
布尼亚病毒核蛋白的结构与功能
2024-01-09 12:13李祥亮王朝溪欧阳松应
中国生物化学与分子生物学报 2023年11期
李祥亮, 王朝溪, 欧阳松应,3)*
(1)福建师范大学生命科学学院生物科学系, 福州 350117;2)华中农业大学水产学院水产动物医学系, 武汉 430070;3)福建师范大学南方生物医学研究中心, 福州 350117)
布尼亚病毒是有包膜,分节段的负链RNA病毒,对全世界的公共卫生事业有重大的影响[1]。布尼亚病毒种类繁多,包括近500个成员。繁多的种类使得抗布尼亚病毒药物的研发变得愈加困难,鉴于核蛋白(nucleoprotein,NP)的高保守性,目前,越来越多的研究者将注意力集中在布尼亚病毒的NP上。NP在保护病毒RNA和免疫抑制过程中发挥重要的作用,但其分子机制尚不完全清楚,针对NP的研究不仅有助于人们进一步了解病毒的生命周期,而且对于药物靶点的开发具有重大意义[2]。目前,已经有许多布尼亚病毒NP的结构被解析,从这些结构可以看出属于同科病毒的NP结构相似性较高。研究已经发现一些以NP末端臂及RNA结合口袋为靶标的抗病毒抑制剂,这对布尼亚病毒疫苗和药物研发具有重大意义。本文将从布尼亚病毒及其NP的背景、NP参与的免疫反应、NP的三维结构、NP药物靶点的预测以及NP的应用现状等方面来进行总结阐述。
1 布尼亚病毒的背景
布尼亚病毒目(Bunyavirales)包含的病毒种类丰富,据国际病毒命名委员会(International Committee on Taxonomy of Viruses,ICTV)最新发布的报告,布尼亚病毒目包括496个成员,分为14个科,60个属。14个科分别为沙状病毒科(Arenaviridae)、甲壳类林肯病毒科(Cruliviridae)、盘状病毒科(Discoviridae)、费莫病毒科(Fimoviridae)、汉坦病毒科(Hantaviridae)、利什曼原虫布尼亚病毒科(Leishbuviridae)、多足类湖北病毒科(Mypoviridae)、内罗病毒科(Nairoviridae)、泛布尼亚病毒科(Peribunyaviridae)、幻影病毒科(Phasmaviridae)、白纤病毒科(Phenuiviridae)、番茄斑萎病毒科(Tospoviridae)、图拉斯病毒科(Tulasviridae)和武汉蜈蚣病毒科(Wupedeviridae)。
布尼亚病毒以节肢动物、锯齿状动物为载体进行传播,并在自然界的吸血节肢动物和易感脊椎动物中循环[3]。近年来,布尼亚病毒目中的许多新兴病毒对人类健康产生了重大影响。例如,裂谷热病毒(Rift Valley fever virus,RVFV)和克里米亚-刚果出血热病毒(Crimean-Congo hemorrhagic fever virus,CCHFV)感染人类后,可引发严重的病毒性肾综合征出血热(hemorrhagic fever with renal syndrome,HFRS),死亡率高达30%[4]。中国中部的患者身上分离出的发热伴血小板减少综合征病毒(severe fever with thrombocytopenia syndrome virus,SFTSV)引起了高达10%~30%的死亡率[5]。感染这些病毒的病例主要人群为农民,且在发病前很可能有过野外工作的经历[6]。布尼亚病毒的主要传播媒介是蜱虫和蚊子,SFTSV和CCHFV甚至可以通过人与人之间的直接接触进行传播[7]。布尼亚病毒的感染已经成为一个不容忽视的公共卫生问题,因此,寻找有效且安全的药物对抗布尼亚病毒十分紧迫。
布尼亚病毒是有包膜、分节段的负链 RNA病毒。布尼亚病毒呈球形,直径80~100 nm,病毒基因组由3段负链RNA片段组成,根据长短分别命名为L、M和S。L段编码RNA依赖的RNA聚合酶(RNA-dependent RNA polymerase,RdRp);M段编码病毒包膜糖蛋白(glycoprotein,GP);S段编码病毒NP和1个非结构蛋白质[8](Fig.1A)。

Fig.1 Bunyaviruses, their genomic patterns and NP amino acid sequence alignment (A) Bunyavirus are enveloped, with GP arranged on the membrane exterior. The bunyavirus genome comprises three RNA segments (S, M and L). (B) NP amino acid sequence alignment of SFTSV, RVFV and TOSV
2 布尼亚病毒核蛋白简介
布尼亚病毒的基因组S、M、L可以与NP相互作用,形成NP-RNA复合体,即核糖核蛋白复合体 (ribonucleoprotein complex,RNP),RNP在病毒的复制和转录中发挥重要作用[9]。NP与RNA结合后作为一个整体既充当复制的活性模板,又保护RNA不被宿主酶消化。当病毒进入宿主细胞时,RNP可以保护病毒的基因组物质不被宿主免疫系统识别,从而使病毒的RNA再复制和转录成为可能。研究RNP的功能和形成机制对于明确病毒的复制和组装非常关键,且对新型药物靶点的开发具有重要意义。除此之外,在病毒的组装过程,NP也发挥着极大的作用,它能够与RdRp和GP相互作用,一起完成病毒感染周期。
布尼亚病毒目中同科的病毒NP在一级序列上有较高的同源性,而不同科病毒之间NP一级序列的同源性很低。白纤病毒科中的SFTSV和托斯卡纳病毒(Toscana virus,TOSV),与同是白纤病毒科的RVFV NP在氨基酸序列上显示出较高的同源性,多序列比对显示同源性分别高达38%和50%[10](Fig.1B)。除此之外,泛布尼亚病毒科中大多数病毒也表现出高度的氨基酸序列同源性。
3 布尼亚病毒核蛋白参与的免疫反应
NP是高免疫原性的病毒蛋白质,在宿主细胞免疫反应中发挥重要作用[11]。NP在免疫反应中的作用已经在许多其他病毒,例如狂犬病病毒、甲型流感病毒等中得以验证:狂犬病病毒NP可以抑制宿主细胞I型干扰素反应[12];传染性造血坏死病毒(infectious hematopoietic necrosis virus,IHNV)NP的过表达显著降低了诱导I型干扰素表达蛋白质(mediator of IRF3 activator,MITA)的水平,数据表明,IHNV NP可以通过降解MITA来抑制宿主细胞I型干扰素的产生[13]。布尼亚病毒NP同样可以参与宿主细胞免疫反应,NP的不同结构域发挥不同的作用。例如,沙状病毒科的拉沙病毒(Lassa virus,LASV)NP的 C-端结构域有外切酶活性,是NP抑制干扰素所必需的[14]。外切核酸酶结构域可以降解病毒复制过程中产生的双链RNA(double-stranded RNA, dsRNA),从而防止它们在细胞质中的积累以及被宿主细胞识别消化的风险[14, 15]。CCHFV NP和LASV NP在氨基酸序列和结构上有较高的同源性,但CCHFV NP不会像LASV NP一样,通过抑制干扰素调节因子3(interferon regulatory factor 3,IRF3)的激活途径来执行类似的免疫逃避反应。事实上,CCHFV NP在N-端结构域中含有1个内切酶活性位点,可能在免疫抑制中发挥重要作用[16, 17]。RVFV NP的结构和内切酶活性域与CCHFV也不相同,这表明每种病毒的免疫抑制机制可能会有所差异。
LASV NP可以与DEAD-box RNA解旋酶3(DEAD-box RNA helicases 3,DDX3)相互作用,进而调控感染期间宿主细胞和病毒的生命周期[18]。DDX3是一种抗病毒蛋白质,是ATP依赖的RNA解旋酶[19],在RNA代谢过程中参与转录和翻译等。DDX3通过其解旋酶结构域使dsRNA展开,然后NP与DDX3相互作用以将解旋酶活性招募到复制转录复合体(replication transcription complexes,RTC)中,以促进病毒RNA的合成,从而促进病毒复制[18]。DDX3也是宿主细胞针对病毒感染的先天免疫反应的一个组成部分,在核因子κB激酶抑制剂ε(inhibitor of kappa-B kinase ε,IKKε)及TANK结合激酶1(TANK-binding kinase 1,TBK1)的上游和下游发挥作用,导致β干扰素的发生[20]。病毒感染早期,DDX3协助病毒RNA复制;感染晚期,DDX3充当干扰素的抑制因子[18]。黑色素瘤分化相关分子(melanoma differentiation-associated gene 5,MDA5)是细胞内的异体RNA监测蛋白,属于视黄酸诱导基因I (retinoic acid-inducible gene I,RIG-I)样受体家族的重要成员。NP介导RIG-I和MDA5与病毒RNA结合后,激活IKKε/TBK-1复合物和IKKα/β复合物[21],这些复合物磷酸化IRF-3和核因子κB(nuclear factor kappa-B,NF-κB),进而启动β干扰素、α干扰素和细胞因子的表达[22]。双链RNA依赖的蛋白质激酶(double-stranded RNA-dependent protein kinase,PKR)是一种抗病毒蛋白质,一旦检测到病毒dsRNA,它会通过磷酸化核糖体真核起始因子2(eukaryotic initiation factor 2,eIF2)来抑制病毒蛋白质翻译起始[23]。PKR也可以被干扰素诱导的PKR蛋白激活因子(PKR-activating protein,PACT)激活,随后通过磷酸化其抑制亚基介导转录因子NF-κB的激活[24]。对病毒感染早期的研究表明,NP阻断了IRF-3的核易位和转录活性,从而抑制了I型干扰素的产生[25]。干扰素活性丧失是上游几个协同过程的结果[26]。因此,RIG-I和MDA5未激活IKKε磷酸化的级联反应,从而不能激活IRF3通路。PACT和PKR都未被激活,导致活跃的eIF2持续存在,使病毒蛋白质继续翻译并抑制NF-kB通路的激活[27]。沙状病毒科病毒NP-IKKε的相互作用阻断了其自催化活性和磷酸化IRF3的能力[28]。综上所述,NF-κB,RIG-I-MAVS和TBK1-IRF3等信号通路中的很多关键分子是NP的作用靶点,NP可以通过对其进行调控诱导免疫反应,进而影响病毒和宿主细胞的生命周期[29](Fig.2)。
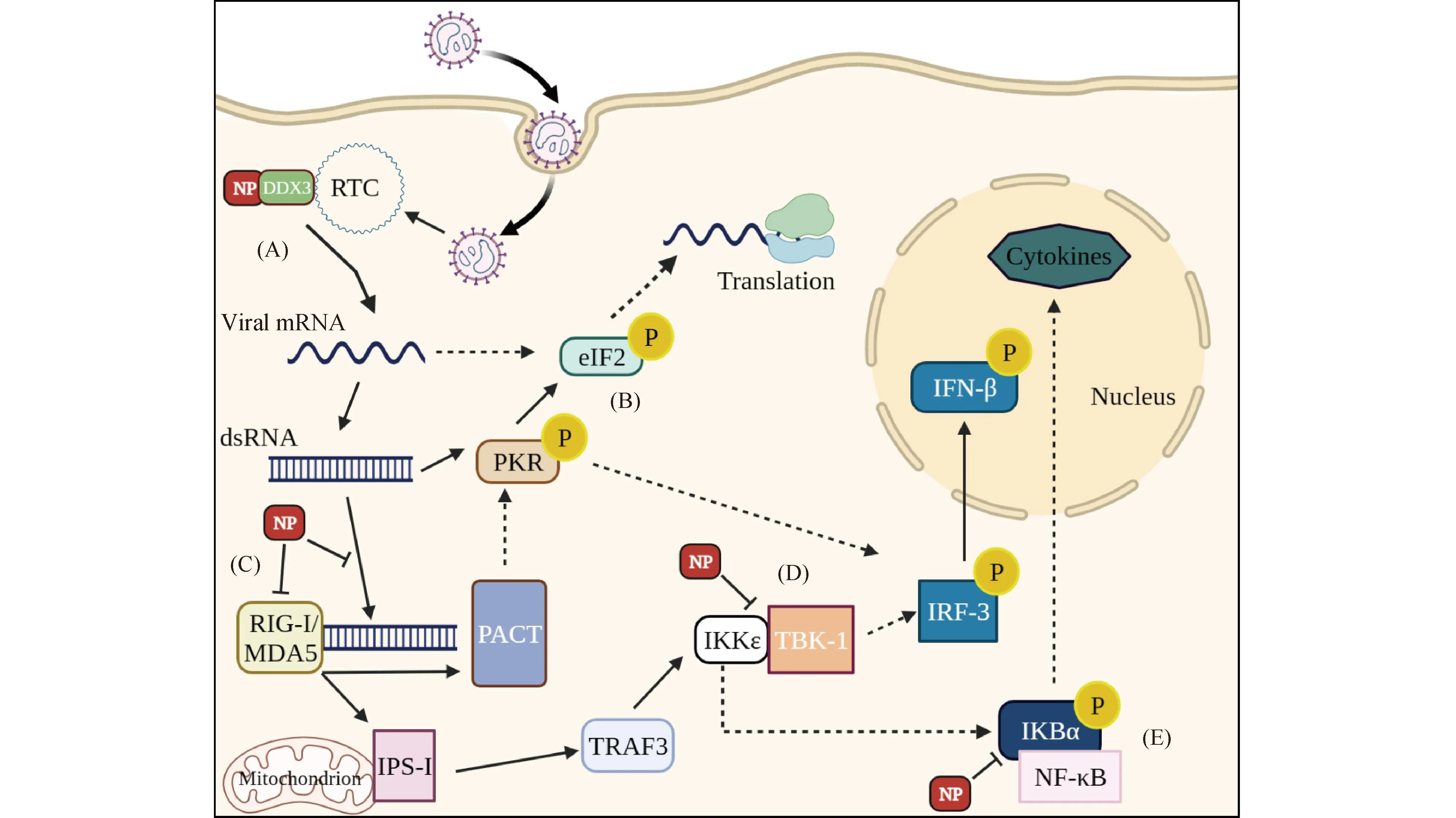
Fig.2 Diagram of the innate immune pathway and function of LASV NP (A) NP interacts with DDX3 to assist viral RNA replication, suppress IFN production and facilitate infection. (B) PKR phosphorylates eIF2 upon recognition of dsRNA and inhibits protein translation. (C) The interaction of NP with MDA5 transmits signals to downstream. (D) NP-IKKε interactions block its autocatalytic activity and ability to phosphorylate IRF3, NP blocks the nuclear translocation and transcriptional activity of IRF-3, which results in inhibition of IFN-I.(E)NP regulates phosphorylation of IKBα and NF-κB, prevents their nuclear translocation and inhibits cytokine expression
布尼亚病毒NP还可以同宿主细胞的许多其他分子相互作用,进而影响宿主细胞生命活动。最近的研究表明,汉坦病毒(Hantan virus,HTNV)感染宿主细胞后,NP与热休克蛋白40(heat shock proteins 40,Hsp40)竞争结合P58IPK(干扰素诱导的PKR抑制因子),导致Hsp40-P58IPK复合物解聚。随后,NP通过与核糖体小亚基RPS19相互作用,将活化的P58IPK招募到核糖体40S亚基抑制PKR的抗病毒作用[30]。
4 布尼亚病毒核蛋白的三维结构
NP是RNP的主要组成部分,RNP在病毒中可以折叠成双螺旋结构,以保护基因组RNA不被消化[31]。与GP相比,NP在病毒中具有较高的保守性,是研发抗病毒药物的一个重要靶蛋白。NP在病毒的生命周期中参与很多过程:病毒RNA的包被和保护;转录、复制过程的调节以及诱导宿主免疫抑制[1]。到目前为止,布尼亚病毒目中许多病毒NP单体结构和结合RNA的复合物结构已经被解析。RVFV NP结构是布尼亚病毒目中最先解析的[32]。RVFV NP的结构显示,其N-端臂可以介导蛋白质寡聚作用,每一个NP单体的N-端臂与相邻单体的背面相互作用,形成一个6聚体环(Fig.3A),在LASV NP的结构中C-端臂发挥类似的功能[16]。这可能是负链RNA病毒NP形成多聚体和包被RNA的普遍规律。

Fig.3 Structure of several Bunyavirus NPs (A) Structure of RVFV NP hexamer (left) and monomer (right) (PDB: 4H5M). (B) Structure of LEAV NP tetramer-bound ssRNA (left) and monomer surface electrostatic potential (right) (PDB: 4J1G). (C) HTNV NP is a two-lobed structure, and the RNA-binding cleft is located at the intersection between the two lobes(PDB:5FSG). (D)Structure of the TSWV NP trimer (left) and monomer (right) (PDB: 5IP1). (E) Superposition of monomeric structures of RVFV NP (green,PDB:4H5M), SFTSV NP (blue,PDB:4J4U) and TOSV NP (purple,PDB:4CSF) in the family Phenuiviridae. (F)Superposition of monomeric structures of BUNV NP (green,PDB:3ZLA), LACV NP (blue,PDB:4BHH) and LEAV NP (purple,PDB:4J1G) in the family Peribunyaviridae
通过解析布尼维尔病毒(Bunyamwera virus,BUNV)NP-RNA复合物四聚体的三维结构,揭示了BUNV NP独特的结构形式。电镜研究表明,重组表达的BUNV NP-RNA复合物在溶液状态下会以四聚体、五聚体和六聚体的形式共同存在[33]。同年,本研究团队报道了与BUNV同属于泛布尼亚病毒科的Leanyer virus(LEAV)的NP-RNA复合物的晶体结构(Fig.3B),显示了与BUNV类似的保护基因组RNA的模式。从结构中显示,NP由N-端和C-端两部分结构域组成,它们中间形成强烈的正电荷沟槽,此部位结合并保护单链病毒RNA。同时,在NP的N-端和C-端分别伸出柔性手臂,分别搭在前后相邻的2个NP上,稳定蛋白质的多聚化形式[34]。HTNV是出血热肾综合征的病原体,HTNV NP的结构与泛布尼亚病毒科NP结构相似性较小。它的 RNA 结合口袋位于双叶(lobe)之间的交叉处,NP的寡聚化由与相邻单体结合的N-端臂和C-端臂介导(Fig.3C)[35]。布尼亚病毒目番茄斑萎病毒科的番茄斑萎病毒(tomato spotted wilt virus,TSWV),分布范围很广,可侵害1 000多种植物,包括双子叶植物和单子叶植物,给人类社会造成了重大的经济损失[36]。TSWV NP的单体由3部分组成:N-端臂、C-端臂和核心结构域。与其他负链RNA病毒的NP不同,TSWV NP形成不对称的三聚体环。NP的N-端臂和C-端臂与2个相邻NP的核心结构域相互作用,进而形成三聚体环[37]。通过解析TSWV NP与RNA复合物的晶体结构,发现不对称三聚体环的内部沟槽是RNA结合位点(Fig.3D)。这些特征与其他布尼亚病毒的NP相似。
结构叠加表明,白纤病毒科中SFTSV,RVFV和TOSV的NP结构是非常保守的(Fig.3E)。此外,在泛布尼亚病毒科中BUNV、拉克罗斯病毒(La Crosse virus,LACV)和LEAV的单体构象高度相似[38](Fig.3F)。而不同科的病毒,其NP序列和结构的同源性较小,甚至几乎无同源性。这表明,布尼亚病毒同一科的病毒NP包裹其基因组RNA并形成RNP的机制相似。这些结果对于进一步认识布尼亚病毒转录复制复合体形成的分子机制,以及针对布尼亚病毒引起的重大传染性疾病防控都具有重要的意义。
5 核蛋白抗病毒药物靶点的预测
布尼亚病毒目中大多数病毒都缺乏有效的疫苗和抗病毒药物,因此,揭示NP上的潜在靶点,以开发抗病毒药物和有效的疫苗,对于解决布尼亚病毒的感染问题至关重要。
NP是一种多功能和高保守的蛋白质,在寻找抗病毒药物靶点方面具有重要意义。甲型流感病毒中已经发现了具有不同作用机制的靶向NP抑制剂[39],目前发现的布尼亚病毒科的靶向NP抑制剂仍很少。由于布尼亚病毒的研究实验需要在生物安全等级为3级或4级的实验室中进行,许多实验室不具备研究条件,阻碍了疫苗的研发[40]。因此,保守的抗病毒药物靶点对于布尼亚病毒的防治具有重要意义。
5.1 以NP的末端臂为靶标
NP寡聚是通过其末端臂来介导的,末端臂可以与相邻NP相互作用进而形成寡聚体。通过对RNP的负染结果分析发现,与相邻单元相互作用的末端臂是影响RNP构象的关键因素[31]。许多突变研究表明,打破2个NP单体之间的相互作用不仅对NP形成寡聚有影响,而且对RNP活性也有影响[41]。SFTSV NP通过其N-端臂的疏水键和氢键与相邻分子相互作用,NP寡聚体是通过1个NP单体结合另1个NP单体的N-端臂实现的。删除SFTSV NP的N-端或将关键位点突变,会使得环形寡聚体无法形成[42, 43]。突变或删除BUNV NP的前10个残基,删除RVFV NP的N-端,都可以减弱NP的寡聚作用[33]。
由此可见,将NP形成寡聚的关键位点突变或者用末端臂的类似物进行抑制可以阻止NP的组装。甲型流感病毒的NP抑制剂是一种包含尾环区域的环形肽,作用于NP尾环结合口袋,可以破坏NP的三聚体化进而抑制病毒的复制[44]。布尼亚病毒也可以根据同样的机制设计抑制剂。CCHFV NP与短单链RNA孵育会导致构象发生变化并解聚成单体NP。因为与短单链RNA的孵育会引起NP茎部结构域(参与寡聚体的形成)的旋转,导致二聚体界面的空间位阻,破坏聚合[45]。由此表明,能够改变CCHFV NP构象的短单链RNA片段可能是CCHFV复制和组装的理想抑制剂。最近研究发现,莫罗尼白血病病毒10(Moloney leukemia virus 10,Mov10)可以通过与布尼亚病毒NP的N-端臂结合,阻断NP形成寡聚体,进而抑制NP与RNA或RdRp相互作用,从而抑制RNP的组装[46](Fig.4A)。这为以NP的末端臂为靶标开发抗病毒药物提供了新的见解。

Fig.4 Pattern of antiviral targets on NP (A) Diagram of MOV10 interaction with NP. (B) NP cannot bind RNA in its RNA-binding groove in the presence of suramin
由此可见,NP的末端臂可以作为小分子药物的作用靶点。布尼亚病毒各个科的序列和结构的保守性有利于针对此作用靶点的理想药物的开发。
5.2 以NP与RNA的结合口袋为靶标
NP最重要的功能之一是保护基因组RNA不被宿主细胞消化[47]。每个NP亚基都有一个深而窄的RNA结合口袋,这个结合口袋富含正电残基,口袋会随着NP的聚合在环内侧表面形成RNA结合槽[33]。布尼亚病毒目同科中的病毒可能具有相似的RNA结合位点,并且有类似RNA结合机制。白纤病毒科中RVFV的RNA结合在NP上17个氨基酸的正电荷区。将RVFV NP-RNA复合物结构与白纤病毒科中的其他病毒,例如SFTSV NP比对,发现存在一个相似的RNA结合口袋,这个结合口袋的多个残基高度保守[48]。将SFTSV NP的RNA结合口袋上的关键位点进行突变,导致NP几乎丧失了结合核酸的能力,无法进行转录和复制[49]。内罗病毒科的CCHFV NP的RNA结合位点中发现的关键残基,在整个科中都是保守的[50]。因此,针对NP的RNA结合口袋为靶标设计药物,以阻碍RNA与NP的结合,是非常有前景的。
研究通过虚拟筛选发现的萘普生,是甲型流感病毒的NP抑制剂。抑制机制是通过与流感病毒NP的RNA结合口袋结合,诱导靠近其结合位点的关键残基发生构象变化,干扰了RNP的自组装,阻止NP在病毒的生命周期中发挥作用[51-53]。近年来,苏拉明的发现证实了RNA结合位点在抗病毒感染中的重要意义。苏拉明是在抑制剂筛选研究中发现的非核苷类抑制剂,是首个报道的针对布尼亚病毒NP的RNA结合口袋的抑制剂,通过与SFTSV NP共结晶得到证实[54]。苏拉明位于SFTSV NP的RNA结合位点上,该位点与RVFV NP的RNA结合位点有相似的特征[55]。苏拉明通过氢键和静电作用与NP相互作用,结合部位由高度保守的残基组成。功能研究表明,SFTSV感染宿主细胞后添加苏拉明,对体外细胞中的病毒复制产生了明显的抑制作用。另外的研究显示,苏拉明能够抑制病毒复制过程中起始的特异性NP-RNA结合,以及抑制尚未确定的一个或多个后续步骤,并且苏拉明能够通过直接与RNA结合口袋结合来渗透并破坏NP-RNA复合物(Fig.4B)。这些结果证明,苏拉明可能对大多数布尼亚病毒的复制有抑制作用[56]。
由此可见,抑制RNA与病毒NP的结合是一种有吸引力的抗病毒治疗策略,因为在感染过程中,NP执行的所有功能都涉及到与RNA的结合。因此,NP的RNA结合位点是临床治疗中寻找抗病毒药物的关键靶点,具有很大的潜力,可能对遏制布尼亚病毒的爆发具有重大意义。
6 核蛋白的应用
在布尼亚病毒的生命周期中,NP有着不可或缺的地位。针对NP的结构和功能方面进行的研究,能帮助人们更好的认识和了解布尼亚病毒复杂的生命周期和致病机制。所以研究布尼亚病毒的NP,不仅有基础层面的意义,还拥有更大的应用价值,例如疾病的诊断、疫苗的研发、靶向NP抗病毒药物的开发以及疾病的治疗等(Table 1)。
6.1 NP在疾病诊断中的应用
对于布尼亚病毒的检测已经有很多方法,包括通过酶联免疫吸附试验(enzyme linked immunosorbent assay,ELISA)检测IgM和IgG、病毒中和试验、具有高特异性和灵敏度的定量逆转录PCR以及进行病原体分离等[57]。但这些方法都存在某些不足,核酸检测仪器设备价格昂贵,极易引起交叉污染;IgM抗体一般需要在感染后5~6 d后才出现。胶体金免疫层析(immunochromatographic assay,ICA)使用胶体金标记的抗原或抗体作为示踪剂,分别检测抗体或抗原[58]。虽然胶体金技术检测方法简单快速,不需要仪器设备,但是不能做到定量检测,灵敏度还有待提高。

Table 1 Applications of NP and their characteristics
SFTSV NP的抗体在感染后早期可以检测出来,表明NP是早期诊断SFTSV感染的一个候选靶抗原。迄今为止,重组抗原已被用于准确、特异地检测布尼亚病毒的一些病毒的抗体。研究通过使用荧光碳点/SiO2实现了对SFTSV的超灵敏检测,该测定法对 SFTSV表现出快速、灵敏和高度选择性的反应,并用于检测实际样品,为 SFTSV 检测开辟了新的可能性[59]。目前,已经利用SFTSV的重组NP建立了一种间接ELISA方法,用于特异、敏感、安全和快速的检测SFTSV[60]。CCHFV NP可以在昆虫、细菌和哺乳动物、植物细胞中表达,能作为蛋白质抗原来检测病毒的特异性抗体[61]。基于CCHFV NP的ELISA检测病毒感染的方法亦可能在全球范围内应用[62]。
6.2 NP在疫苗研发中的应用
NP是最丰富的高免疫原性的病毒蛋白质。在一些动物模型中,NP靶向的疫苗可以诱发提供基础保护的细胞免疫反应。NP似乎在加强或激发某种程度的保护方面有一定的作用。
近年来,研究的重点是将布尼亚病毒的NP作为疫苗开发的潜在目标。NP是布尼亚病毒的高免疫原性的蛋白质,能刺激特异性抗体的产生,具有出现早、滴度高和时间长的特点,尤其是它还能诱发保护性体液免疫和细胞免疫[1]。RVFV、CCHFV和HTNV的DNA疫苗在各种组合的小鼠模型中都进行过试验。汉坦病毒科的普马拉病毒 (puumala virus,PUUV)NP显示了一个特殊的N-端区域,这个区域有助于保护模型免受后续PUUV的感染。在80%动物中,诱导保护的PUUV NP的N-端区域为1~45aa,而且更有助于免疫反应的区域被确定[63]。更重要的是,NP可以诱导高度交叉反应的抗体反应。免疫反应的选择性压力低于GP。此外,已经证明RVFV NP是一种有效的人类T细胞免疫原,可以诱导广泛的免疫优势的CD8+T细胞反应[64]。将构建的质粒DNA进行泛素化修饰可增强DNA疫苗的效力,这可能会解决未修饰的抗原与NP显示出较弱的细胞反应的问题。与编码RVFV GP的重组DNA疫苗相比,用pCMV-Ub-N(一种可编码泛素化NP的质粒)免疫小鼠,可显著提高干扰素受体基因敲除的小鼠的存活率[65]。在CCHFV中也观察到了类似的现象,即编码泛素连接的CCHFV GP和NP的DNA质粒,可更有效地促进细胞反应,对干扰素受体基因敲除的小鼠的致命CCHFV感染,表现出100%预防性免疫力[66]。由此表明,NP靶向DNA疫苗是一种潜在的候选疫苗,并为针对某些布尼亚病毒的疫苗设计提供了新策略。
7 问题与展望
布尼亚病毒引发一些人类疾病综合症,例如出血热、脑炎和致命的呼吸道综合症等,通过节肢动物和啮齿动物传播。布尼亚病毒种类繁多的原因在于它的基因组容易发生突变,且基因组的分段性质增加了病毒成员之间基因重排的可能性。随着病毒的进化,可通过与阳性患者直接接触而在人与人之间传播。由于病毒的传播途径越来越广,传播速度在不断加快,所以对人类和牲畜的健康构成了严重的威胁。另一方面,由于许多布尼亚病毒主要是以宿主动物为载体进行传播,因此,彻底消灭这些病毒是非常困难的。传统的减毒疫苗也可能由于与自然界的野生型毒株进行基因重排而恢复或增加其致病性。目前,抗病毒药物主要有两种类型,包括病毒靶标和宿主靶标。病毒靶标类型的药物可能对不同血清型的病毒产生不同的抑制作用。尽管利巴韦林和法匹拉韦已被证明对某些布尼亚病毒有抗病毒作用,但其抗病毒作用是有限的[67, 68]。所以,寻找有效的保守的布尼亚病毒疫苗或抗病毒药物是非常迫切的。
NP是一种多功能与高序列保守的蛋白质,是一种理想的抗病毒靶标蛋白,已广泛应用于病毒抗病毒药物的开发。NP为病毒的治疗提供了许多机会,并正在发展成为一个高度特异的抗病毒靶点。在本综述中,我们讨论了布尼亚病毒NP预防或治疗的潜在药物靶点。正如所讨论的那样,布尼亚病毒中同科的NP结构最为相似,特别是白纤病毒科和泛布尼亚病毒科,有利于这种理想的药物靶点的开发。另外,苏拉明也被证明可以分别与RVFV NP和SFTSV NP结合,通过与RNA结合口袋相互作用,破坏RNP的组装,干扰病毒的转录和复制。这进一步证实了NP可以作为理想的抗病毒靶蛋白,针对布尼亚病毒NP的研究,将有望有效的控制布尼亚病毒的感染和爆发。
猜你喜欢
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
环球时报(2019-07-09)2019-07-09
中国医药指南(2019年3期)2019-01-07
中国继续医学教育(2015年6期)2016-01-07
实用皮肤病学杂志(2015年4期)2015-12-22
哈尔滨医药(2015年6期)2015-12-01
中国医疗美容(2015年2期)2015-07-19
医学研究杂志(2015年12期)2015-06-10
西安交通大学学报(医学版)(2015年4期)2015-02-28
