单细胞视角下铅的体内代谢与检测
2024-01-09 02:54霍诗薇田皓中张伟灿胡立刚江桂斌1
中国无机分析化学 2024年1期
霍诗薇 刘 念 田皓中 张伟灿 郭 华 李 裕 胡立刚,4* 江桂斌1,
(1.中国科学院生态环境研究中心,环境化学与生态毒理学国家重点实验室,北京 100085;2.中国科学院大学 中丹学院,北京 100190;3.中国科学院大学,北京 100049;4.国科大杭州高等研究院 环境学院,杭州 310024)
尽管以含铅汽油为代表的铅污染问题不断得到治理,但是慢性铅暴露仍然是影响人体健康的重要环境问题。环境中新出现的铅污染源与人体内的骨铅蓄积,使得铅的环境外暴露和人体内暴露情况日趋复杂。血铅作为人体铅暴露风险的主要评价指标存在一定的局限性。因此,铅暴露的复杂性以及有关研究手段的缺乏为铅研究带来了新的挑战。随着单细胞技术的迅速发展,单细胞水平上铅的检测技术可用于建立和完善铅动力学的机制与模型,从而在单细胞水平上理解人体内的铅循环与铅的毒理效应,有望为控制铅污染的健康危害提供新的系统性认识。
1 铅污染与铅暴露来源
铅的污染与暴露一直是广受关注的环境健康问题。铅污染的来源主要包括矿产的开采冶炼及含铅汽油和电池的使用等,其中含铅汽油的大规模应用给人类健康造成了严重的危害[1]。从20世纪90年代开始,全球大部分国家陆续停止使用含铅汽油。2021年7月,随着最后一个国家——阿尔及利亚停止供应含铅汽油,联合国环境规划署随即宣布,全球已正式停止使用含铅汽油[2]。含铅汽油的禁止使得全球范围内成人和儿童铅中毒的患病率显著降低[3-5]。尽管含铅汽油这一里程碑式的铅污染问题已经解决,来源复杂的铅污染所造成的公众健康威胁仍十分严重,且面临诸多新的挑战。因此,为厘清人体铅暴露的来源,本章节通过分析铅污染现状,总结铅暴露源特征以及新的铅暴露源,为有关铅暴露的分析、毒理等环境健康研究提供新的参考,以便控制和削减其产生的环境风险和健康威胁。
1.1 铅污染现状与铅暴露源特征
近十年来,铅减排的相关举措有力地削减了环境中的铅暴露源,但铅污染对人体健康的风险仍然存在且不容乐观。例如,联合国儿童基金会发布报告显示,全球大约三分之一的儿童(约8亿)的血铅水平超过安全限值[6]。美国毒物与疾病登记署结合各种有害物质的毒性、频率及接触可能性,将铅列为有害物质优先清单第二名[7]。
人体从环境中接收的铅暴露源主要分为两类:人类活动直接释放的铅与留存在环境中的铅。铅矿的开采与加工是人类活动直接释放的主要铅暴露源之一,且并未随着铅减排的推行而停止。2011年至2021年间,全球铅的年开采产量约为400万t至500万t,铅的年精炼化产量(包括新开采的铅与回收的铅)约在1 000万t至1 100万t[7]。人类活动直接释放的铅暴露源存在典型的区域性特征。例如,非洲地区的铅污染问题主要归因于其矿产资源的不当开采[8]。在以美国为首的部分国家及地区,含铅管道的安装引起了严重的日常用水中的铅污染问题[9]。在中国,铅矿的开采量和冶炼量遥遥领先于世界[7],同时中国还是煤炭的生产和消耗大国,因此铅生产以及燃煤导致的铅排放是中国铅污染的主要来源[10]。
环境中留存的铅可通过人类活动再次释放,成为人体接收的铅暴露源。冰芯中铅浓度的数据显示,工业革命后铅的环境浓度呈现上升趋势,2000年铅的环境浓度约为工业革命时铅浓度的3倍且远高于工业革命前,因此铅在环境中的积累以及再释放导致的浓度上升不容忽视[11]。
1.2 新的铅暴露源
铅已被广泛应用于光伏电池和风力涡轮机的储能,如铅酸电池与铅基钙钛矿太阳能电池[12-13]。随着可再生能源的普及,全球储能需求增加,新的铅暴露源不断涌现。在很多中低收入国家,不受管制的铅回收活动也因此成为了环境铅污染与人体铅暴露的重要来源[13]。铅基钙钛矿作为目前最有效的光伏材料,其潜在的毒性已被验证[12,14],如果发生泄露,遇水会释放高水溶性与高生物利用度的碘化铅[15],该化合物的生物有效性约为其他含铅污染物的十倍[16]。随着全球范围内对清洁能源的推广使用,铅基钙钛矿可能成为一种不容忽视的铅暴露来源。中国是全球铅酸电池的主要生产国、精炼国和消费国[17],同时随着“双碳”政策的推行,新的铅污染源的防治将成为我国铅暴露风险防控中所要面临的重要挑战。
2 单细胞水平上铅的动力学机制
环境中的铅污染短期内难以根治,人体面临着持续性的外源铅暴露,由此导致铅在人体内不断积累,对人体健康可能产生不可逆转的负面影响。传统的铅动力学研究主要关注铅在区室间的周转,仅以控制铅的摄入来减少铅对人体的危害,因此人体内铅动力学的研究亟需完善单细胞水平上的循环机制,进而寻找富有临床意义的铅排出路径,为解决铅暴露引发的健康问题提供重要支撑。本章节较为详细地描述了人体内铅的循环路径,对现有的单细胞水平上的铅动力学机制进行总结,并进一步探讨其研究意义,为建立单细胞水平上铅的动力学模型以及人体内铅的排出方法提供新的思路。
2.1 传统的铅动力学机制
铅主要通过呼吸道与胃肠道进入人体[18]。肺部的铅颗粒通过肺泡毛细血管膜进入血液,其余的颗粒会被肺泡巨噬细胞或淋巴系统清除[7,19]。胃肠道部分的铅吸收与肠上皮细胞、肠壁排列细胞等有关[18,20],通过主动运输、被动运输以及胞饮作用进入肠道组织[20-21]。在运输过程中,铅与其他金属竞争运输蛋白以及结合蛋白,因此铅的吸收受人体摄入的营养素影响[18,22]。
铅分布过程中,血液将铅运送到各个靶器官,起到关键的连接与运输作用。血液中约99%的铅被红细胞吸收[23],其余的铅在血浆中进行传输,血液中的大部分铅与蛋白质或者配体络合[24-27]。骨骼以不溶性磷酸盐的形式储存铅,承担了人体中约95%的铅含量[23]。铅可以随新骨生成储存在骨矿物基质中。同时,铅以异质离子交换返回血浆,或者在破骨细胞主导的骨吸收过程中释放返回血液[28-31]。软组织中的铅主要与蛋白质结合,目前已有研究在大鼠的大脑与肾脏中发现了与铅亲和力很高的铅结合蛋白[32-33]。
人体内不同形式的铅存在不同的代谢途径。烷基铅会在完全氧化后生成无机铅,四甲基铅和四乙基铅会生成有高度神经毒性的代谢物[34]。无机铅通过配体反应来进行代谢,可与各种蛋白质、氨基酸以及非蛋白硫醇结合形成配合物[18]。
铅经过肾小球滤过作用和近端肾小管重吸收作用随尿液排出人体[35-36]。粪便和唾液等均是铅的排出途径[37],如图1所示。
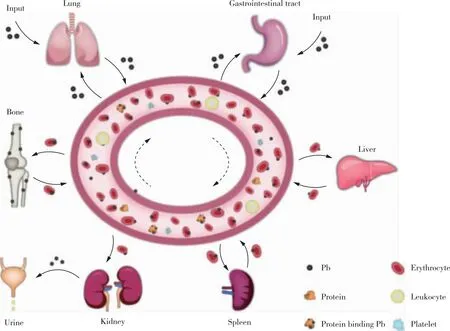
图1 铅的动力学机制示意图Figure 1 Schematic diagram of kinetic mechanism of lead.
2.2 单细胞水平上铅动力学机制
随着单细胞技术的发展,厘清铅传输与铅分布的细胞机制,完善单细胞水平上的铅动力学机制,成为探究人体内铅动态变化以及改善铅中毒治疗效果的一种有效途径。
铅在生物体内的细胞转运机制与大多数离子相近。血液中的铅通过介导Cl-与HCO3-的阴离子交换蛋白[38],以及钠泵与钙泵的阳离子转运机制进入红细胞[39-40]。研究发现,铅可以通过钙离子通道进入大鼠垂体瘤细胞(GH3细胞)与大脑的神经胶质细胞[38,41-44]。阴离子交换蛋白也参与了脑部星形胶质细胞的铅运输过程[38,41-44]。利用体外建立的血脑屏障模型,可以证明二价金属离子转运蛋白1在血脑屏障中铅转运的介导作用[38,41-44]。肾脏中的铅通过二价金属间共同存在的竞争性转运蛋白进行转运,包括锌转运蛋白以及二价金属离子转运蛋白1,但具体细节仍存在争议[44]。
随着细胞异质性的不断验证[45-46],同一类型细胞对同种污染物的不同反应也与人体内的铅动力学高度相关。细胞异质性所导致的随机过程会在微观角度上影响铅的转运与分布,但铅总量层面的检测与研究忽略了细胞的异质性,这也是当前铅治疗效果受限的关键所在。铅的总量分布差异可以间接地体现细胞间的铅含量存在异质性。不同组织中铅的分布显然不同,但同一组织中的不同区域同样存在铅分布的差异性。如,肺部不同肺段累积的铅浓度存在显著差异性,区域淋巴结中累积的铅浓度则明显高于肺部平均铅浓度[47]。骨骼中的铅在不同部位的积累机制不同,其浓度分布与该部位的矿化程度有关[48-50]。更为直观的是,有研究发现,胎儿血红蛋白特有的γ链对铅的亲和力相比成人血红蛋白中的α或β链明显增强[25-27]。近期的一篇研究发现,血液中单个红细胞中的铅含量呈现差异性分布。其中,成熟红细胞中的铅含量明显低于网织红细胞中的铅含量。由于红细胞主要在骨髓生成,网织红细胞是红细胞的前体,说明随着血细胞的分化成熟,单个血细胞中的铅含量呈现逐步下降的趋势[51]。因此,血细胞极有可能在骨髓生成阶段吸收大量铅,随着其分化成熟逐渐释放铅。
细胞异质性不仅体现在污染物的分布吸收中,还体现在污染物对不同状态细胞的损伤程度上[52]。这也表明,随着单细胞研究的不断深入,不同细胞对某一类型的化学污染物的反应差异是可预测的[51]。现有的铅动力学机制多数从组织层面进行解读,单细胞水平上的铅动力学机制研究占比较少,所以忽视了铅分布的细胞异质性及其他在人体健康风险中所起的作用。因此,需要完善单细胞水平上铅的动力学机制,揭示各种随机过程导致的细胞间的铅分布差异性,形成单细胞水平上铅循环的闭环路径,从而对铅暴露可能造成的细胞毒性进行预估,进一步对各种临床检测的低水平铅暴露症状进行单细胞水平上的溯源和治疗。
3 铅的动力学模型
根据铅的动力学机制与模型,人体内的铅循环可主要划分为三个区室:血池、骨池和软组织池[37,53-56]。随着铅的生物动力学、骨骼中矿物质的代谢规律等相关的研究进展,建模工作得以进一步完善和扩展。目前,在铅风险评估中广受认可并且被广泛应用的动力学模型有三种:1)O’Flaherty模型;2)美国环境保护署提出的儿童铅综合暴露吸收生物动力学模型;3)Leggett模型[18]。O’Flaherty模型主要考虑了年龄因素对模型的影响,可应用于广泛年龄范围内的人体血铅浓度评估[31,57-60],如图2(a)所示;儿童的铅综合暴露吸收生物动力学模型则主要应用于0~7岁儿童人群,其中包含对于铅暴露来源的详细分类[61],如图2(b)所示;Leggett模型可以广泛适用于碱土元素以及动力学机制与钙类似的相关元素,其区室划分细致,被认为是铅药代动力学中较为先进的模型[62],如图2(c)所示。
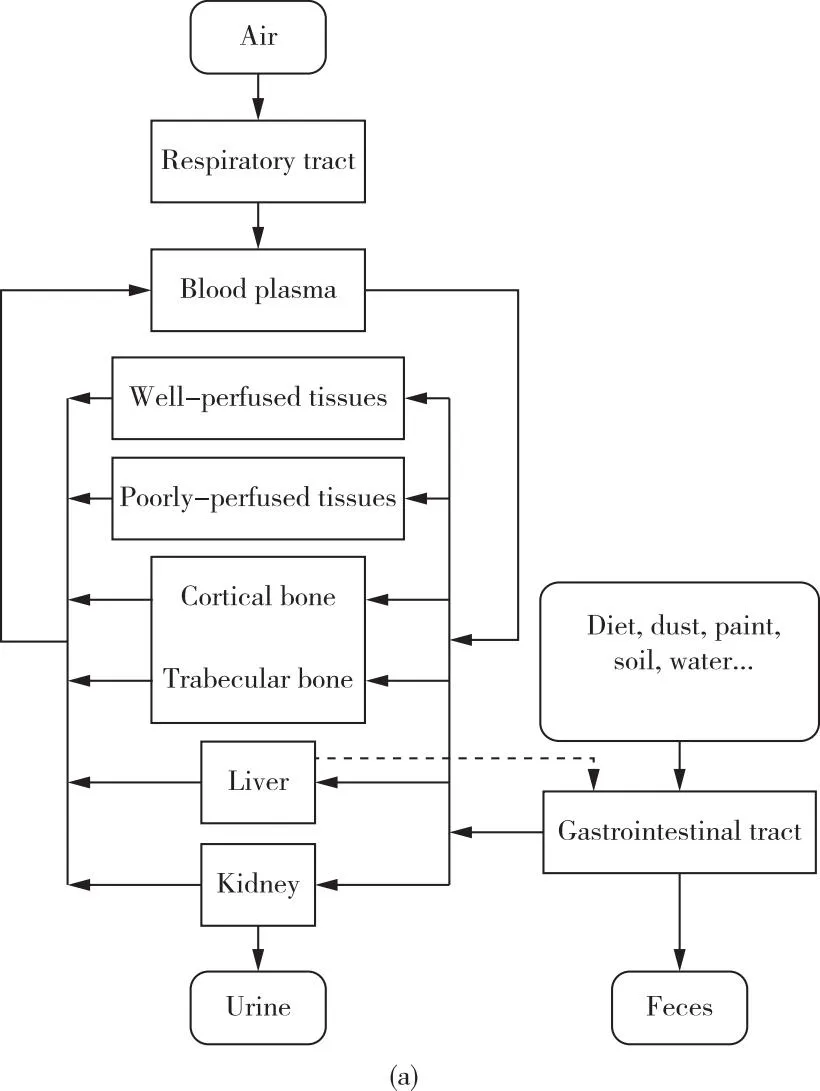
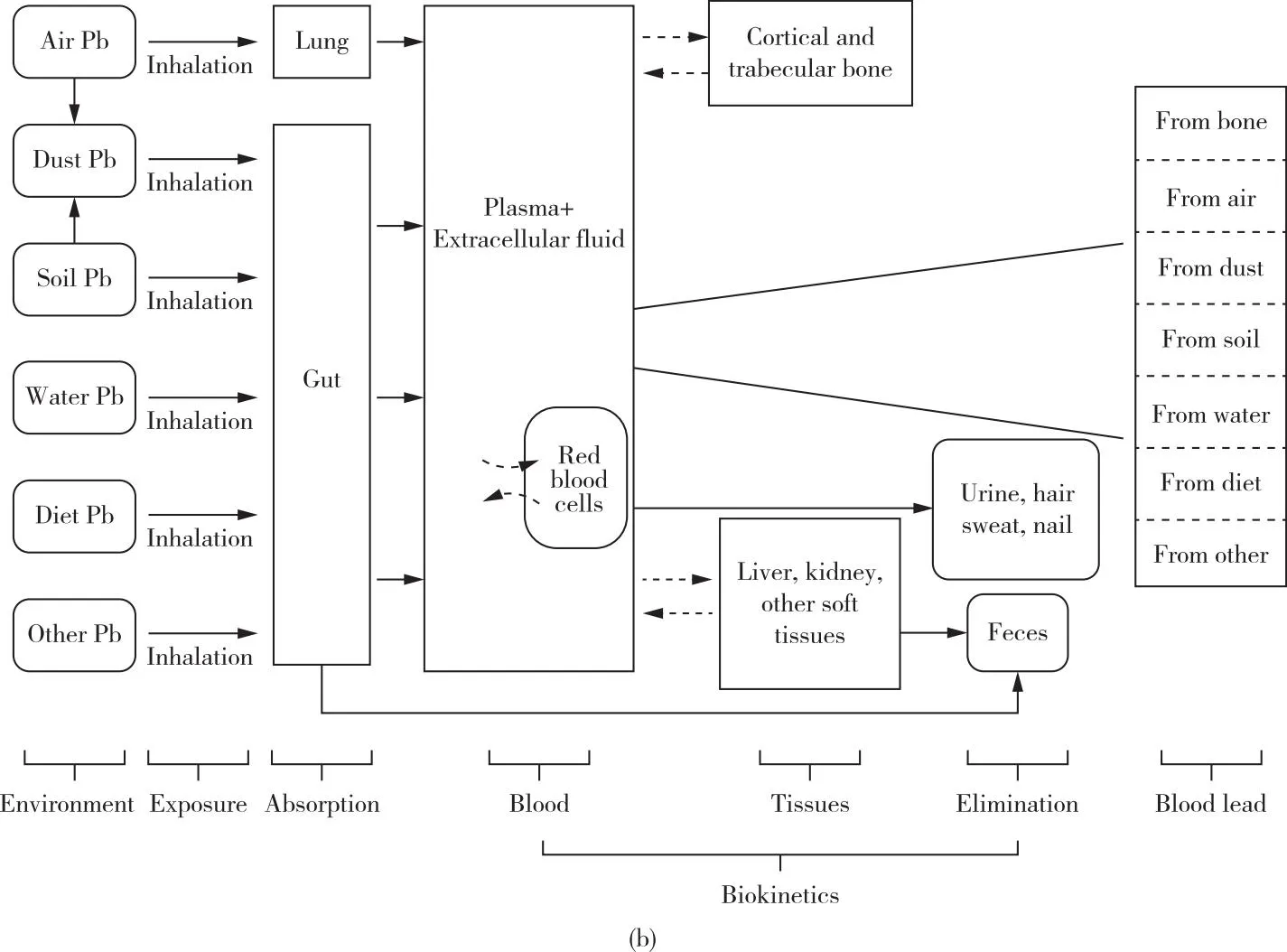
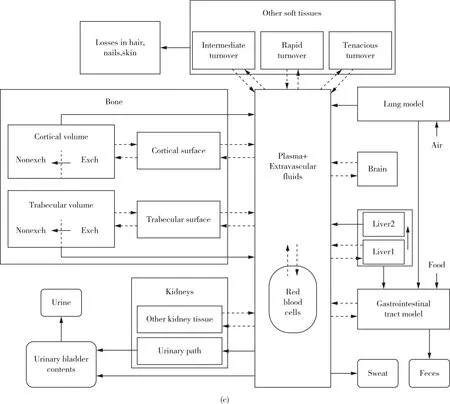
图2 广泛应用的铅动力学模型示意图[31,57-62](a)O’Flaherty模型;(b)儿童的铅综合暴露吸收生物动力学模型;(c)为Leggett模型Figure 2 Schematic diagram of lead kinetic models widely used[31,57-62].(a)O’Flaherty model;(b)The integrated exposure uptake biokinetic model for lead in children;(c)Leggett model
在铅动力学模型由三区室向多区室发展的过程中,模型中不同铅周转率的区域划分更加详细,越来越多的影响因素被纳入考量,如表1所示。有研究以钙与铅的竞争性动力学为主导,尝试将分子机制转化为具体的数学描述。但该模型同样存在一定的局限性,例如,缺少更加具体的生物学基础、分子和细胞机制作为理论依据[63]。

表1 不同铅动力学模型的特征
值得关注的是,铅动力学模型的建立愈加重视细胞机制的作用,考量不同离子的交换作用及元素竞争作用。模型的发展趋势也更加体现了铅动力学领域对细胞、分子水平上铅转运与铅分布机制的探寻与需求。当前的铅动力学模型将复杂的运输机制转化为不同区室之间的周转率,平均化地预估各个区室的铅总量水平,忽视了单细胞水平上存在的分布差异。单细胞水平上的铅动力学模型将形成单细胞水平上铅转运与铅分布机制的闭环,结合细胞机制与细胞的异质性进行研究,完善人体内单细胞水平上的铅运输路径,有针对性地识别铅相关毒理效应的诱因,构建铅从输入到传输再到排出的完整通路,并且根据铅动力学的运输与分布机制进行引导,建立临床上的高效治疗方法。单细胞水平上的铅动力学模型需要更为成熟的单细胞检测技术支撑,并且借助相关的大数据研究与数学建模方法对检测得到的批量单细胞数据拟合分析,实现体内铅暴露来源的定位追踪,从而完成人体内铅的溯源。
单细胞水平上铅动力学机制的完善以及铅动力学模型的建立对单细胞检测技术提出了新的要求,而传统的铅检测技术无法实现细胞机制与细胞异质性的研究,单细胞水平上的铅检测技术成为了铅的单细胞研究的关键所在。
4 单细胞水平上铅的检测技术
传统的铅检测方法已经具有较高的灵敏度和准确度,但只能得到样本中铅的总量或者平均指标,无法获得单细胞层面的信息。单细胞水平上铅的检测技术可以弥补传统检测方法的缺陷,探索单细胞层面的传输与作用机制,更好地体现单细胞分析所强调的细胞异质性。本章节总结了现有单细胞水平上的金属检测技术,并对这些技术及其应用进行简要介绍,以便为人体内单细胞水平上铅的细胞机制研究提供有效的工具。
4.1 流式细胞技术
细胞流式技术是当前铅研究领域应用最广且较为成熟的单细胞检测技术。该技术通过荧光探针将荧光基团与金属连接,再利用流式细胞技术进行分选与分析,从而实现单细胞层水平上的金属检测。得益于发展较为成熟的荧光成像技术,目前已有较多用于检测铅及其他金属的荧光探针[64]。相比其他体外检测或确定细胞内是否含铅的荧光探针,目前的最新研究构建了一种基于若丹明的荧光探针RPb1,可用于细胞内不稳定铅的检测,同时利用结直肠腺癌细胞(DLD-1细胞)对其与铅结合的灵敏度进行验证。随后该研究利用流式细胞技术对不同分化程度的人慢性髓系白血病细胞(K562细胞)中的不稳定铅进行检测,结果表明,随着细胞分化程度的提高,蛋白结合铅的水平增加[65]。
4.2 质谱流式细胞技术
质谱流式技术将流式细胞仪与质谱仪进行结合,利用重金属同位素标记与抗原特异性结合进行鉴定区分,解决了多参数检测中荧光发射光谱较宽导致的重叠问题,同时实现了细胞亚群分析与多参数分析[66]。该技术方法在与免疫相关的研究中已经发展得较为成熟,在金属领域的应用仍然需要拓展。目前有研究利用质谱流式技术与单细胞数据的拟合方法对铅中毒患者与健康受试者中的铅分布模式进行分析,发现个体成熟红细胞中的铅含量具有高度异质性,铅中毒患者与健康受试者的单个成熟红细胞中的铅含量均符合伽马分布,且血细胞中的铅含量随着其分化程度的上升而下降。该研究也为铅的体内循环研究提供了单细胞水平研究上的新思路[51]。
4.3 基于电感耦合等离子体质谱的单细胞检测技术
电感耦合等离子体质谱技术作为近些年金属检测应用最广泛的研究技术,其灵敏度得到了普遍认可[67-68]。目前单细胞水平上可应用的金属检测技术包括单细胞电感耦合等离子体质谱技术、电感耦合等离子体飞行时间质谱技术和激光烧蚀-电感耦合等离子体质谱技术等。
单细胞电感耦合等离子体质谱技术根据稀释法及微流控等方法,利用单个细胞间的充分分离来得到非连续信号,实现单细胞水平上的金属检测,其信号强度与单个细胞中的金属含量成正比[69]。有研究采用单细胞电感耦合等离子体质谱技术测定了单个酿酒酵母细胞中的银浓度[70],也有研究使用该方法实现了单个细胞中的多种内源性金属元素的准确测定[69]。
电感耦合等离子体飞行时间质谱可以实现大量分析物中短瞬态信号的多元素测量。根据电感耦合等离子体飞行时间质谱的单细胞分析方法,有研究探究了包括铅在内的6种重金属对酵母细胞吸收磷的影响[71]。已有研究利用电感耦合等离子体飞行时间质谱技术实现了高达16种金属的同时间多元素特异性检测[72]。
激光烧蚀-电感耦合等离子体质谱技术主要通过前处理方法生产单细胞阵列,用激光烧灼单个细胞产生气溶胶,气溶胶被引入电感耦合等离子体质谱实现单细胞水平上的分析。利用激光烧蚀-电感耦合等离子体质谱技术可以对单个巨噬细胞中的银纳米颗粒进行定量[73]。基于压电声学微阵列的单细胞阵列新技术,有研究首次利用激光烧蚀-电感耦合等离子体飞行时间质谱完成了单细胞内源性同位素的指纹图谱[74]。
但是,基于电感耦合等离子体质谱技术的单细胞分析方法仍存在较多挑战,需要标准化的技术方案[75]与前处理方法来实现该技术的定量检测。
4.4 单细胞水平上的其他金属检测技术
除以上较为系统的研究技术外,现有研究通过结合荧光探针与微流控制系统实现了单细胞中多种金属离子的分析[76],也有研究利用透射电子显微镜、X射线荧光显微镜、纳米二次离子质谱等实现了单细胞水平上金属的成像[77-79],但其成像精度使其应用范围受限,主要应用于细胞体积较大的植物细胞等。
虽然当前单细胞技术日益完善,但单细胞水平上铅的检测技术仍需进一步探索与完善,以便支撑铅暴露有关的单细胞研究。
5 结论与展望
可再生能源的日益普及,可能导致新的人体铅暴露来源;而体内骨骼对于铅的储存作用,可能使大量人群面临老年时期的体内铅释放风险,甚至在母体怀孕阶段威胁婴幼儿的发育健康,所以铅有关的人体健康风险研究仍十分重要。当前的铅检测技术可以实现高精度的铅总量检测,但单细胞水平上铅的检测技术仍有待推广应用。检测技术的局限性导致铅细胞机制的研究较为欠缺,细胞异质性的相关研究十分有限。多数单细胞水平的铅研究从毒理效应入手寻求其对应的细胞机制,以减少靶器官的毒理效应,忽略了铅动力学中可能存在的调控作用与细胞异质性作用,因此较少有研究致力于单细胞水平上铅的运输与分布。铅动力学模型依旧停留在组织层面的铅总量估算,缺乏对于铅动力学的动态考量,限制了铅循环完整体系的构建与铅临床治疗效果的改善。
铅循环中,血液作为关键的铅运输载体,在不同发育阶段直接接收不同的铅暴露,包括发育过程中人体骨骼的铅暴露以及成熟后外界环境直接摄入和吸入的铅暴露。而其他软组织则由血液进行铅的传输,间接性地暴露骨骼以及外界环境中的铅,这表明血液在铅的溯源机制中起到关键作用。铅污染造成的骨铅蓄积问题对于血液存在着暴露风险,我们对于人体内铅循环细胞机制的认识尚不成熟,无法对现阶段铅的暴露风险和健康效应进行预估,也无法进一步完成临床治疗中铅的溯源与根除性治疗。
随着单细胞技术和生物数学模型的发展,铅的研究重点应该逐渐由总量转移到细胞层面。从而完善铅动力学中的细胞机制,建立血液中铅运输的路径以及动态的动力学模型,探索铅分布异质性的影响因素与可引导的铅排出路径,提出有效的临床治疗方法。完善的单细胞理论基础,标准化和体系化的前处理与检测方法,仪器的联用和创新是其中的决定性因素。利用单细胞水平上铅的检测技术进行样品的分析,结合动物实验进行验证,将单细胞水平上铅含量与动力学机制及铅来源等信息相结合,能为我们解决单细胞水平上人体内铅动力学的未知机制提供新思路。
猜你喜欢
黑龙江大学自然科学学报(2022年1期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
科学(2020年4期)2020-11-26
当代化工研究(2016年5期)2016-03-20
浙江大学学报(工学版)(2015年2期)2015-05-30
生殖医学杂志(2015年11期)2015-02-28
化学工业与工程(2015年1期)2015-02-10
特产研究(2014年4期)2014-04-10
火炸药学报(2014年1期)2014-03-20