
黑水虻幼虫粉替代不同比例鱼粉对加州鲈生长、机体健康和肌肉品质的影响
2024-01-11 09:38游翠红李鸿伟董烨玮谭小红黄燕华
水生生物学报 2024年3期
游翠红 李鸿伟 周 萌 林 蠡 董烨玮 谭小红 黄燕华
(仲恺农业工程学院健康养殖创新研究院 & 动物科技学院, 广州 510225)
大口黑鲈(Micropterus salmoides)俗称加州鲈,隶属鲈形目(Perciformes)、太阳鱼科(Centrarchidae)、黑鲈属(Micropterus), 是中国从北美引进的一种暖水肉食性淡水鱼类。加州鲈生长速度快、肉质鲜美、肌间刺少, 深受消费者们的欢迎。加州鲈已经成为我国重要的淡水养殖鱼类。2021年, 我国加州鲈的养殖产量高达70.21万吨, 仅次于四大家鱼、鲫和罗非鱼的产量[1]。加州鲈对蛋白质的需要量在40%以上, 对脂肪的需要量也超过10%, 且其配合饲料中需要添加高比例的鱼粉(40%—50%)[2]。由于鱼粉价格昂贵, 且资源的获得不具有可持续性。近年来, 许多研究尝试用其他动植物蛋白源如鸡肉粉、血粉、豆粕、羽毛粉等替代加州鲈配合饲料中的部分鱼粉, 以降低养殖成本[2—4]。
黑水虻(Hermitia illucens)是双翅目水虻科昆虫, 繁殖速度快, 抗逆性强, 幼虫以畜禽粪便、餐厨垃圾、木质纤维素等为食物, 转化为高质量的昆虫蛋白[5]。黑水虻幼虫的蛋白质、脂肪含量很高, 分别占干物质的17.5%—63%和12.8%—49%, 其蛋白富含精氨酸、亮氨酸、赖氨酸、缬氨酸等必需氨基酸, 必需氨基酸组成与鱼粉相似; 其脂肪富含棕榈酸(C16∶0)和月桂酸(C12∶0)等饱和脂肪酸及油酸(C18∶1n-9)和亚油酸(C18∶2n-6)等单不饱和脂肪酸和多不饱和脂肪酸[5—7], 可被高效利用产能促进养殖动物生长。此外, 黑水虻幼虫还富含钾、钙、镁等矿物质及几丁质、抗菌肽等生物活性物质[6,8]。因此, 黑水虻幼虫是一种优质的昆虫蛋白源。已经在很多鱼类中开展了黑水虻虫粉部分替代鱼粉的研究, 如虹鳟(Oncorhynchus mykiss)[9,10]、大西洋鲑(Salmo salar)[11]、欧 洲 鲈(Dicentrarchus labrax)[12]、乌鳢(Channa argus)[13]、建鲤(Cyprinus carpiovar.Jian)[14]、加州鲈[15—18]等。但是, 已经报道的在加州鲈中开展的用黑水虻虫粉替代鱼粉的研究中, 基础饲料中鱼粉的含量很高, 一般都超过40%[15—18]。本研究拟在鱼粉含量较低(30%)的基础饲料中, 用黑水虻幼虫粉替代不同比例的鱼粉(16.67%、33.33%和50%), 研究对加州鲈幼鱼的生长性能、全鱼和组织的营养成分、机体健康、肌肉和肝脏的氨基酸、脂肪酸组成的影响, 为研制营养组成科学合理且成本低的加州鲈配合饲料提供资料。
1 材料与方法
1.1 实验饲料
根据加州鲈的营养需求, 配制 4 种等氮、等脂的实验饲料(45%蛋白, 15%脂肪), 其中, 基础饲料含30%鱼粉(FM30), 其他3组实验饲料分别用黑水虻幼虫粉替代16.67%鱼粉(FM25)、33.33%鱼粉(FM20)、50%鱼粉(FM15), 4种实验饲料的原料组成及实测的营养成分见表1, 实验饲料的氨基酸组成见表2。黑水虻幼虫购于广州飞禧特生物科技有限公司, 海藻多糖由青岛海兴源生物科技有限公司提供, 其他原料由湖州海皇生物科技有限公司提供。鱼粉、豆粕、烘干的黑水虻幼虫等饲料原料粉碎后, 过60目筛, 充分混合均匀后, 用SLX-80型双螺杆挤压机制成2.0 mm颗粒饲料, 60℃烘干后室温冷却, 用封口袋密封包装, 置于-20℃冰箱保存备用。
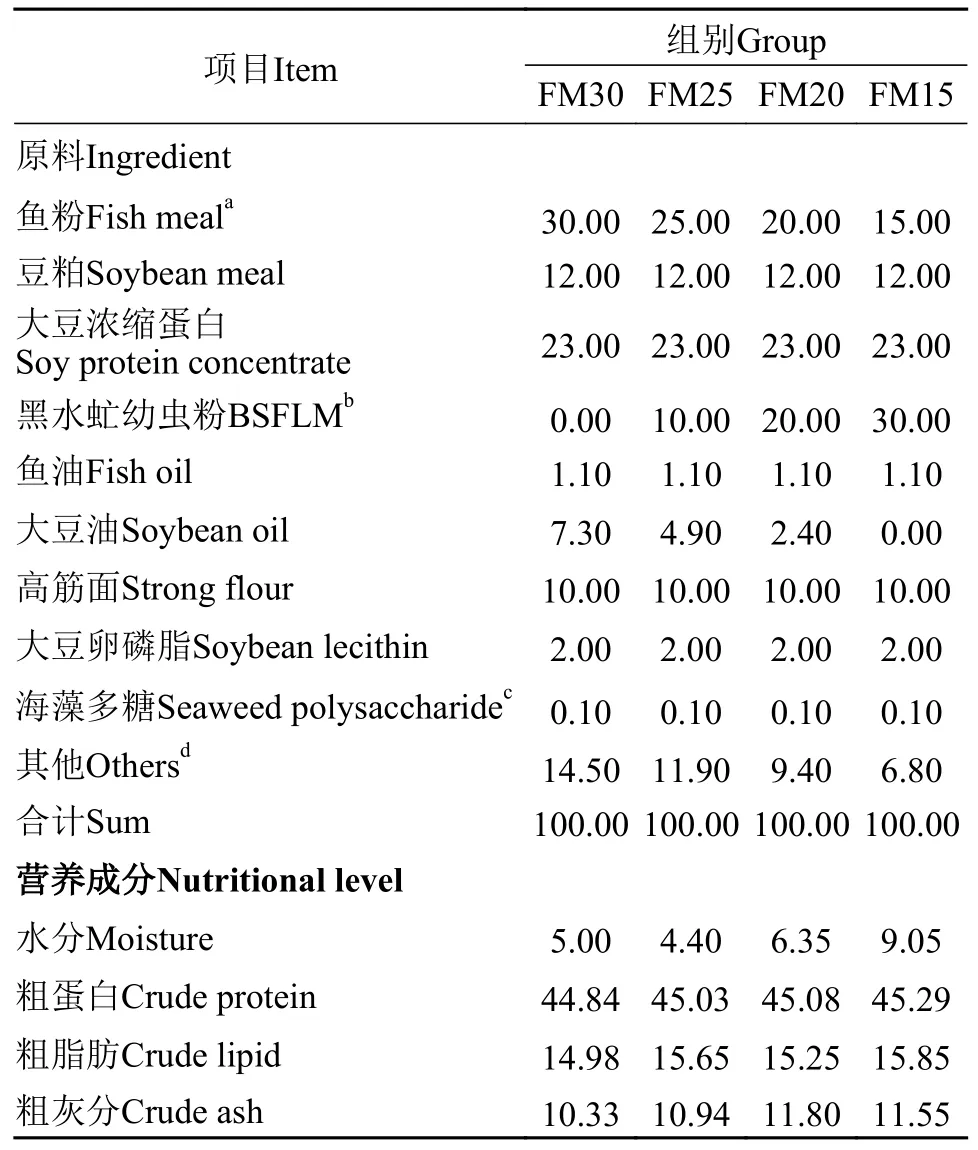
表1 实验原料的组成和营养成分(风干基础, %)Tab.1 Ingredients and proximate composition of the experimental diets (air-dry basis, %)
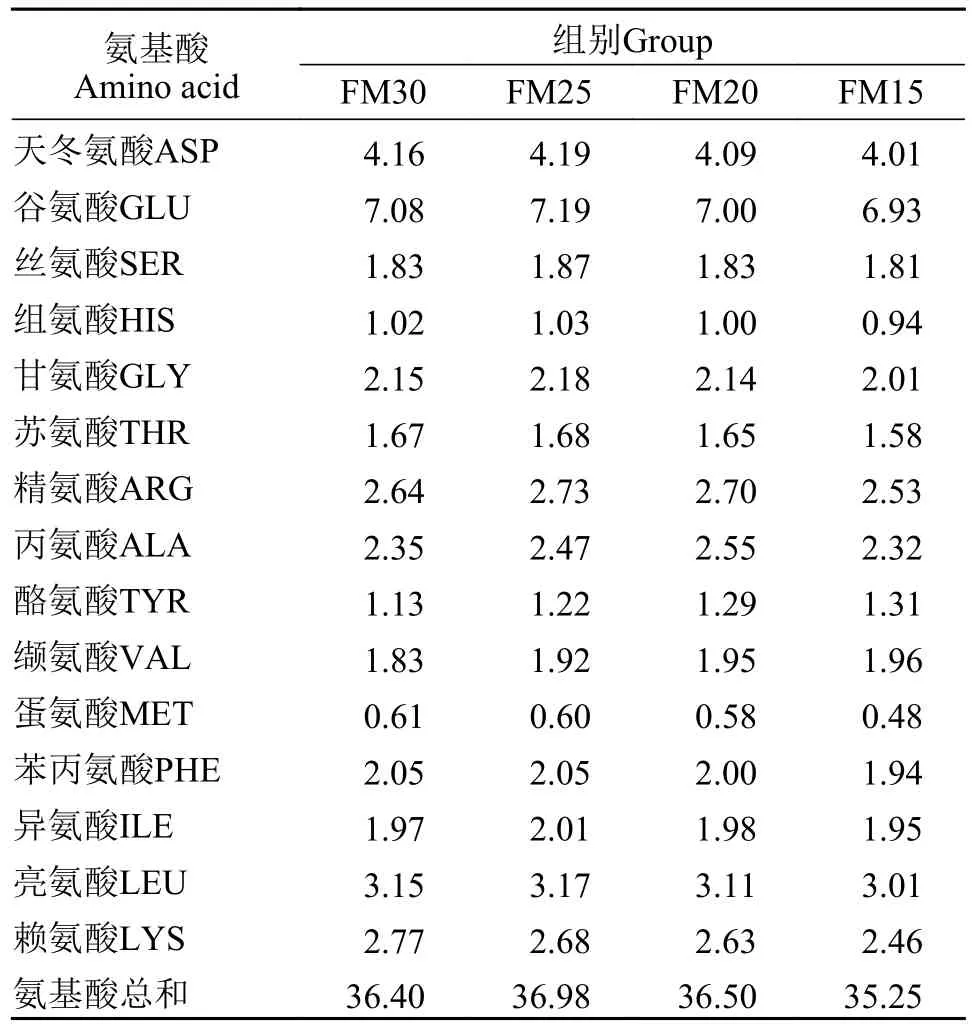
表2 实验饲料的氨基酸组成(%总氨基酸)Tab.2 Amino acid composition of the experimental diets (% total amino acids)
1.2 实验鱼及养殖实验
养殖实验在室内循环水养殖系统中进行, 养殖缸体积为300 L。放鱼前3天, 使用0.01%高锰酸钾溶液将养殖系统浸泡过夜进行消毒处理。实验鱼加州鲈幼鱼购于广东省佛山市三水白金水产种苗有限公司, 用水车运回后, 用4组实验饲料等比混合后的饲料在鱼塘网箱中驯化2周, 挑选体表健康、活力强的幼鱼(初始体质量约为17.05 g) 360尾, 随机分成4组, 置于室内循环水养殖系统的养殖缸中饲养, 每组3个重复缸, 每缸放鱼30尾鱼, 养殖8周。养殖期间, 每天饱食投喂2次(9:00和16:00), 每隔1—2d进行换水, 换水比例为30%—40%, 保持水温在25—27℃, pH在6.8—7.8, 溶解氧>6.0 mg/L。
1.3 样品采集
在养殖实验结束后, 实验鱼饥饿24h后进行采样。用100 mg/L间氨基苯甲酸乙酯甲磺酸盐(MS-222)将实验鱼进行麻醉, 准确测量每尾鱼的体质量和体长以计算生长指标。随后, 从每缸中随机取2尾鱼作为全鱼样品, 保存在-20℃冰箱中用于营养成分分析。另外, 从每缸中随机取6尾鱼, 使用一次性注射器从尾静脉处采血, 之后置于冰盘上, 无菌取内脏团, 分离肝脏、肠道组织, 并对内脏团、肝脏进行称重; 取肝脏、前肠、背肌样本用液氮速冻后置于-80℃冰箱保存, 用于酶活测定及营养成分分析; 另取1 cm长的前肠样本保存于4%多聚甲醛中, 用于组织切片分析。抽取的血液样品在4℃放置2h后, 3000 r/min离心10min, 吸取上层血清于2 mL离心管中, 置于-80℃中保存备用。
1.4 生长性能测定
测定增重率、特定生长率、饲料系数、蛋白质效率、肝体比、脏体比、肥满度、存活率等生长指标, 计算公式如下:
式中,Wt为终末鱼平均体质量(g),W0为初始鱼平均体质量(g),t为实验天数(d),Nt为终末鱼尾数,N0为初始鱼尾数,Ws为累计死鱼的体质量(g),Wf为摄入饲料量(g),P为饲料的蛋白质含量(%),Wh为鱼肝脏重(g),Wv为鱼内脏重(g),W为鱼体质量(g),L为鱼体长(cm)。
1.5 饲料、全鱼、组织样本的营养成分测定
水分含量测定采用烘箱105℃烘干至恒重的方法(GB/T6435- 2014), 粗蛋白含量测定采用凯氏定氮法(GB/T 6432-1994), 粗脂肪含量测定采用乙醚索氏抽提法(GB/T 6433-2006), 灰分含量测定采用马弗炉550℃灼烧至恒重的方法(GB/T 6438-2007)。
1.6 氨基酸组成和脂肪酸组成的测定
氨基酸测定参照赵红霞等[19]的方法, 肝脏和肌肉干物质样本经6 mol/L盐酸在110℃水解24h后, 采用安捷伦科技公司LC1260高效液相色谱仪(测试柱: 安捷伦ZORBAX c18, 150 mm × 5 μm)进行测定。脂肪酸测定参照Li等[20]的方法, 采用日本岛津公司GC-17A气相色谱仪, 通过脂肪酸标品(Sigma-Aldrich, 美国)鉴定脂肪酸, 采用面积归一法计算每种脂肪酸所占的百分比含量。
1.7 血清生化指标的测定
总胆固醇(Cholesterol, CHOL)、甘油三酯(Triglyceride, TG)、低密度脂蛋白(Low density lipoprotein cholesterin, LDL-C)、高密度脂蛋白(High density lipoprotein cholesterin, HDL-C)、血糖(Glucose,GLU)、谷 草 转 氨 酶(Aspartate aminotransferase,AST)、谷丙转氨酶(Alanine aminotransferase, ALT)、碱性磷酸酶(Alkaline phosphatase, ALP)、总蛋白(Total protein, TP)含量的测定交由广东省广州市海珠区新海医院采用日立7600全自动生化分析仪检测。
1.8 肠、肝的酶活力的测定
脂肪酶(Lipase; A054-2-1)、α-淀粉酶(Amylase;C016-1-1)和胰蛋白酶(Trypsin; A080-2-2)采用南京建成生物工程研究所试剂盒进行测定, 将肠道用0.86%的生理盐水清洗、剪碎, 按照组织∶生理盐水=1∶9(W/V), 冰浴匀浆后离心(4℃, 5000 r/min, 20min), 取上清液作为酶粗提液, 暂存于-20℃冰箱备用, 后续测定按照试剂盒说明书进行操作。超氧化物歧化酶(SOD; A001-1-2)、丙 二 醛(MDA; A003-1-2)含量、谷胱甘肽过氧化物酶(GSH-Px; A005-1-2)、过氧化氢酶(CAT; A007-2-1)、溶菌酶(LYS; A050-1-1)、碱 性 磷 酸 酶(AKP; A059-2-2)和 酸 性 磷 酸 酶(ACP; A060-2-1)也采用南京建成生物工程研究所试剂盒进行测定, 所有实验操作严格遵循说明书进行。
1.9 肠道组织学切片
用4%多聚甲醛固定的前肠样本, 通过酒精梯度脱水和石蜡包埋后切片, 切片厚度为 4 μm。将蜡片依次用苏木素-伊红 (HE) 染色, 中性树胶封片,用光学显微镜观察肠道切片并且拍照。
1.10 数据统计与分析
数据用平均值±标准误(mean±SE)表示, 使用SPSS 20.0 软件(IBM, armonk, USA)进行组间单因素方差分析(One-way ANOVA), 并进行 Duncan 氏多重比较分析, 显著性差异设为P<0.05。
2 结果
2.1 黑水虻幼虫粉替代鱼粉对加州鲈幼鱼生长性能的影响
如表3所示,各组鱼的成活率在97.67%—100%,无显著性差异(P>0.05)。各组鱼在终末体质量、增重率、特定生长率上也未呈现显著性差异(P>0.05)。在蛋白质效率上, FM20组显著高于其他组(P<0.05)。在饲料系数上, FM20组显著低于FM15和FM25组(P<0.05), 与对照组FM30没有显著性差异(P>0.05)。在肝体比和脏体比上, FM20和FM15组显著高于对照组(P<0.05)。上述结果显示, 在含30%鱼粉的基础饲料中, 用黑水虻幼虫粉替代0、16.67%、33.33%和50%鱼粉, 对加州鲈幼鱼的生长性能没有显著影响, 其中, FM20组(替代33.33%鱼粉)的生长性能较佳, 甚至优于对照组。
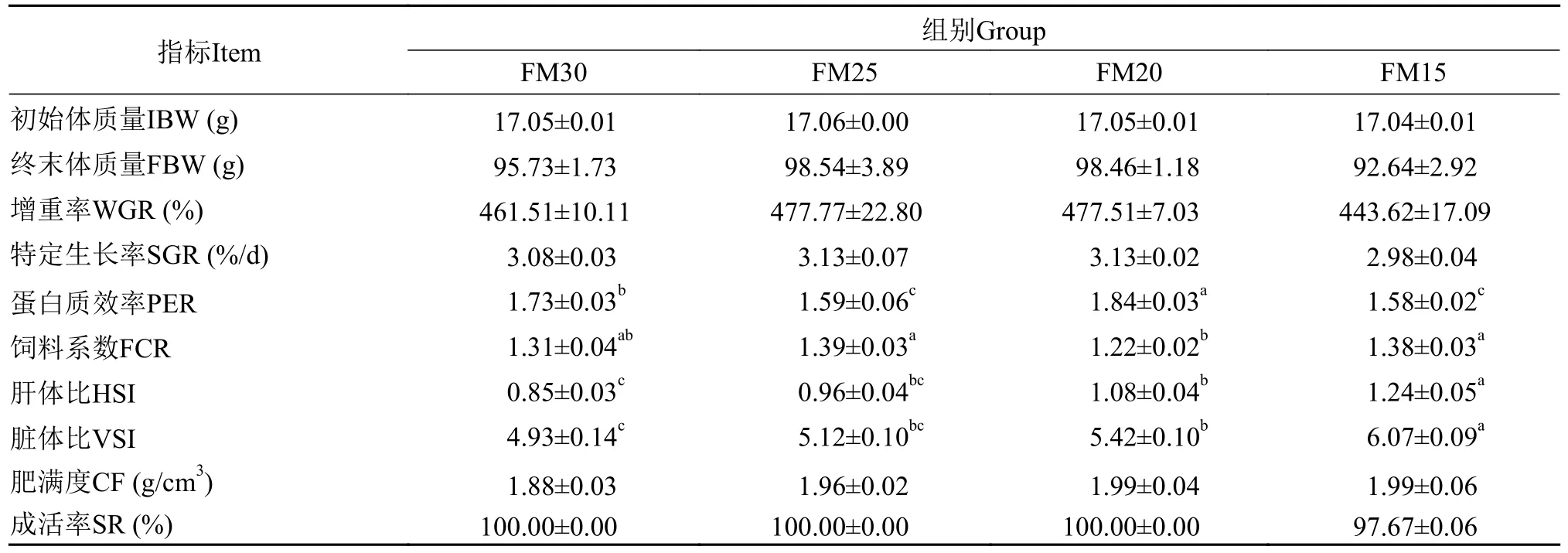
表3 养殖8周后加州鲈幼鱼的生长性能Tab.3 The growth performance of juvenile Micropterus salmoides fed the experimental diets for 8 weeks
2.2 黑水虻幼虫粉替代鱼粉对加州鲈全鱼、肌肉、肝脏的营养成分的影响
随着配合饲料中黑水虻幼虫粉替代鱼粉的比例升高, 全鱼、肝脏的粗脂肪含量呈现降低的趋势,FM15组鱼全鱼、肝脏的粗脂肪含量显著低于对照组(P<0.05); 全鱼的粗蛋白含量呈现先降低后升高再降低的趋势, 以FM20组全鱼的粗蛋白含量最高,显著高于对照组(P<0.05); 肌肉的粗脂肪含量呈现先升高后降低的趋势, 以FM20组肌肉的粗脂肪含量最高, 显著高于对照组(P<0.05); 肌肉的粗蛋白含量在组间没有显著性差异(P>0.05; 表4)。与养殖实验启动时的初始鱼相比, 养殖8周后, FM20组鱼肝脏和肌肉的粗脂肪含量明显增加, 以肝脏粗脂肪含量的增加幅度最为显著(表4)。
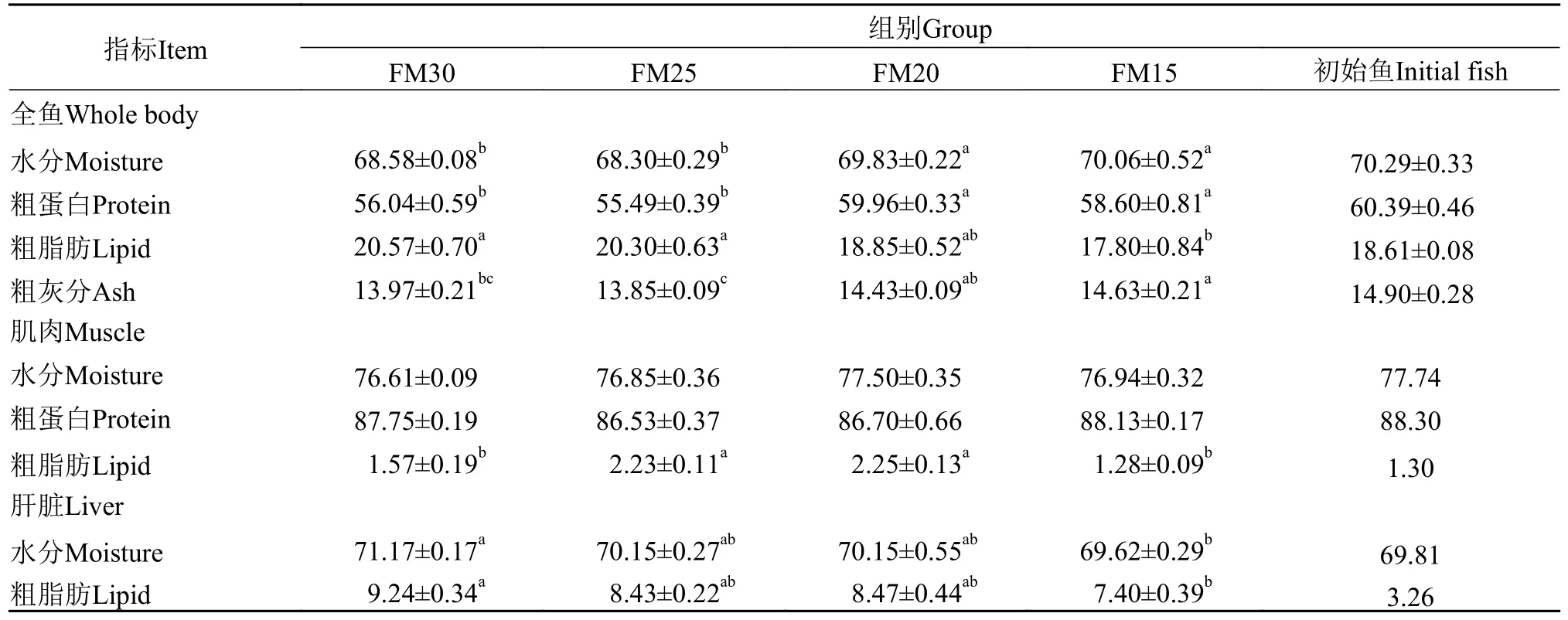
表4 养殖8周后加州鲈幼鱼的全鱼、肌肉、肝脏的营养成分(%干物质)Tab.4 The nutritional composition of the whole body, muscle and liver in juvenile Micropterus salmoides fed the experimental diets for 8 weeks (% dry matter)
2.3 黑水虻幼虫粉替代鱼粉对加州鲈血清生化指标的影响
如表5所示, 随着黑水虻幼虫粉替代鱼粉的比例升高, 血清谷丙转氨酶和谷草转氨酶活性、血糖含量呈现先降低后升高再降低的趋势; 与之相反,血清碱性磷酸酶活性及总胆固醇、高密度脂蛋白、低密度脂蛋白含量呈现先升高后降低再升高的趋势。添加黑水虻幼虫粉的3组鱼的血清甘油三酯的含量均显著低于对照组(P<0.05)。血清LDLC/HDL-C比率不受黑水虻幼虫粉替代鱼粉比例的影响(P>0.05)。与对照组相比, FM20组鱼在血清总胆固醇、谷丙转氨酶、谷草转氨酶、高密度脂蛋白、低密度脂蛋白含量上没有呈现显著性差异(P>0.05), 但是, 显著降低了血清碱性磷酸酶活性和甘油三酯含量(P<0.05), 增加了血清血糖含量(P<0.05)。
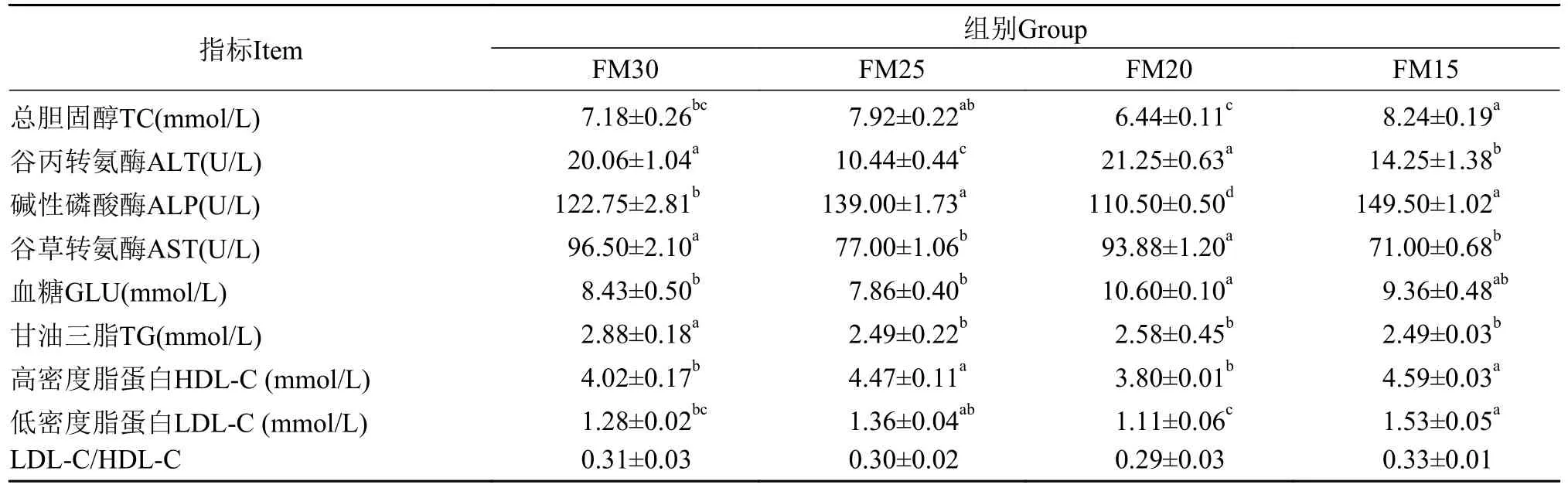
表5 养殖8周后加州鲈幼鱼的血清生化指标Tab.5 The serum biochemical index of juvenile Micropterus salmoides fed the experimental diets for 8 weeks
2.4 黑水虻幼虫粉替代鱼粉对加州鲈肝脏酶活的影响
如表6所示, 配合饲料中用黑水虻幼虫粉部分替代鱼粉, 提高了加州鲈幼鱼肝脏丙二醛含量, 降低了肝脏溶菌酶活性, 如FM20组与对照组相比, 显著增加了丙二醛含量, 降低了溶菌酶活性(P<0.05)。随着黑水虻幼虫粉替代鱼粉的比例升高, 肝脏超氧化物歧化酶、过氧化氢酶活性呈现先降低后升高趋势, 在FM15组达到最大值, 其与对照组相比差异不显著(P>0.05); 肝脏碱性磷酸酶、酸性磷酸酶活性呈现先升高后降低的趋势, 在FM25组达到最大值, 其与对照组相比差异不显著(P>0.05)。肝脏谷胱甘肽过氧化物酶活性在组间无显著性差异(P>0.05)。
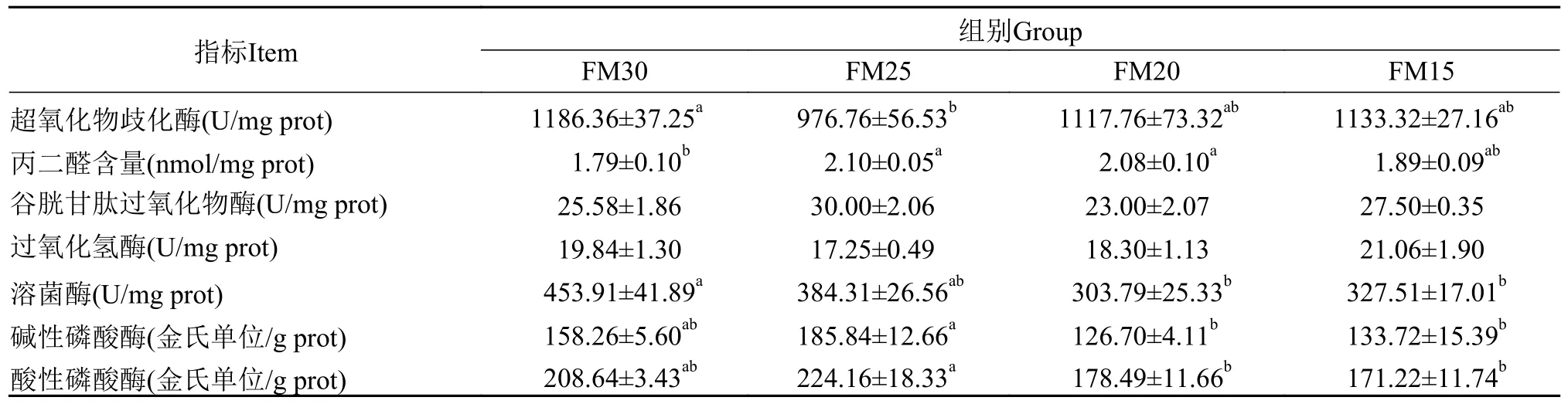
表6 养殖8周后加州鲈幼鱼的肝脏酶活Tab.6 The liver enzyme activity of juvenile Micropterus salmoides fed the experimental diets for 8 weeks
2.5 黑水虻幼虫粉替代鱼粉对加州鲈前肠酶活的影响
如表7所示, 在前肠组织中, 各组鱼的谷胱甘肽过氧化物酶、过氧化氢酶、溶菌酶活性未表现出显著性差异(P>0.05)。与对照组相比, FM20组鱼的前肠丙二醛含量显著升高(P<0.05), 碱性磷酸酶、酸性磷酸酶和α-淀粉酶的活性显著降低(P<0.05)。胰蛋白酶活性随黑水虻幼虫粉替代鱼粉比例的增加呈升高的趋势, 且FM15组鱼的胰蛋白酶活性显著高于对照组(P<0.05), FM20组鱼的脂肪酶活性显著高于FM25组(P<0.05)。

表7 养殖8周后加州鲈幼鱼的肠道酶活Tab.7 The intestinal enzyme activity of juvenile Micropterus salmoides fed the experimental diets for 8 weeks
2.6 黑水虻幼虫粉替代鱼粉对加州鲈肠道组织形态的影响
在配合饲料中用黑水虻幼虫粉部分替代鱼粉,加州鲈幼鱼前肠的上皮组织出现肠绒毛破损、肠皱褶粘连等现象, 以FM25和FM20组较为明显(图1)。对前肠组织结构形态学指标进行统计, FM25和FM20组鱼的肠壁厚度和肌层厚度显著低于对照组(P<0.05), FM25、FM20和FM15三组鱼的杯状细胞数量显著低于对照组(P<0.05); 同时, FM20和FM15组鱼的肠皱褶高度显著高于对照组(P<0.05; 表7)。随着黑水虻幼虫粉替代鱼粉的比例升高, 肠皱褶数量呈现升高的趋势, 但在各组间没有显著性差异(P>0.05; 表8)。
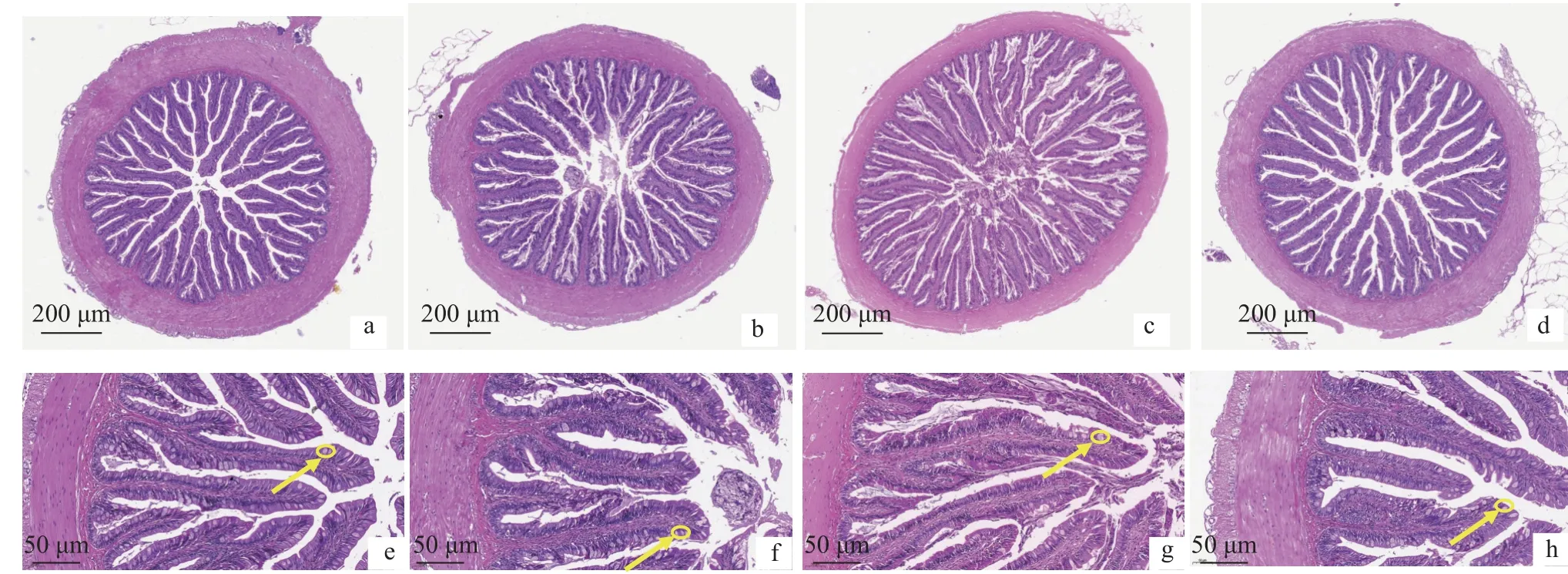
图1 养殖8周后加州鲈幼鱼的前肠组织形态Fig.1 Intestinal histology of the foregut in juvenile Micropterus salmoides fed the experimental diets for 8 weeks
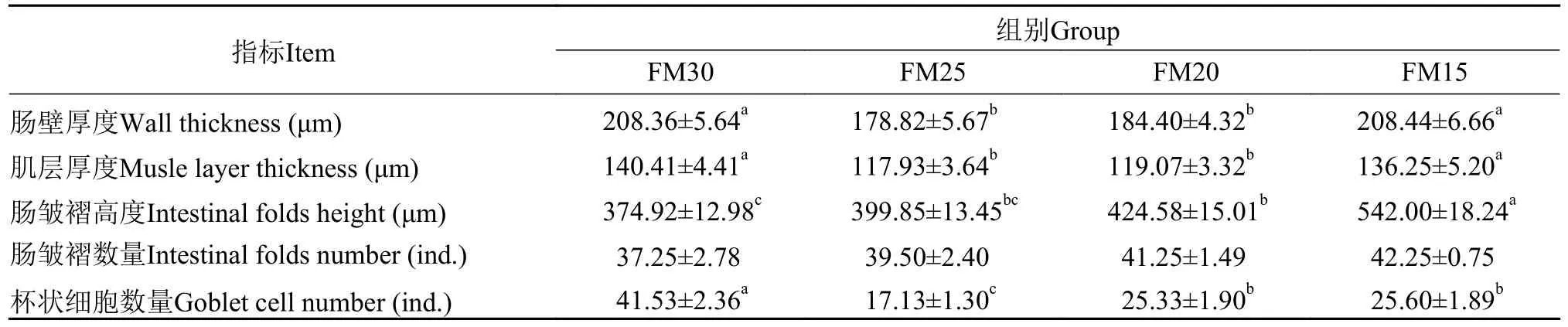
表8 养殖8周后加州鲈幼鱼的前肠组织形态学指标Tab.8 The intestinal morphological index of the foregut in juvenile Micropterus salmoides fed the experimental diets for 8 weeks
2.7 黑水虻幼虫粉替代鱼粉对加州鲈肝脏和肌肉的氨基酸组成的影响
如表9所示, 仅FM15组鱼的丙氨酸含量显著低于对照组(P<0.05), 其他氨基酸含量和总氨基酸含量与对照组相比差异不显著(P>0.05)。如表10所示, 仅FM20组鱼的总氨基酸含量显著低于对照组(P<0.05), 其他氨基酸含量在组间差异不显著(P>0.05)。

表9 养殖8周后加州鲈幼鱼的肝脏氨基酸组成(%总氨基酸)Tab.9 Amino acid composition in liver of juvenile Micropterus salmoides fed the experimental diets for 8 weeks (% total amino acids)
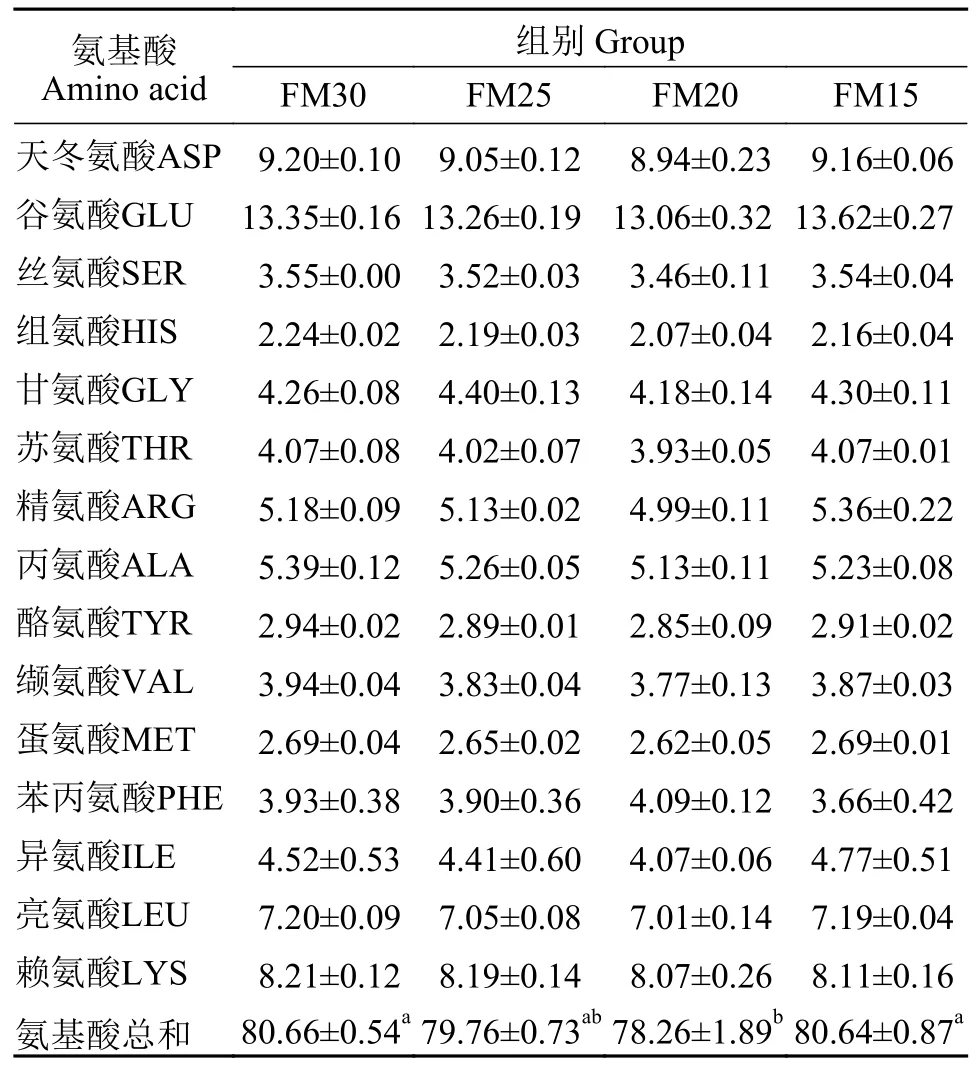
表10 养殖8周后加州鲈幼鱼的肌肉氨基酸组成(%总氨基酸)Tab.10 Amino acid composition in muscle of juvenile Micropterus salmoides fed the experimental diets for 8 weeks (% total amino acids)
2.8 黑水虻幼虫粉替代鱼粉对加州鲈肝脏和肌肉的脂肪酸组成的影响
如表11所示, 用黑水虻幼虫粉替代鱼粉, 降低了肝脏饱和脂肪酸C14∶0 和C16∶0的含量, 相反增加了C18∶0含量; 同时, 显著降低了单不饱和脂肪酸C16∶1n-7和C18∶1n-9的含量(P<0.05); 提高多不饱和 脂肪 酸C18∶2n-6、C20∶3n-3、C20∶5n-3 (Eicosapentaenoic, EPA)、C22∶5n-3 和C22∶6n-3 (Docosahexaenoic, DHA)的含量。值得注意的是, 3组黑水虻幼虫粉添加组肝脏的DHA、EPA和C22∶5n-3含量显著高于对照组(P<0.05), 以FM20组鱼的DHA、EPA和C22∶5n-3的含量最高; 同时, FM25和FM20组鱼的肝脏C20∶4n-6 (Arachidonic, ARA)含量也显著高于对照组(P<0.05)。但是, 3组黑水虻幼虫粉添加组肝脏的C18∶3n-3含量却显著低于对照组(P<0.05)。

表11 养殖8周后加州鲈幼鱼的肝脏脂肪酸组成(%总脂肪酸)Tab.11 Fatty acid composition in liver of juvenile Micropterus salmoides fed the experimental diets for 8 weeks (% total fatty acids)
如表12所示, FM15组鱼肌肉的C14∶0、C16∶0和FM20组鱼肌肉的C18∶0含量显著低于对照组(P<0.05)。用黑水虻幼虫粉替代鱼粉, 降低了肌肉单不饱和脂肪酸C18∶1n-9含量; 多不饱和脂肪酸C18∶2n-6含量随黑水虻幼虫粉添加水平的增加呈升高趋势, 且3组黑水虻幼虫粉添加组肌肉的C18∶2n-6含量均显著高于对照组(P<0.05), 但是, ARA含量却显著低于对照组(P<0.05)。随着黑水虻幼虫粉添加水平的升高, 肌肉C18∶3n-3含量呈现降低趋势,而EPA、DHA的含量呈现先降低后升高的趋势, 但FM15组鱼的肌肉EPA、DHA的含量依然显著低于对照组(P<0.05)。

表12 养殖8周后加州鲈幼鱼的肌肉脂肪酸组成(%总脂肪酸)Tab.12 Fatty acid composition in muscle of juvenile Micropterus salmoides fed the experimental diets for 8 weeks (% total fatty acids)
3 讨论
本研究结果表明, 在含30%鱼粉的基础饲料中,用黑水虻幼虫粉替代0、16.67%、33.33%、50%鱼粉, 对加州鲈幼鱼的生长性能和饲料利用效率无显著影响, 可使加州鲈幼鱼配合饲料中鱼粉的添加量最低可到15% (FM15); 当用黑水虻幼虫粉替代33.33%鱼粉, 即配合饲料中添加20%鱼粉(FM20)时, 加州鲈幼鱼的生长性能较高, 饲料系数最低, 甚至优于FM30对照组。在之前报道的对加州鲈幼鱼开展黑水虻幼虫粉替代鱼粉的研究中, 用黑水虻幼虫粉可替代5%—30%鱼粉, 使得加州鲈幼鱼配合饲料中鱼粉的添加量可降低到51.34%[15]或者31.5%[17], 均显著高于本研究得出的最低15%鱼粉添加量。有研究显示, 使用脱脂黑水虻幼虫粉, 可替代75%鱼粉,使得配合饲料中鱼粉的添加量可降低到10%, 而不影响加州鲈幼鱼的生长性能[18]。黑水虻幼虫的蛋白含量偏低, 脂肪含量偏高, 使用脱脂黑水虻可提高幼虫的蛋白含量, 因此可以提高配合饲料中鱼粉的替代比例[18]。在本研究中, 在黑水虻幼虫粉以适宜水平替代鱼粉的条件下, FM20组(配合饲料中添加20%鱼粉)加州鲈幼鱼(初始体重约为17.05 g)的特定生长率为3.13%/d, 与之前研究中生长性能最优的黑水虻幼虫粉替代组(配合饲料中添加51.34%鱼粉)加州鲈幼鱼(初始体重约为15.90 g)的特定生长率(3.26%/d)[15]持平, 但低于彭凯等[17]研究中生长性能最优组(配合饲料中添加34.87%鱼粉)加州鲈幼鱼(初始体重约为4.90 g)的特定生长率(4.06%/d)。前二者研究中加州鲈初始鱼体重比较接近(15.90—17.05 g), 远大于后者研究中初始鱼体重(4.90 g)。体重越小的鱼生长速度越快, 这也可能是后者的生长性能较高的原因之一。但是, 也有研究显示, 用黑水虻幼虫粉或预蛹黑水虻虫粉混合豆粕替代含20%鱼粉基础饲料中的66.67%鱼粉, 会导致加州鲈幼鱼的生长性能显著降低[16]。上述这些研究结果表明, 在加州鲈幼鱼的配合饲料中, 用黑水虻幼虫粉替代部分鱼粉(替代鱼粉的比例不超过50%), 不会影响加州鲈的生长性能甚至具有促进生长的作用, 这可能与黑水虻幼虫粉具有与鱼粉相似的氨基酸组成及富含几丁质、月桂酸、抗菌肽等生物活性物质有关[21,22]。几丁质能够刺激鱼类产生非特异性免疫, 并通过促进肠道乳酸菌等益生菌的增殖而抑制致病菌感染, 对革兰氏阴性菌具有抗菌作用;月桂酸具有抗炎作用, 能够有效抵抗革兰氏阳性菌;抗菌肽对多种菌具有抗菌作用, 有希望作为抗生素的替代品应用于水产养殖中[22]。在虹鳟[9]、大西洋鲑[11]、欧洲鲈[12]、乌鳢[13]、建鲤[14]、黄颡鱼(Pelteobagrus fulvidraco)[23]和 斑 马 鱼(Danio rerio)[24]等鱼类中开展的黑水虻幼虫粉部分替代鱼粉的研究, 也得到类似的不影响鱼类生长的结论。但是,黑水虻幼虫粉替代鱼粉的比例必须要控制在适宜范围之内, 过高会对鱼类的生长起负面影响。因为黑水虻外骨骼中的几丁质很难被鱼类消化, 且黑水虻富含饱和脂肪酸和n-6系列脂肪酸, n-6/n-3脂肪酸比例高, 机体脂肪酸组成不平衡, 不符合大多数鱼类对饲料脂肪酸的营养需求[21]。
碱性磷酸酶广泛分布于脊椎动物的各种组织中, 以肝脏、肾脏、骨骼和小肠中含量最为丰富。碱性磷酸酶可以使革兰氏阴性菌的膜蛋白脂多糖去磷酸化, 降低脂多糖与受体的结合能力, 从而降低脂多糖对动物机体的毒副作用[25]。碱性磷酸酶活性的降低与肠道炎症增加、生物失调、细菌易位及之后的全身炎症相关[26]。酸性磷酸酶是巨噬细胞溶酶体的标志性酶, 参与磷酸基团的转移和代谢, 可水解表面带有磷酸酯的异物以破坏异物[27]。溶菌酶可以水解革兰氏阳性菌的细胞壁多糖。碱性磷酸酶、酸性磷酸酶和溶菌酶在动物的非特异性免疫中发挥重要作用。丙二醛是动物机体脂质过氧化的产物。在本研究中, 虽然FM20组鱼呈现较佳的生长性能, 但其前肠和肝脏的丙二醛含量显著高于对照组, 前肠碱性磷酸酶、酸性磷酸酶活性和肝脏的溶菌酶活性均显著低于对照组, 血清碱性磷酸酶活性也显著低于对照组, 前肠上皮出现肠绒毛破损、肠皱褶粘连等结构受损情况, 肠道上皮的肠壁厚度、肌层厚度和杯状细胞数量都显著低于对照组, 显示FM20组加州鲈肝脏的非特异性免疫与抗氧化性能不及对照组, 其肠组织出现一定程度的损伤。彭凯等[28]也报道用黑水虻幼虫粉替代7.5%—30%鱼粉, 对加州鲈幼鱼的肠道造成损伤,且黑水虻幼虫粉替代比例越高, 肠道损伤越明显。用黑水虻幼虫粉或者预蛹黑水虻虫粉混合豆粕替代66.67%鱼粉, 加州鲈幼鱼肠绒毛的完整程度不及鱼粉组和鱼粉与豆粕混合组[16]。但是, 也有研究显示, 在大西洋鲑中用黑水虻幼虫粉或者脱脂黑水虻幼虫粉替代100%鱼粉, 不影响大西洋鲑的肠道健康[11,29]。用脱脂黑水虻幼虫粉替代16%—64%鱼粉, 对花鲈(Lateolabrax japonicus)的肠、肝组织形态都没有负面影响[30]。饲料中过量的几丁质对一些鱼类是有害的[16]。不同鱼类对几丁质的耐受程度不同, 可能与不同鱼类的胃腺和胰腺分泌几丁质酶的含量和活力有关[31]。本研究结果显示, 当用黑水虻幼虫粉替代33.3%鱼粉(FM20组)时, 加州鲈幼鱼生长良好, 能够耐受其造成的一定程度的肠、肝损伤, 可能源于黑水虻中的几丁质、月桂酸和抗菌肽等生物活性物质对加州鲈的有益作用大于负面作用。
本研究结果还显示, 用黑水虻幼虫粉替代鱼粉,具有降低加州鲈全鱼和肝脏脂肪的作用, 同时降低了血清甘油三酯的含量。肝脏是动物机体脂肪合成与分解代谢的中枢组织, 推测饲料中的黑水虻通过抑制肝脏合成甘油三酯, 从而造成全鱼和肝脏的脂肪含量及血清甘油三酯含量的降低, 表明其具有降脂作用。在黑水虻最适替代水平的FM20组, 全鱼和肝脏的粗脂肪含量低于对照组, 而肌肉的粗脂肪含量显著高于对照组, 表明用适宜水平的黑水虻幼虫粉替代鱼粉, 可促进肝脏等组织中的脂肪向肌肉组织转移, 有利于提高肌肉品质。类似的提高肌肉脂肪的结果也在虹鳟中有过报道[9]。这可能与几丁质具有结合肠道胆汁酸和游离脂肪酸的能力、促进脂肪转运有关[22]。与此结果相一致的是, FM20组加州鲈幼鱼的前肠脂肪酶活力最高, 显著高于FM25组, 胰蛋白酶活性也比对照组高, α-性淀粉酶却低于对照组, 这些可能导致FM20组加州鲈对饲料脂肪、蛋白质营养物质的消化吸收能力增强, 对饲料淀粉的消化能力降低, 继而促进鱼体生长及全鱼粗蛋白含量的增加。
由于黑水虻幼虫具有与鱼粉相似的氨基酸组成[6,21], 因此, 本研究显示用黑水虻幼虫粉替代0、16.67%、33.33%和50%鱼粉, 4组实验饲料的氨基酸组成相差不大, 对加州鲈幼鱼肝脏和肌肉的氨基酸组成也影响不大[21]。但是, 由于黑水虻富含中链饱和脂肪酸月桂酸、单不饱和脂肪酸C18∶1n-9和多不饱和脂肪酸亚油酸, 缺少EPA、DHA、ARA等高度不 饱 和脂肪酸(Highly unsaturated fatty acid,HUFA), 亚麻酸(C18∶3n-3)含量低, 因此n-6/n-3脂肪酸组成不平衡[5]。本研究结果显示, 在加州鲈的肝脏和肌肉中, 饱和脂肪酸主要以C16∶0为主, 单不饱和脂肪酸主要以C18∶1n-9为主, 多不饱和脂肪酸主要以亚油酸和DHA为主。在肝脏中, 随着黑水虻替代鱼粉比例的增加, C14∶0和C16∶0含量降低, C18∶0含量升高, 而肌肉中C18∶0含量则降低, 表明黑水虻富含的月桂酸主要在肝脏中被用于分解供能和合成C18∶0, 而C18∶0则有一部分被用于继续合成其他更长链的脂肪酸。随着黑水虻替代鱼粉比例的增加, 加州鲈肝脏C18∶1n-9逐渐降低, 表明C18∶1n-9主要被用于分解供能。肝脏和肌肉的亚油酸含量基本呈上升趋势, 与配合饲料中亚油酸含量随黑水虻替代比例上升而逐渐增加相一致。值得注意的是, 黑水虻缺乏DHA等高度不饱和脂肪酸, 亚麻酸含量也低。但是, 随着黑水虻替代鱼粉比例的增加,加州鲈肝脏和肌肉的亚麻酸含量逐渐降低, 但却显著提高了肝脏的DHA含量, 在黑水虻幼虫粉最适替代水平组(FM20组), 肝脏的DHA含量达到最大值。上述结果表明, 加州鲈具有内源的将18C亚麻酸合成DHA的能力, 在饲料中含有适宜的亚麻酸水平(饲料中鱼油和鱼粉含有一定量的亚麻酸)时, 合成DHA等高度不饱和脂肪酸的能力增强[20]。此外,加州鲈肝脏的DHA含量远大于ARA含量, 因此, 加州鲈偏向合成n-3系列高度不饱和脂肪酸。但是,FM20和FM15组加州鲈肌肉的DHA含量却显著低于对照组。DHA等高度不饱和脂肪酸构成动物细胞膜的主要磷脂成分, 其在鱼类个体发生、生长、存活、色素沉着、对应激和疾病的抵抗及在脑、视觉、神经系统的发育和功能中发挥重要的作用[32]。鱼类一般倾向在体内保留DHA, 尤其是在脑、眼等神经组织中保留DHA, 所以, 鱼类这些组织中的DHA含量一般较高且保持稳定[33]。当鱼类从饲料中摄取的DHA加上内源合成的DHA的总量依然不足时, 鱼类可能倾向从肌肉等组织转运DHA到脑、眼等组织, 会造成肌肉DHA含量下降, 肌肉品质降低。之前的研究也发现, 当投喂抽提了脂肪的鱼粉和非鱼油的配合饲料12周后, 加州鲈幼鱼肌肉和肝脏的DHA含量均显著低于鱼油组或未抽提脂肪的鱼粉投喂组; 其肌肉的DHA含量低于肝脏的DHA含量, 也低于初始鱼的肌肉的DHA含量, 表明加州鲈在投喂不含有HUFA的配合饲料时, 肌肉和肝脏的DHA含量显著降低, 而肌肉的DHA含量低于肝脏的DHA含量, 揭示肌肉保留DHA的能力较低[34],与本研究结果相一致。这也提示, 需要在加州鲈配合饲料中提供足量的DHA和适宜水平的亚麻酸,以满足加州鲈对DHA的需求, 增强肌肉品质。黑水虻幼虫粉替代部分鱼粉导致肌肉中DHA等高度不饱和脂肪酸含量降低的现象也曾在虹鳟[9]中报道过。
4 结论
综上所述, 在含30%鱼粉的基础饲料中, 用黑水虻幼虫粉替代16.67%、33.33%和50%鱼粉, 对加州鲈幼鱼的生长性能无显著影响, 降低全鱼和肝脏的脂肪含量及血清甘油三酯含量, 对肠、肝组织结构有一定损伤, 对肌肉和肝脏的氨基酸组成影响较小, 但是显著增加肝脏的DHA含量反而降低肌肉的DHA含量, 表明加州鲈具有内源合成高度不饱和脂肪酸的能力。当配合饲料中用黑水虻幼虫粉替代33.3%鱼粉时, 加州鲈幼鱼的生长性能较佳, 肌肉脂肪含量和全鱼粗蛋白含量最高, 肝脏合成DHA的能力最强。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
广东第二课堂·小学(2017年1期)2017-02-20
辽金历史与考古(2016年0期)2016-02-02
辽金历史与考古(2016年0期)2016-02-02
哈尔滨学院学报(2015年8期)2015-12-14
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
