
日粮中木聚糖、低聚木糖和木聚糖酶对单胃动物肠道健康和生长性能的影响(续2)
2024-01-12 00:33:24张相鑫译自202111609
国外畜牧学(猪与禽) 2023年6期
张相鑫 译自,2021,11(3):609
靳文广 校 王晶晶 审 孟祥光 制表
5 木聚糖酶
除了对木质纤维素材料进行化学和酶处理来生产用于动物饲养的低聚木糖外,在单胃动物饲料中添加木聚糖酶也引起了研究人员极大的兴趣。正如上文所述,常见谷物及相关副产品含有不同水平的非淀粉多糖(non-starch polysaccharide,NSP),这会增加动物小肠中食糜的黏度,导致营养物质的消化率和吸收率降低。此外,食糜黏度增加会导致黏膜表面的形态发生变化,增加小肠中病原体载量,延长食糜的转运时间。
木聚糖酶被归类为糖苷水解酶,可催化水解木聚糖聚合物中的1,4-β-D-木糖苷键。内切木聚糖酶的特点是水解木聚糖聚合物内部的键,外切木聚糖酶的作用是从还原端或非还原端水解木聚糖。最初,Wong 等根据木聚糖酶的理化性质尝试对其进行分类,并提出分为两类,即低分子量(<30 kDa) 和碱性等电点(isoelectric point,pI) 的木聚糖酶,以及高分子量(>30 kDa)和酸性pI 的木聚糖酶。这种分类方法很快被一个更完整的系统所代替,因为在鉴定的木聚糖酶(特别是真菌木聚糖酶)中,约有30%无法按该方法进行分类。新的分类系统不仅促进了木聚糖酶的分类,而且促进了糖苷水解酶的分类(EC 3.2.1.x),至今仍是标准的分类方法。该系统仅根据催化结构域的一级结构对酶进行分类,然后将相关序列的酶分为一个家族。最初,木聚糖酶和纤维素酶被分为6 个家族;目前,在碳水化合物活性酶数据库(截至2020 年12 月22 日)中已确认有168 个糖苷水解酶(glycoside hydrolase,GH) 家族。由于趋异进化,一些家族具有相关的三维结构,被分为更高的级别,称为族。到目前,已经提出了18 个不同的族(GH-A 至GH-R),除了GH-A 包含24 个家族外,大多数族包含2~3 个家族。木聚糖酶通常分布在GH10 和GH11。
到目前为止,GH5、GH8、GH10、GH11、GH30、GH43、GH51 和GH98 是EC 3.2.1.8 归类的8 个糖苷水解酶家族。不同内切木聚糖酶的分类反映了它们在各种应用中的用途,其中GH10 和GH11 是最常用和研究最多的。GH10和GH11 构成了试验中用于生产不同类型低聚木糖的主要酶。从细菌内切-β-木聚糖酶中获得的基因主要来自厚壁菌门和放线菌门的革兰阳性菌,其中最常见的是杆菌纲和梭菌纲。链霉菌属是大多数产内切-β-木聚糖酶的候选菌。已经发现,获得低聚木糖的模式至少在某种程度上依赖于GH 家族。对木霉菌木聚糖酶进行的一项研究显示,GH10 木聚糖酶对小分子底物具有较高的活性,导致优先产生聚合度较低的低聚木糖(木二糖到木戊糖)。相反,GH11 木聚糖酶对大分子底物更具活性,导致产生聚合度较高的低聚木糖(木二糖到木己糖)。此外,Abou-Hachem 发现,湖北拟酵母菌产生的两种木聚糖酶分别为GH10 和GH11,不能产生木糖或木二糖,而只产生木二糖到木庚糖。这一发现表明,低聚木糖的产生模式是酶依赖性,而不仅仅是家族依赖性。GH10 木聚糖酶已被证明能水解异木聚糖中的β-1,4-木糖苷键,当在活性位点时也能在一定程度上水解葡萄糖。据报道,GH10 木聚糖酶作用于不溶性木聚糖的能力较低,但可作用于取代木糖主链的木聚糖。有趣的是,GH10 木聚糖酶对短链低聚木糖表现出很高的亲和力,表明存在一个小的底物结合位点。
GH11 木聚糖酶对有木糖主链且不存在葡萄糖的异木聚糖表现出较高的活性。有趣的是,GH11 木聚糖酶更喜欢长链底物,对聚合度3(木三糖)到聚合度5(木戊糖)的低聚木糖的亲和力较高,而对聚合度小于3 的低聚木糖没有活性。与GH10 木聚糖酶相比,GH11 木聚糖酶的取代限制通常导致取代的木聚糖的总降解量更低,而GH10 木聚糖酶可以在活性位点降解取代度高的木聚糖。然而,由于GH11木聚糖酶的分子量较小,与GH10 木聚糖酶相比,该酶降解不溶性木聚糖的效率更高。
之前被归类到GH5 的木聚糖酶现在被重新归类到GH30,因为系统发育分析表明这些酶与GH30 更相似。GH30 木聚糖酶分为8 个亚家族,主要基于构成β-结构的二级结构的排列。大多数活性较高的木聚糖酶属于第8 亚家族,第7 亚家族中的两种木聚糖酶除外。目前,有9 种GH30 木聚糖酶的结构已经确定,其中4 种已被广泛研究,分别是枯草芽孢杆菌的BSXynC、菊基腐病菌(Erwinia chrysanthemi) 的EcXynA、巴塞罗那类芽孢杆菌(Paenibacillusbarcinonensis)的Xyn30D 和溶纸梭菌(Clostridium papyrosolvens) 的CpXyn30A。BsXynC 和EcXynA 是第一批被归类为GH30 的木聚糖酶,具有GH30_8 木聚糖酶的共同特征。这两种木聚糖酶与GH10 和GH11 木聚糖酶不同,它们对葡萄糖醛酸木聚糖和被甲基葡萄糖醛酸或葡萄糖醛酸取代的低聚木糖具有较高的亲和力,而对未取代的木聚糖、低聚木糖和阿拉伯木聚糖的亲和力较低。来自巴塞罗那类芽孢杆菌的Xyn30D 归类于EC 3.2.1.136,仅对葡萄糖醛酸木聚糖有活性,而来自溶纸梭菌的CpXyn30A 归类于EC 3.2.1.8,对葡萄糖醛酸木聚糖的亲和力一般,但对阿拉伯木聚糖和低聚木糖的亲和力较高。
β-D-木糖苷酶(EC 3.2.1.37) 从非还原端把低聚木糖切割为木糖单体;乙酰木聚糖酯酶(EC 3.1.1.72)、阿魏酸酯酶(EC 3.1.1.73)、α-L-阿拉伯呋喃糖苷酶(EC 3.2.1.55.)、对香豆酸酯酶(EC 3.1.1.B10)和α-D-葡萄糖醛酸酶(EC 3.2.1.139) 催化侧基的去除。因此,从理论上来说,在玉米型饲料(富含纤维) 中添加木聚糖酶,可以增加饲料中纤维提供的能量,从而提高生长性能。然而,很少有研究报道消化率和生产性能都得到了改善。Bedford 提出的一个假设认为,这些非淀粉多糖酶(non-starch polysaccharide enzymes,NSPases) 可能产生低聚糖,然后向微生物区系发出信号,从而提高整个系统降解纤维的能力,而不是产生更多的可发酵糖。根据该假设,最近在鸡上进行的一项研究证明了这一点,与对照组相比,日粮中添加木聚糖酶35 d 后,木聚糖酶组鸡盲肠中的戊糖和阿拉伯低聚木糖的发酵程度更高。在Duarte 等进行的一项研究中,在断奶仔猪的两个饲养阶段的日粮中添加木聚糖酶或蛋白酶或两种酶的组合,共24 d。该研究发现,当两种酶组合使用时,猪的体重增加,而单独添加时则没有增加。该试验得出的结论是,添加木聚糖酶(45 000 XU/kg)可改善断奶仔猪的生长性能和肠道形态,降低食糜黏度,减少肠道氧化应激;此外,由于绒毛长度的增加、隐窝深度和细胞增殖的降低,蛋白酶在维持肠道形态方面更有效。这些结果与Passos等的研究结果有一定的相关性,他们发现,当饲喂含30%干酒糟及其可溶物(distillers dried grains with solubles,DDGS) 的 饲料时,添加木聚糖酶可降低仔猪空肠食糜的黏度,从而提高营养物质利用率。该研究获得的数据还表明,随着日粮中木聚糖酶水平的提高,干物质、有机物、能量、中性洗涤纤维和粗灰分的消化率分别提高了9.2%、8.5%、9.3%、12.4%和10.7%。这些结果与Diebold 等早期进行的研究一致,当在育肥猪日粮中添加木聚糖酶后,有机物、粗蛋白、粗纤维、中性洗涤纤维和能量的回肠消化率均有所提高,但幅度较低。有趣的是,一项关于在生长猪(体重26 kg)饲料中添加木聚糖酶的研究发现,干物质、粗蛋白和能量的表观回肠消化率提高幅度较低(<2%),但效果显著。当然,成年猪的胃肠道更长、更发达,分解纤维素的能力更强,料肉比更低,食糜通过速度更慢。与保育猪或早期生长猪等幼龄猪相比,母猪等大龄和成年猪消化纤维成分的能力更强,因为纤维消化率和纤维降解能力随着体重的增加而提高。事实上,Jørgensen 等报道,与生长猪相比,母猪在小肠中能消化更大比例的NSP。该研究还发现,母猪消化不溶性NSP 的能力更强;然而,母猪和生长猪对可溶性NSP 的消化率没有差异。成年猪和体重较大的猪对纤维降解和消化能力的增强,可能会限制添加到饲料中木聚糖酶的效果。Kerkaert 等通过试验研究了木聚糖酶对1 944 头生长育肥猪(初始体重22.63 kg,终末体重133.64 kg)生长性能和胴体品质的影响,结果发现,添加木聚糖酶组的猪与对照组的猪相比,在平均日增重、日均采食量或饲料转化率方面均没有差异。值得注意的是,该研究在营养充足的玉米-大豆型饲料中分别添加了0、5、10、20、40、75 IU/kg 的木聚糖酶。虽然在生长性能或接受注射治疗的猪的百分比和总体死亡率方面没有差异,但添加中等水平的木聚糖酶时,猪的屠宰率确实增加了。
常见饲料中的木聚糖酶抑制剂是另一个需要克服的障碍。20 世纪90 年代末,研究人员首次在小麦中发现了木聚糖酶抑制剂。在首次发现之后,在大麦、黑麦和玉米中也发现了这些蛋白类木聚糖酶抑制剂。小麦含有三种结构不同的抑制剂,即小麦木聚糖酶抑制剂(T.aestivumxylanase inhibitor,TAXI)、甜蛋白样木聚糖酶抑制剂(thaumatin-like xylanase inhibitor,TLXI) 和木聚糖酶抑制 蛋 白(xylanase-inhibiting protein,XIP)。其他谷物中的木聚糖酶抑制剂与小麦中的XIP 和TAXI 结构相似,因此通常称为XIP样蛋白和TAXI 样蛋白。研究表明,这些木聚糖酶抑制剂可以特异性的影响GH10 和GH11 木聚糖酶,但有趣的是,它们对植物合成的内源性酶和上述家族以外的木聚糖酶没有影响。GH10 和GH11 家族中有一些特定的木聚糖酶对木聚糖酶抑制剂不敏感,但这些被认为是例外。
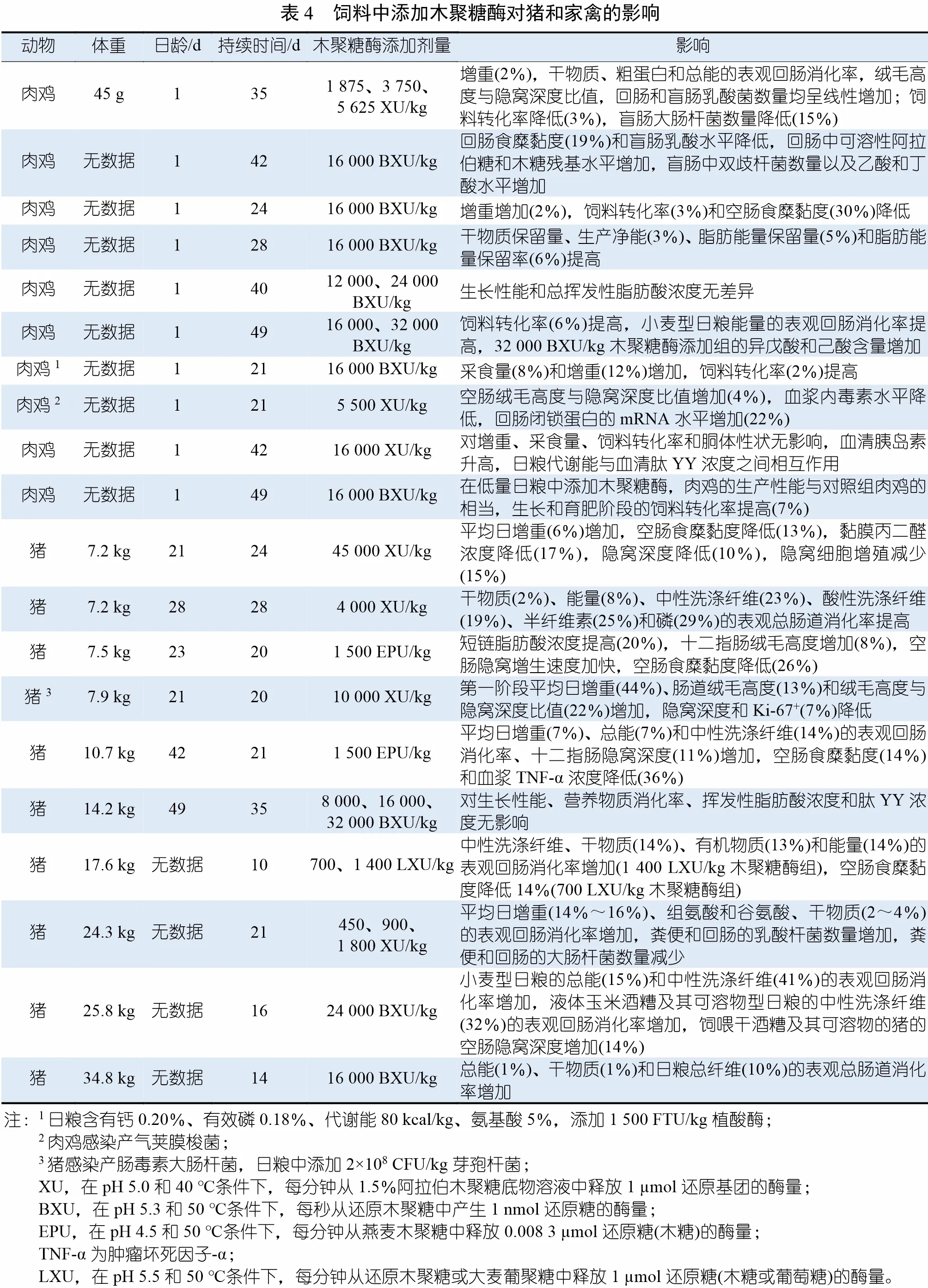
研 究 发 现,XIP-I 对GH10 和GH11 家 族中的真菌木聚糖酶有竞争性抑制作用,但对这些家族中的细菌木聚糖酶的抑制作用却不明显。还发现,XIP-I 可抑制GH13 家族中的α-淀粉酶,大麦α-淀粉酶/枯草杆菌蛋白酶抑制剂(barley α-amylase/subtilisin inhibitor,BASI) 可部分抑制GH11 家族中的木聚糖酶。值得注意的是,一些学者认为,谷物蛋白质对木聚糖酶和淀粉酶的这种交叉抑制作用可能是在植物进化过程中逐渐形成的,是一种抵御外来植物病原体攻击的保护功能。在大麦、玉米、水稻、燕麦和阿尔及利亚珍珠粟中发现了XIP 样抑制剂。TAXI 样蛋白通常抑制GH11 家族的真菌和细菌木聚糖酶,但不抑制GH10 家族的木聚糖酶。TAXI-I 样蛋白通常抑制具有低和高pI 值的木聚糖酶,TAXI-II 样蛋白仅抑制具有高pI 值的木聚糖酶。TLXI 是最新的蛋白类木聚糖酶抑制剂分类,于2006 年发现。同样,TLXI 蛋白倾向于抑制GH11 家族的木聚糖酶,但对GH10 家族的木聚糖酶没有抑制作用。除硬粒小麦外,有关其他谷物中存在TLXI 样抑制剂的数据有限。谷物中的TAXI、XIP 和TLXI 含量变化很大,主要取决于种类、品种、收获时间以及谷物的具体成分。例如,同一品种的小麦,在面粉中XIP-I 的含量为0.12~0.6 mg/g,在谷粒中为0.21~0.56 mg/g。在法国种植的20 种不同品种的小麦谷粒中,TAXI 的含量从0.05 mg/g到0.19 mg/g 不等。在猪和家禽饲料中添加木聚糖酶的效果见表4。
6 结论
总之,了解植物原料中木聚糖的结构差异、类型和浓度,以及各种可用的木聚糖酶,对于未来饲养动物和了解纤维在饲料中的作用至关重要。木聚糖底物的特性差异会直接影响聚合物向功能性更强的低聚糖的内源性和外源性裂解,因此其特性是产生理想效果的关键。添加低聚木糖似乎能对各种动物物种产生生理益处,并且与其他益生元相比,通常添加较低的剂量就能产生这些益处。现有的试验证明,低聚木糖可有助于预防或减轻胃肠道疾病,并增加有益菌群的数量。这些结果令人鼓舞;然而,与果寡糖、甘露寡糖和低聚半乳糖等其他益生元相比,关于低聚木糖的数据仍然有限,因此需要进一步的研究来巩固这些令人鼓舞的初步结果。在家禽和猪饲料中添加木聚糖酶可消除常见饲料原料中NSP 的抗营养作用,并为提高纤维的能量贡献和产生低聚木糖在远端肠道发酵提供机会。然而,由于动物年龄、饲料成分、添加持续时间和木聚糖酶抑制剂的存在等多种因素,添加木聚糖酶的效果差异很大。此外,即使报道了添加木聚糖酶的益处,但这些益处究竟是NSP 释放营养物质、降低食糜黏度还是产生短链低聚木糖的结果,仍然存在争论。与许多其他饲料添加剂一样,其效果以及作用方式可能是多因素的。与食糜黏度有关的问题主要与家禽饲养有关;然而,家禽业在遗传变异和日粮组成方面更加复杂。随着养猪业的发展变得更加可持续,为了养活不断增长的全球人口,越来越多来自不同行业的副产品可能会用于猪饲料,因此了解食糜黏度对动物饲养的影响十分重要。未来的研究重点是在家禽和猪的玉米型饲料中添加木聚糖酶,研究其在动物体内的作用模式,这将大大提高饲养结果的一致性,并加深我们对碳水化合物酶的整体了解。是在饲料中添加木聚糖酶来水解动物肠道内的木聚糖,还是在动物饲料中添加体外制备的低聚木糖以提高动物生长速度和肠道健康,目前仍存在争议,未来的研究应关注这一空白。在物种间和物种内比较低聚木糖产生的生理效应程度,不仅会受到所使用的低聚木糖产品纯度差异的影响,还会受到产品聚合度和取代类型的影响。
续完。
猜你喜欢
阅读(中年级)(2022年9期)2022-10-08 01:56:14
林产化学与工业(2022年4期)2022-09-13 09:47:12
动物营养学报(2022年7期)2022-08-09 09:38:52
红蜻蜓·低年级(2022年2期)2022-03-19 22:44:41
红蜻蜓·低年级(2021年12期)2021-12-19 15:06:23
湖南饲料(2019年5期)2019-10-15 08:59:10
动物营养学报(2018年12期)2018-12-13 06:01:48
中国酿造(2016年12期)2016-03-01 03:08:21
广东饲料(2016年8期)2016-02-27 11:10:01
中国医科大学学报(2015年10期)2015-03-01 02:09:50
