土壤质量变化也受土壤水分含量变化的影响。
4 结论
疏勒河源区高寒草甸退化强烈影响螨类和跳虫等中型土壤动物密度及多样性变化,高寒草甸退化过程中草原化草甸(即中等退化高寒草甸)具有最高的中型土壤动物密度和多样性。中型土壤动物群落对高寒草甸退化的响应存在一定的季节变异,6月和10月中型土壤动物密度、类群丰富度和多样性指数均显著高于其他高寒草甸生境,而10月沼泽化草甸中型土壤动物密度和类群丰富度均显著高于草甸和沙化草甸。中型土壤动物QBS-ar 指数变化与密度、类群丰富度和多样性变化规律相近,A/C 值则与中型土壤动物群落和QBS-ar 指数变化相反。螨类和跳虫对高寒草甸退化的响应模式不同并存在季节变异,6 月螨类的密度在草原化草甸显著高于其他草甸生境,10 月草原化草甸和沼泽化草甸螨类的密度显著高于草甸和沙化草甸;6 月和10 月跳虫的密度在草原化草甸显著高于其他草甸生境。进一步研究发现,螨类中革螨亚目、甲螨亚目和辐螨亚目密度对高寒草甸退化的响应模式不同并存在季节变异,跳虫中长角跳科和等节跳科密度对高寒草甸退化的响应模式相近。高寒草甸退化过程中土壤水分含量与中型土壤动物密度、类群丰富度、多样性指数和中型土壤动物QBS-ar指数均呈显著的二次曲线关系,中型土壤动物群落指数均随着高寒草甸土壤水分含量的增加先增后降。总之,高寒草甸退化过程中土壤水分变化强烈影响中型土壤动物群落组成及多样性,适宜的土壤水分含量有利于维持高寒草甸中型土壤动物多样性及其功能。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
江苏农业科学(2019年5期)2019-09-02 14:01:46
活力(2019年21期)2019-04-01 12:17:10
广东农业科学(2017年5期)2017-08-29 10:37:31
绿色中国(2017年7期)2017-08-02 17:47:11
环境保护与循环经济(2017年11期)2017-03-16 03:17:07
绿色中国(2016年1期)2016-06-05 09:03:00
绿色中国(2016年1期)2016-06-05 09:03:00
