
狗头枣ACO Ⅰ基因RNAi表达载体的构建及遗传转化
2024-01-08 06:33尚秉众田瑞康雒淑婷王真珍陈国梁
延安大学学报(自然科学版) 2023年4期
尚秉众,田瑞康,雒淑婷,王真珍,陈国梁,2,3*
(1.延安大学 生命科学学院;2.陕西省红枣重点实验室;3.陕西省黄土高原菌产业生态循环发展工程技术研究中心,陕西 延安 716000)
枣树(ZizyphusjujubeMill)是原产于我国的最具特色和优势的果树之一。近年来国内外市场对鲜枣的需求量日渐增加[1],但是鲜枣采摘后易失水、皱缩,果肉酒软、褐变、腐烂,保鲜期极短,大大缩短了鲜枣市场的货架期[2]。乙烯作为一种气态的植物激素,在生理、生化和分子水平上参与果实成熟软化[3-4]。乙烯生物合成途径中有两个关键反应:在第一个反应中,S-腺苷-L-甲硫氨酸(SAM)被ACC-合酶(ACS)转化为1-氨基环丙烷-1-羧酸(ACC);在第二个反应中,ACC 被ACC 氧化酶(ACO)转化为乙烯。其中ACO 是乙烯生物合成中的关键限速酶[5]。研究表明,ACO 基因为多基因家族编码的,在番茄中拥有9 个ACS 和5 个ACO 基因,其中在果实成熟过程中参与乙烯代谢的ACO Ⅰ为主要表达基因[6-8],故采用RNAi 技术对该基因进行特异性沉默来提高其果实的耐储藏性成为可能,而有关以ACO 基因为靶标利用RNAi 技术改变其耐贮性的报道较多。吴晓庆等[9]通过正义,反义ACO 基因转化石竹得到了花期较长的石竹材料;胡春华等[10]通过反义技术抑制香蕉ACO基因,得到了成熟期相比于对照推迟了十天的转基因香蕉;张岩等[11]也通过RNAi 技术对双孢蘑菇的ACO 基因进行干扰,有效的降低了其表达量,并使ACO 酶活力降低至68%以上。目前,RNAi 技术已经应用于番茄、珠子参、柑橘、草莓、马铃薯等[12-16]研究中。但将RNAi 技术应用在枣属植物研究相关基因功能还鲜有报道。基于此,本文选取狗头枣ACO Ⅰ基因保守序列作为靶标构建RNAi表达载体[17],以木枣叶片为外植体对其进行遗传转化,以期获得转基因材料,为培育出耐贮藏的转基因红枣奠定基础。
1 材料与方法
1.1 材料与试剂
ZK品系木枣试管苗,由陕西省红枣重点实验室提供。大肠杆菌DH5α、农杆菌LBA4404、质粒pGM-TR(含基因片段ACO Ⅰ3-4、pART27 与pKANNⅠBAL 均为本课题组保存。实验所需的各种酶均购自TaKaRa公司。
1.2 引物与酶切位点设计
引物与酶切位点组成:
F:5′-GCGGATCCBamHⅠCTCGAGXhoⅠATGAAATT GGAGACACTAGCTGAGG-3′
R:5′-GCATCGATClaⅠGAATTCEcoRⅠAACATTAGT TTCCACTGCTTTC-3′
1.3 RNAi表达载体pART ACO I3-4的构建
采用XhoⅠ/EcoR Ⅰ双酶切pGM-TR(含ACO Ⅰ3-4)及pKANNⅠBAL载体,分别回收靶标片段及线性载体并进行连接,获得pKF1 载体(含正向片段);再将pGM-TR 和pKF1 载体用BamH Ⅰ/ClaⅠ双酶切,分别回收靶标片段及线性载体pKF1 并进行连接,得到中间载体pKF(含正反向片段)。用NotⅠ分别酶切pKF 与pART 27 质粒载体,回收4 200 bp 目的片段并与pART 27线性载体连接,得到了pART ACOⅠ3-4植物表达载体(图1)。
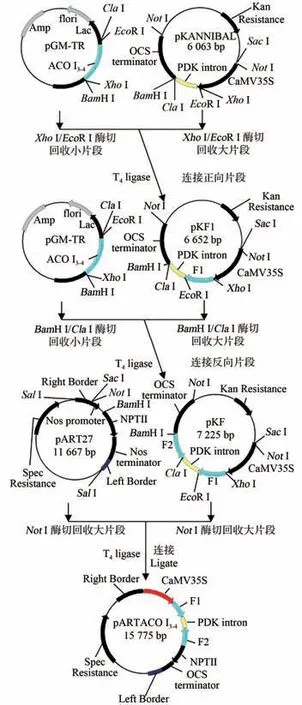
图1 pART ACO Ⅰ3-4载体构建过程
1.4 木枣叶片分化及继代培养Kan选择压筛选
设置了含有5、10、15、20、25、30 mg·L-1Kan 浓度梯度的叶片分化培养基及继代培养基进行Kan选择压的筛选实验。选取生长健壮生长20 d 左右的木枣组培苗将其叶片剪成大小约为0.5 cm2的小块、将茎芽切段分别接种在相应培养基上,观察叶片愈伤组织形成及茎芽继代生长情况并记录,进行Kan选择压的确定。
1.5 木枣遗传转化及筛选鉴定
将pART ACOⅠ3-4表达载体质粒导入农杆菌LBA4404 中获得工程农杆菌,对ZK 品系木枣苗叶片进行转化。在MS固体培养基础上每升添加6-BA 1.0 mg,IBA 0.2 mg,TDZ 0.01 mg 作为基础培养基,在此基础上每升添加AgNO30.5 mg 作为共培养基,在共培养基中添加适量Carb+Kan 进行选择抑菌培养基。将抗性筛选获得的阳性再生木枣植株的叶片作为材料,利用DNA 提取试剂盒提取植物DNA组并进行PCR与电泳鉴定。
2 结果分析
2.1 pART ACO Ⅰ3-4表达载体的成功构建
获得的pKF1 经XhoⅠ/EcoR Ⅰ酶切及PCR 鉴定均获得大小约为600 bp 的片段(图2)与预期大小一致,表明成功获得pKF1。将pKF 用NotⅠ酶切,经电泳检测得到大小约为4 200 bp 和3 100 bp 的片段(图3)与预期结果一致,表明中间载体构建成功。将得到的pART ACO Ⅰ3-4分别用BamH Ⅰ/ClaⅠ双酶切及EcoR Ⅰ单酶切,电泳共检出大小约为1 300 bp,600 bp 和750 bp 的三条带(图4),与预期结果一致;再用Hind Ⅲ、XhoⅠ分别酶切pART ACO Ⅰ3-4,电泳出现约1 000 bp、2 000 bp 大小的片段(图5)。对pART ACO Ⅰ3-4进行PCR 扩增鉴定,结果出现了一条大小约为600 bp 的条带(图6),说明pART ACOⅠ3-4表达载体构建成功。
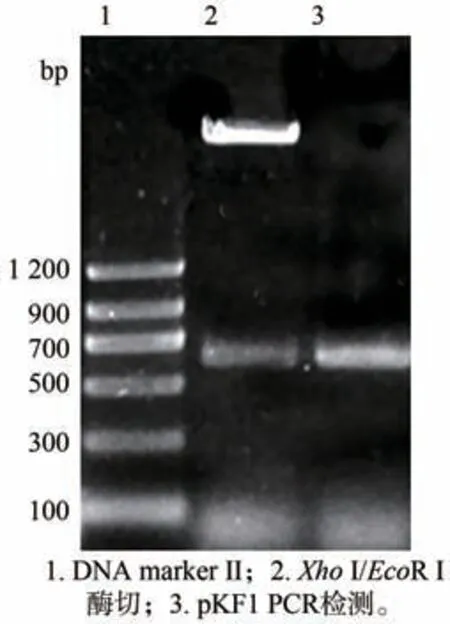
图2 pKF1酶切与PCR鉴定

图3 pKF酶切图
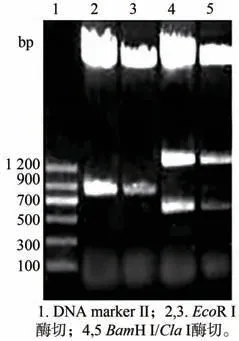
图4 pART ACO Ⅰ3-4的酶切鉴定
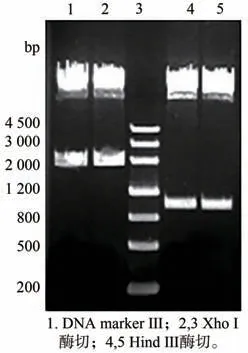
图5 pART ACO Ⅰ3-4的酶切鉴定
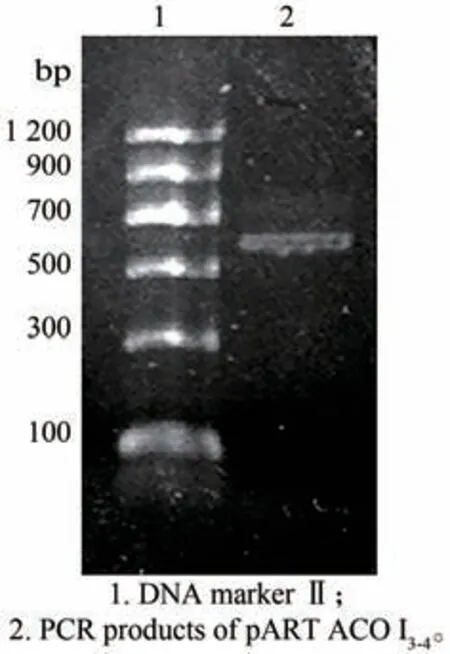
图6 pART ACO Ⅰ3-4的电泳鉴定
2.2 Kan选择压的确定
木枣组培苗在10 mg·L-1Kan 的继代培养基上能够正常生长,随着继代培养基中Kan浓度的升高,木枣组培苗的叶子逐渐变黄,开始出现白化现象,生长态势受阻。直到继代培养中Kan浓度升为30 mg·L-1时,木枣组培苗叶子全部变黄、白化,并且死亡。故将30 mg·L-1Kan 浓度设为木枣遗传转化再生苗继代选择培养基的浓度上限。在Kan 浓度为5 mg·L-1时,叶片出愈率为98.3%,愈伤组织不定芽分化率为61.7%;当Kan 浓度达到10 mg·L-1时,叶片出愈率为37.7%,愈伤组织不定芽分化率11.7%,当Kan 浓度达到15 mg·L-1时,叶片出愈率为6.7%,愈伤组织不定芽1.7%。当Kan浓度达到20 mg·L-1时,愈伤组织趋于死亡,无不定芽分化(图7),所以可将20 mg·L-1Kan浓度设为木枣叶片遗传转化愈伤组织及不定芽形成的Kan标记选择压浓度。
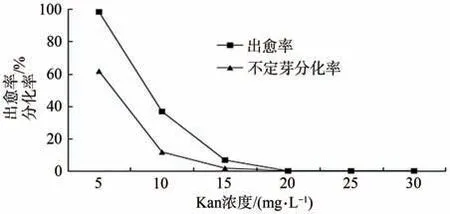
图7 Kan选择压的确定
2.3 转化植株的筛选与PCR鉴定
共侵染木枣叶块600片,共培养之后,放入暗箱进行暗培养3 d,之后移到光下进行培养。4 周后分化出愈伤组织及并逐渐分化成苗(图8A);而对照组叶片没有愈伤组织形成及分化并且开始发黄(图8B)。以分化成苗植株叶片DNA 为模板(图9),经过PCR 与电泳检测鉴定,阴性对照只能扩增出一个条带,大小约为750 bp,然而转化再生植株除可以扩增出两条条带,除了阴性对照组的750 bp 条带外,还能扩增出一条大小约为600 bp 的特异性目的条带(图10),初步证明目的基因转化成功。

图8 木枣愈伤组织及不定芽的形成在分化培养基上

图9 阳性苗在30 mg·L-1 Kan选择压下的生长情况
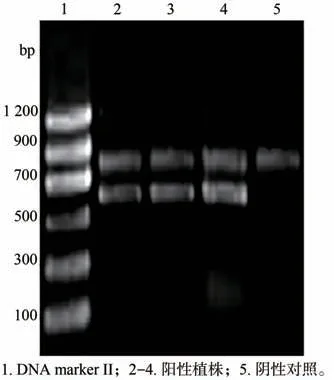
图10 阳性植株的PCR检测
3 讨论与结论
3.1 讨论
鲜枣采收后不易保鲜、不耐贮藏,其果实保鲜期的长短是影响鲜果销售价值的关键品质,是影响其经济价值的重要因素之一,严重制约着红枣产业的发展[18]。目前采用RNAi 技术赋予植物抗性和调控植物代谢等领域已研发出多种具有优良性状的作物,如已成功培育出耐贮藏转基因番茄、桃子、哈密瓜等[19-21],表明通过调控ACO 基因的表达可达到调控乙烯的生成,缓解果实成熟衰老速度,提高果实的耐贮性。人们在枣基因组数据库中发现有ZjACO1、ZjACO2和ZjACO3等3 个ACO 同源基因且其相对表达量与果实成熟度呈显著正相关,其中ZjACO1在枣果实七个不同发育阶段的转录表达模式具有极显著差异[22],因此ZjACO1与果实成熟度相关性最高,而我们沉默的靶标基因正是ZjACO1。
枣品种繁多,呼吸类型复杂,薛梦林等[23]认为枣为非呼吸跃变型果实,由于非跃变型果实成熟衰老过程产生的乙烯远低于跃变型果实,乙烯被认为在非跃变型果实成熟衰老中发挥的作用非常有限。然而,在对柑橘果实的研究表明,低水平的内源乙烯调控了果实的成熟衰老。近年来,随着乙烯参与非跃变型果实成熟衰老相关研究陆续报道,研究人员发现,非跃变型果实诸多成熟相关过程也直接受乙烯调控,且非跃变型果实和跃变型果实品质变化的某些分子调控途径非常相似[24]。越来越多的研究表明,乙烯在非跃变型果实成熟衰老的调控中扮演着重要的角色。如在香蕉、荔枝、柑橘和番茄等[25-28]果实的延迟成熟、延长保质期等方面取得成功。虽然ACO 是催化乙烯生物合成途径的最后关键步骤,并作为靶向基因构建了内含子嵌入的倒置重复序列载体对其进行调控,但干扰效果如何及该基因对枣的成熟和储藏影响如何,还有待于进一步验证。
3.2 结论
成功构建了ihRNAi 植物表达载体,初步筛选到木枣叶片愈伤组织分化培养基,其配方为MS+6-BA 1.0 mg·L-1+IBA 0.2 mg·L-1+TDZ 0.01 mg·L-1,添 加0.5 mg·L-1Carb+20 mg·L-1Kan 可作为筛选培养基。确定了木枣叶片愈伤组织形成Kan 筛选浓度为20 mg·L-1,再生苗的Kan 筛选浓度为30 mg·L-1。采用叶盘法对木枣进行转化,经抗生素筛选和PCR 检测,初步得到了16株阳性木枣转基因植株。
猜你喜欢
数学大王·趣味逻辑(2022年8期)2022-07-10
军民两用技术与产品(2021年10期)2021-03-16
装备制造技术(2019年12期)2019-12-25
世界农药(2019年3期)2019-09-10
天然产物研究与开发(2018年9期)2018-10-08
小资CHIC!ELEGANCE(2018年24期)2018-08-13
上海建材(2017年4期)2017-04-06
现代检验医学杂志(2016年1期)2016-11-12
光学精密工程(2016年4期)2016-11-07
网络空间安全(2016年3期)2016-06-15
