
阳春砂与草果叶片解剖结构分析
2024-01-08 02:15杨加珍徐鹤萌曾亚文
湖北农业科学 2023年12期
杨加珍,徐鹤萌,陈 佳,何 伟,曾亚文
(1.云南省农业科学院生物技术与种质资源研究所/云南省农业生物技术重点实验室/农业农村部西南作物基因资源与种质创制重点实验室,昆明 650205;2.普洱学院,云南 普洱 665000;3.红河麟源农业科技发展有限公司,云南 河口 661300)
阳春砂(Amomum villosumLour.)与草果(Amomum tsaokoCrevost et Lemaire)均为豆蔻属多年生草本植物。阳春砂属亚热带半阴性植物,喜高温、高湿,适合生长在600~800 m 低海拔的热带、南亚热带温暖湿润适当阴蔽的阔叶常绿林地,大多栽培或野生于山地阴湿之处。阳春砂分布于云南、福建、广东、广西等省,云南省是阳春砂仁的主产区,其药材产量占全国的70%以上。草果适宜生长在热带、亚热带湿热阴蔽的阔叶林中和1 300~1 800 m 高海拔的林下阴湿处[1],主要分布在中国云南、广西和贵州三省的局部地区,在云南省主产于南部、东南部和西南部。叶是植物进行呼吸、光合、蒸腾作用的重要器官,是植物应对逆境胁迫最敏感的部位。生长在不同生态环境中的植物,表现出不同的形态结构,是植物对特定生境的演化适应[2]。叶片解剖结构能够反映植物对环境变化的响应和适应[3],叶片的表皮结构特征具有一定的遗传稳定性,在一定程度上叶片的表皮微形态特征可以反映植物类群间的亲缘关系,在植物的种间、种内分类上具有重要的研究意义[4,5]。气孔是植物进行蒸腾作用、光合作用和气体交换的重要通道,是研究植物间亲缘关系的重要器官[6,7],气孔研究已在许多植物中有报道。何荆洲等[8]揭示了25 个蝴蝶兰品种叶片的气孔特征指标作为形态指标和评价的重要依据。徐梅等[9]研究得出薯蓣属(DioscoreaL.)植物叶表皮微形态存在差异,可作为种间分类的依据。陈方永等[10]用杨梅叶片气孔的相似度作为区别杨梅品种的重要依据。王连珍等[11]揭示栎属(QuercusL.)植物叶片气孔的稳定性特征,可作为该属植物亲缘关系分析和分类的重要依据。近年来在草果与阳春砂在产业发展[12,13]、挥发油的提取[14]、化学成分的鉴定[15]、药理作用[16]、真假砂仁辨别[17]等研究方面取得了一定的进展,但对其解剖学方面的研究较为鲜见。黄桂玲[18]揭示了砂仁的叶片为背腹型叶,气孔常分布于下表皮,下表皮还含有油细胞,有一层栅栏组织,海绵组织较疏松,含有草酸钙结晶,叶脉表面的表皮中含有单个的圆球形的硅质体。唐源江等[19]研究豆蔻属(AmomumL.)的12 种植物叶片得出砂仁的远轴面和近轴面均有一层不连续的下皮层,且远轴面下皮层细胞体积小,近轴面下皮层细胞体积大;草果没有下皮层细胞,砂仁与草果的叶肉均有1 层栅栏组织,且砂仁叶肉组织含有少量方晶。唐源江等[20]对28 种姜科(Zingiberaceae)植物进行了研究,表明3 种豆蔻属(AmomumL.)植物的根表皮均由1 层表皮细胞组成,其中砂仁与草果的根含有表皮毛;皮层都由薄壁细胞组成,其中内皮层细胞壁均呈U 形增厚,野草果较明显,砂仁和草果则不明显。为了解豆蔻属2 种植物阳春砂与草果的形态特征及叶解剖结构,本研究采用指甲油印痕法和超薄切片法制片,对2 种植物叶片的上、下表皮气孔情况、叶片的横切面以及叶片超微观结构比较分析。探讨二者结构是否存在差异性及叶片形态特征的种间差异,揭示其叶片形态差异的多样性,以期为豆蔻属植物的分类和亲缘关系研究提供依据,也可以为适应温暖湿润及其阴蔽的林下山区沟谷林栽培提供解剖结构依据。
1 材料与方法
1.1 材料
1.1.1 供试植物材料 阳春砂植株,采自云南省红河州河口县老放寨乡沙仁基地(海拔约700 m),属于红河麟源农业科技发展有限公司;草果植株,采自云南省文山州马关县。
1.1.2 试验设备 超薄切片机(EM UC7 型)、倒置荧光显微镜(DMI 4000B 型)、玻璃制刀机(EM KMR3 型)、修块机(EM TRIM2 型)、组织处理机(EM TP 型),德国Lecia 公司;透射电子显微镜(TECNAI SPIRIT G2 型),美国FEI公司。
1.1.3 试验试剂 2.5%戊二醛固定液、0.1 mol/L PBS 缓冲液、1.0%锇酸、去离子水、无色指甲油、包埋剂、乙醇溶液(30.0%、50.0%、70.0%、80.0%、95.0%、100.0%)、丙酮、甲苯胺蓝染液、醋酸双氧铀染液、柠檬酸铅染液。
1.1.4 试验耗材 玻璃刀、滤纸、双面刀片、镊子、牙签、载玻片、2 mL 离心管、0.5 mL 离心管、盖玻片、胶头滴管、酒精灯、透明胶带。
1.2 方法
1.2.1 植物叶片上、下表皮气孔观察 采用指甲油印痕法[21]制片,取植物叶片中部叶脉两侧部分,切成1 cm×2 cm 的小块,在叶片上、下表皮分别涂一层无色指甲油,待风干后小心用透明胶带将叶片上、下表皮指甲油轻轻撕下,置于载玻片上制片,倒置荧光显微镜下观察并拍照,标记气孔数量、计数,测量气孔大小及气孔开度。每片叶片制作3 张切片,每张切片选取10 个清晰视野,分别测量相关指标,取平均值作为该叶片的相关指标数据。
1.2.2 植物叶片横切面解剖结构观察 参考邓立红[22]的超薄切片技术制片,并加以改进,具体步骤如下:①取样,采集无病植物叶片,沿着叶脉纹理用双面刀片切成长条,于载玻片上再分段切成1 mm×2 mm 的小长条,切好后迅速用牙签挑取放入有2.5%的戊二醛溶液的2 mL 离心管中;②抽真空,放入真空仪中抽真空5 min,抽去气泡(空气),使样品完全浸没于2.5%戊二醛溶液中;③前固定,2.5%戊二醛溶液固定72 h;④漂洗,用0.1 mol/L 的PBS 缓冲液(磷酸盐缓冲剂和盐水组成)漂洗3 次,15 min/次;⑤后固定,1.0% 锇酸固定1.5 h;⑥再漂洗,用0.1 mol/L 的PBS 漂洗3 次,15 min/次;⑦梯度脱水,30.0%、50.0%、70.0%、80.0%、90.0%、95.0%、100.0%乙醇溶液各级脱水15 min;⑧脱水,100.0%乙醇溶液二次脱水20 min 后,丙酮脱水20 min;⑨包埋,包埋剂+丙酮(1∶1)室温1 h,包埋剂+丙酮(3∶1)室温3 h;⑩纯包埋过夜,用牙签轻轻地从2 mL 离心管中取出样品放在滤纸上,吸干液体(包埋剂+丙酮),选取2 个切样好的样品用牙签挑起,装入0.5 mL 离心管中(垂直背靠埋在纯包埋剂底部),重复3 次;○1聚合,放入聚合箱中,60 ℃聚合24 h,聚合2 次;○12树脂修块,聚合成型后,取出树脂包埋块,切片机上修去多余树脂,修成梯形,便于进行半薄定位;○13半薄定位,把多余的树脂修掉,露出组织块,便于进行超薄切片,同时,制成半薄切片,用甲苯胺蓝染色后再用倒置荧光显微镜观察叶片横切面解剖结构并拍照;○14超薄切片,将修好的生物样品切片,厚度50~70 nm,用醋酸双氧铀染液和柠檬酸铅染液染色;在透射电子显微镜下观察超微结构并拍照。
2 结果与分析
2.1 阳春砂与草果叶片表皮气孔特征比较
如图1 所示,在观察的阳春砂和草果叶片材料中,上、下表皮均有气孔分布,下表皮气孔分布较多(图1A、图1B),下表皮气孔分布不规则;上表皮气孔分布较少,且上表皮气孔大多分布在叶脉附近(图1C、图1D)。2 种植物的气孔器均由2 个保卫细胞和2 个副卫细胞组成,保卫细胞为肾形保卫细胞,副卫细胞近半圆形(图1E、图1F、图1G、图1H),气孔器微向外凸。

图1 阳春砂与草果叶片上、下表皮气孔分布情况
如表1 所示,阳春砂和草果叶片的气孔存在一定差异,其中阳春砂叶片的上、下表皮气孔数目分别比草果叶片的上、下表皮气孔数目多;草果的上、下表皮气孔长度比阳春砂的上、下表皮气孔长度长;草果的上、下表皮气孔宽度分别比阳春砂的上、下表皮气孔宽度略宽;草果的上表皮气孔开度比阳春砂的上表皮气孔开度略大,草果的下表皮气孔开度较阳春砂的下表皮气孔开度小。
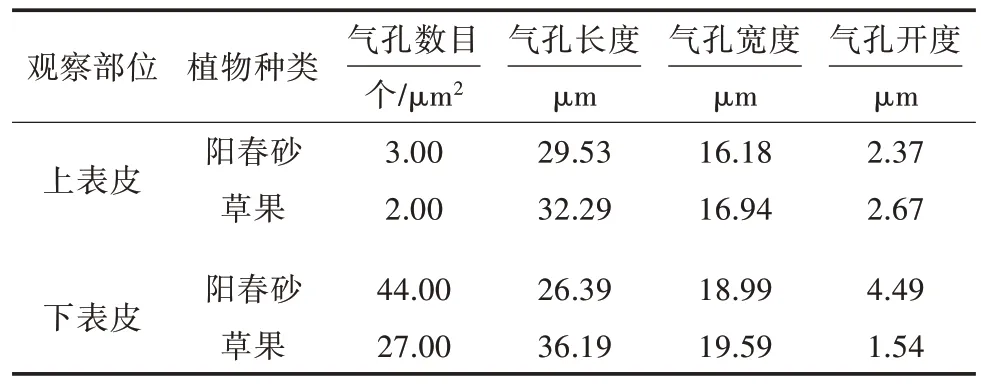
表1 阳春砂与草果叶片的上、下表皮气孔比较
阳春砂的上、下表皮气孔也存在差异,下表皮气孔数目比上表皮气孔数目多13.7 倍,下表皮气孔宽度比上表皮气孔宽度宽17.4%,下表皮气孔开度比上表皮气孔开度大89.5%,下表皮气孔长度比上表皮气孔长度短10.6%。
草果的上、下表皮气孔也存在差异,草果的下表皮气孔数目比上表皮气孔数目多12.5 倍,下表皮气孔长度比上表皮气孔长度长12.1%,下表皮气孔宽度比上表皮气孔宽度宽15.6%,下表皮气孔开度比上表皮气孔开度小42.3%。
2.2 阳春砂与草果叶片横切面解剖结构特征差异比较
如图2A、图2B 所示,从横切面来看,2 种植物均为异面叶,阳春砂与草果的叶片均由表皮、叶肉和叶脉3 个部分组成。表皮分为上表皮和下表皮,表皮由单层表皮细胞组成,分别称为上表皮细胞和下表皮细胞。表皮细胞近方形或不规则形,排列紧密,上表皮细胞较大,下表皮细胞较小。下表皮可观察到气孔器,对应气孔器的位置内侧处有孔下室。叶肉均包括栅栏组织和海绵组织,二者均有一层栅栏组织,海绵组织不规则分布,有2~6 层细胞不等。草果叶片的栅栏组织和海绵组织的分化较明显(图2B),阳春砂叶片的栅栏组织和海绵组织的分化则不太明显,且草果的栅栏组织排列更加整齐、规则。草果的上、下表皮细胞比阳春砂的上、下表皮细胞大。叶脉均由维管束鞘、木质部、韧皮部组成。
2.3 阳春砂与草果叶片电镜超微观结构特征比较
阳春砂与草果的叶片超微观结构如图3A、图3B 所示,其叶肉细胞共有的细胞结构有细胞壁、细胞膜、细胞质、细胞核,原生质体有叶绿体、线粒体。不同的是阳春砂叶片叶肉细胞的叶绿体中含有白色结晶状的淀粉粒、黑色圆点状的嗜锇颗粒,细胞质中含有黑灰色圆点状的内生菌(图3A),而草果叶片叶肉细胞的叶绿体中含有灰色圆点状的脂滴,细胞质中不含内生菌(图3B)。阳春砂与草果叶片下表皮超微结构的气孔由2 个保卫细胞围合而成(图3C、图3D),阳春砂的保卫细胞中含有叶绿体,叶绿体中含有淀粉粒(图3C);草果的保卫细胞中也有叶绿体,但不含淀粉粒(图3D)。
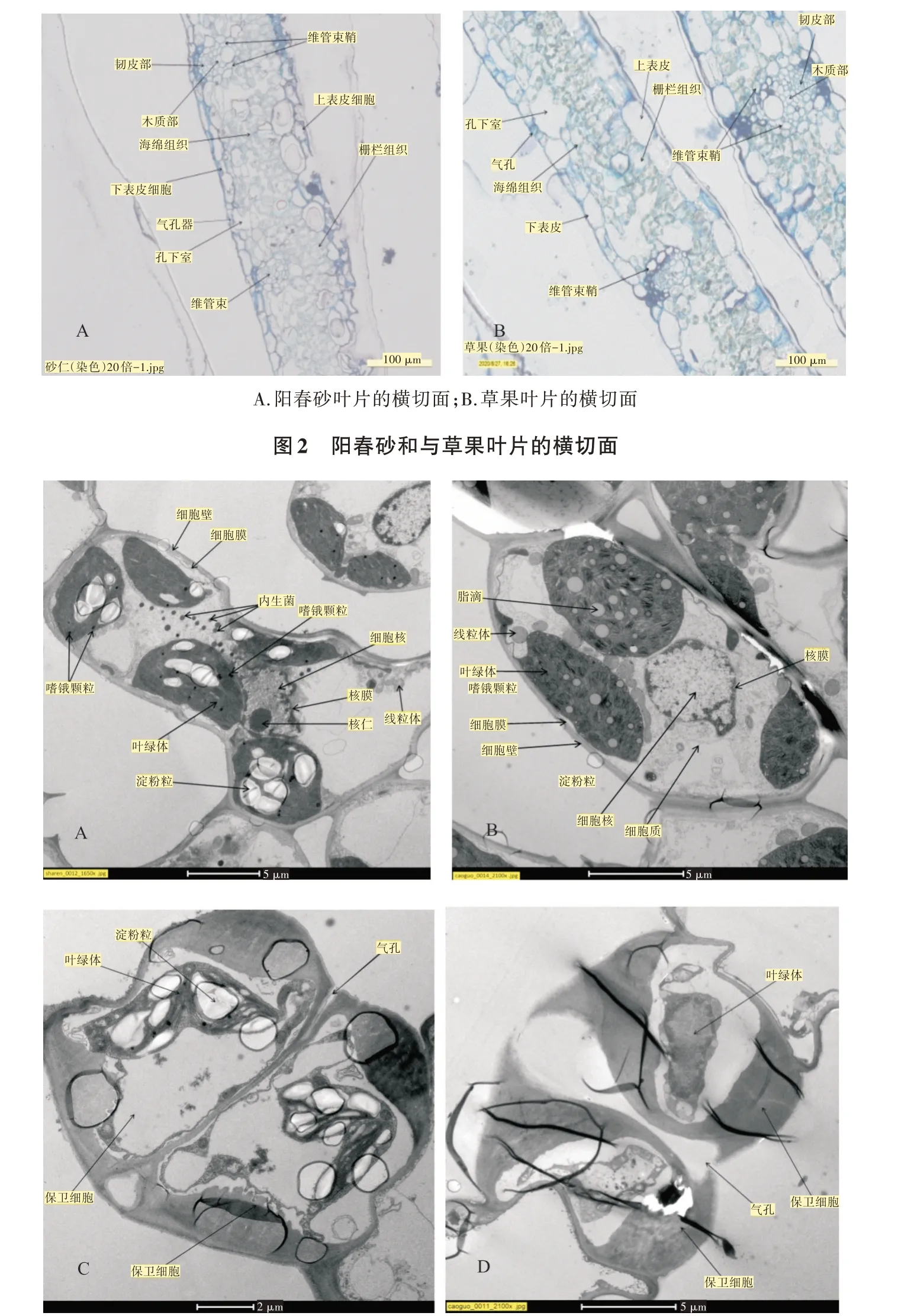
图3 阳春砂与草果叶片横切面超微观结构
3 讨论
3.1 阳春砂和草果叶片气孔特征
本研究观察发现,2 种植物的气孔器均由2 个保卫细胞和2 个副卫细胞组成,保卫细胞均为肾形保卫细胞,副卫细胞近半圆形。邱东萍等[23]、林植芳等[24]研究表明,阳生植物的气孔都分布在下表皮,而阴生植物的上、下表皮都有气孔分布,只是上表皮气孔分布较少,下表皮气孔分布较多。阳春砂和草果2 种植物叶片的气孔上、下表皮均有分布,但大多分布于下表皮,这与黄桂玲[18]对砂仁的研究结果相吻合,与唐源江等[19]对豆蔻属12 个种的叶片横切面解剖结构观察结果一致。阳春砂与草果叶片气孔的分布特点说明二者具备半阴生植物的结构特征。
3.2 阳春砂和草果叶片横切面超微结构特征
近年来,对豆蔻属植物的研究主要集中在叶片表皮表观特征方面[18,19],对叶片横切面结构的研究较为鲜见,本研究主要采用超薄切片法对阳春砂和草果叶片切片后,再用透射电子显微镜对其叶片横切面的解剖结构进行观察分析。吴涛等[25]研究表明发达的海绵组织是叶片对水分充足、弱光环境的一种适应特性,草果和阳春砂栅栏组织不发达,海绵组织较发达可能是对湿热阴蔽的阔叶林下、林边阴湿处条件适应的结果。栅栏细胞是叶片进行光合作用、制造有机物的主要场所,其发达程度与光合强度直接相关,反映植物光合组织的生境适应能力,其厚度影响光合作用中对水分和光能的利用效率[26]。草果的栅栏组织排列整齐,且栅栏组织和海绵组织的分化较明显,而阳春砂的这2 种组织分化不太明显,表明草果的光合作用可能更强一些。
本研究发现,阳春砂与草果叶片下表皮超微结构的气孔由2 个保卫细胞围合而成,气孔器的保卫细胞中有叶绿体,蒋雅婷等[27]也观察到了这一特征。阳春砂和草果的叶肉细胞中叶绿体数目都比较多,但阳春砂的叶绿体内淀粉粒比较大、比较多,这与朱巧玲等[28]观察的半阴生植物石斛的超微结构相似,这可能是阳春砂和草果长期生长于林下阴蔽环境的结果。嗜锇小滴的主要成分是叶绿体内的亲脂性脂类物质,其生理功能可能是提供脂类仓库的作用。姜建福等[29]对葡萄属植物叶片的显微和超微结构研究表明有些葡萄叶绿体中含有较多的嗜锇颗粒,预示着含有较强的抗性。范燕萍等[30]对花叶线艺兰叶片的超微结构研究时在基粒中观察到有较多较大的淀粉粒,在基质中存在较多嗜锇滴。阳春砂叶片叶绿体中有较分散且数量少的嗜锇颗粒,草果中并未发现嗜锇颗粒,却积累了较多的脂滴,表明阳春砂对环境的适应性可能会更强一些,还有待进一步的研究。
观测植物叶片的方法有很多,如印痕法、刮去叶肉法、明胶印迹法、超薄切片法、石蜡切片法等。石蜡切片法和超薄切片法一般适用观察植物组织的横切面,本研究采用超薄切片法,而超薄切片对植物组织的要求较高,阳春砂的叶片质地结构紧密,切成的超薄切片容易产生褶皱,影响观察效果。制作半薄切片时不易得到完整的叶片横切面,且叶片表面革质,用指甲油印痕法制片印痕效果不佳,拍照后图片不够清晰。此外,其他的一些结构特征未能观察到,存在局限性,试验方法有待进一步改进。且超薄切片制成的切片较小只能得到部分横切面,张大鹏等[31]采用冷冻切片法制成的切片观察效果较好,进一步研究时可采用此种方法。
4 小结
研究表明,阳春砂和草果具备半阴生植物的结构特征,体现出对半阴生环境的适应性。2 种植物虽同属,大部分结构相似,但还是存在一定的结构差异,这些差异特征较为稳定且均是2 种植物长期适应林下阴湿环境的结果。本研究从电镜超微观方面观察豆蔻属2 种植物的细胞结构,为该属植物的进一步研究提供了一定的解剖依据,同时为适应温暖湿润、阴蔽的林下生长的植物提供解剖结构依据。
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
热带亚热带植物学报(2021年6期)2021-12-14
食品安全导刊(2021年21期)2021-08-30
建材发展导向(2021年14期)2021-08-23
装备制造技术(2019年12期)2019-12-25
祝您健康·文摘版(2019年4期)2019-06-11
科技创新与品牌(2019年12期)2019-02-06
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中央民族大学学报(自然科学版)(2016年1期)2016-06-27
中国房地产业(2016年9期)2016-03-01
