
利用CRISPR/Cas9技术创制水稻Waxy基因等位突变体
2024-01-01 00:00:00朱梦珂王雯雯陈瑜孙宏飞曹黎明林冬枝董彦君
上海师范大学学报·自然科学版 2024年4期







摘""要:"利用CRISPR/Cas9基因编辑技术,在粳稻品种“嘉花1号”的颗粒结合型淀粉合成酶Ⅰ基因(Waxy,Wx)的启动子区域、5'UTR区域,以及第2、3外显子区域设计基因敲除靶点,获得4种不同类型的纯合编辑水稻突变体,随后对其进行稻米品质与农艺性状分析. 结果表明:不同类型Wx纯合编辑突变体与野生型“嘉花1号”相比,核心启动子区域编辑的突变体wxpro直链淀粉含量(AC)下降至14.09%;5'UTR区域编辑的突变体wxutr的AC下降至11.76%,且淀粉粒排列松散;而在外显子区域编辑的突变体wxex2和wxex3的AC下降至2%以下,"与糯稻品种类似. 本研究创制了能稳定遗传且AC不同的纯合突变株系,不仅使水稻Wx基因的等位突变类型变得更加丰富,也为水稻食味品质改良提供了优良的遗传育种材料.
关键词:"水稻;"CRISPR/Cas9;"Wx基因;"直链淀粉含量(AC)
中图分类号:"Q 344 """文献标志码:"A ""nbsp;文章编号:"1000-5137(2024)04-0502-10
Creation of Waxy"allelic mutants of the rice Waxy gene"using CRISPR/Cas9 technology
ZHU Mengke WANG Wenwen CHEN Yu SUN Hongfei "CAO Liming LIN Dongzhi DONG Yanjun
(1.College of Life Sciences,"Shanghai Normal University,"Shanghai 200234,"China;"2.Crop Breeding and Cultivation Research Institute,"Shanghai Academy of Agricultural Sciences,"Shanghai 201403,"China)
Abstract:"In this study,"CRISPR/Cas9 gene editing technology was used to design four knockout targets located in the promoter region,"5'UTR region and exons 2 and 3 of the granule bound starch synthase gene (Waxy,"Wx)"in the japonica"rice variety “Jiahua 1”. We then obtained four different types of T1"generation homozygous edited-mutants without transgene to investigate their quality and agronomic traits. Compared to the wild type “Jiahua 1”,"the amylose content (AC)"of the wxpro"mutants in the promoter region decreased to 14.09%,"while the wxutr mutants"in the 5'UTR region decreased to 11.76%,"with a loose arrangement of starch grains. The wxex2"and wxex3"mutants in the exon regions had an AC of to less than 2%,"similar to that of glutinous rice. In summary,"all stable-inherited homozygous wx mutants,"with varying AC,"not only enrich the Wx allelic mutant genes in rice,"but also provide valuable genetic and breeding materials for improving rice eating quality.
Key words:"rice;"CRISPR/Cas9;"Wx"gene;"amylose content (AC)
0 "引"言
水稻是我国主要的粮食作物,随着社会经济的发展和国民生活水平的提高,人们对稻米蒸煮食味品质的要求愈来愈高,食味品质改良受到了水稻育种家们的高度关注. 科学研究已经证实,影响稻米蒸煮食味品质的3个主要评价指标是:胶稠度、糊化温度和直链淀粉含量(AC),其中AC被认为是主要指标[1-2]. AC通常与稻米食味品质成反比,它决定着水稻胚乳的外观和适口性,但是AC并非越低越好[3]. 低AC的水稻胚乳呈现乳白色,煮熟后米饭柔软且有光泽,黏度大、适口性好. AC主要由编码颗粒结合淀粉酶I(GBSSI)的Wx基因调控[4]. 水稻不同品种间AC的差异主要是由Wx位点的变异引起的[5]. 目前,已被发现和克隆的Wx复等位基因(如Wxa,Wxb,Wxlv,Wxin,Wxop/hp,Wxmq,Wxmp,Wxmv和wx等)均已应用于水稻育种[6-9]. 进一步发掘和创制新的Wx复等位基因将有助于水稻新品种的开发和稻米食味品质的改良[10].
近年来,CRISPR/Cas9基因编辑技术由于其操作简单高效,而且靶向性强等优点,已被广泛应用于水稻性状的遗传改良[11]. ZHANG等[12-13]通过分别编辑CSA和TMS5基因,开发了用于两系法杂交水稻的水稻光温敏雄性不育系. 本实验室也成功对水稻谷蛋白基因GluA3和丙酮酸磷酸双激酶(PPDK)基因进行编辑,分别获得了谷蛋白含量降低的突变体[14]以及粉质突变体[15]. 值得一提的是,ZENG等[16]也成功利用CRISPR/ Cas9技术对Wx基因的核心启动子区域和5'UTR区域进行编辑,"获得具有不同AC的水稻突变体. 由此说明,可采用对基因不同位点编辑的方式对Wx基因的表达进行精准调控,可产生新的Wx复等位基因,创制不同AC的突变体.
本研究选取江浙沪主栽水稻品种“嘉花1号”作为研究材料,在Wx基因的编码区和非编码区位置设计不同靶位点,以创制新的Wx基因等位突变体,为水稻食味品质改良提供良好的材料.
1 "材料与方法
1.1 实验材料
本实验选取江浙沪主栽水稻品种“嘉花1号”作为受体材料,江苏省农科院选育的“太湖糯2号”作为糯稻对照品种. 实验材料均种植于上海师范大学遗传研究所的封闭水稻基地,采用常规水肥管理.
1.2 CRISPR/Cas9敲除表达载体的构建和水稻的遗传转化
本实验使用的CRISPR/Cas9载体由华南农业大学刘耀光院士提供. 利用CRISPR Primer Designer靶点引物设计软件分别在Wx基因(LOC_Os06g04200)启动子,5'UTR,第2、3外显子(Ex2,"Ex3)区域内各设计2个长度为19~20 bp的靶位点(表1). 参照曾栋昌等[17]的方法,在每个区域构建1个双靶点表达载体,共4个表达载体.参照孙佳等[15]的方法,进行水稻的遗传转化和组织培养实验,最后成功获得水稻转基因T0代幼苗.
1.3 编辑植株的鉴定及靶位点检测
本实验采用简易快捷DNA提取(TPS)法[18]提取水稻叶片DNA. 由于构建的表达载体上含有潮霉素抗性(HPT)基因,利用HPT基因的引物(表1)对T0代幼苗进行检测. 确定为转基因阳性幼苗后,选取不同靶位点位置上下各约200 bp,设计靶点特异性引物(表1),DNA聚合酶链反应(PCR)扩增得到大小约600 bp的DNA片段,测序后与“嘉花1号”序列比对,确定突变类型. 然后,利用载体特异性引物HPT- F/HPT-R(表1)对加代获得的T1代植株进行扩增,鉴别和区分是否携带HPT基因的T1代编辑植株.
1.4 qRT-PCR分析
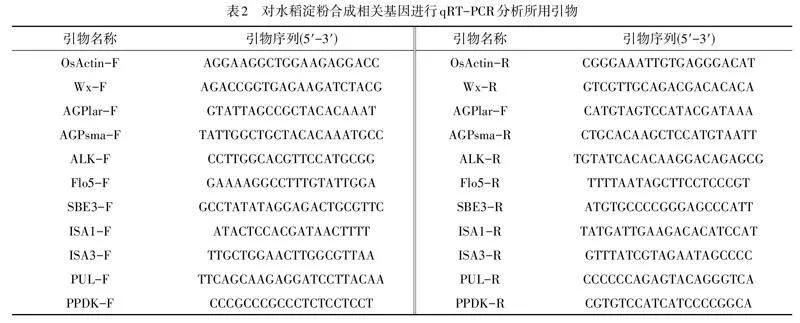
提取突变体、“嘉花1号”以及“太湖糯2号”开花后第5,10,15,20,25,30,35天的籽粒总RNA,并反转为cDNA(RNA提取试和反转录试剂盒同文献[14]). 以OsActin(LOC_Os03g50885)作为内参基因,利用实时定量反转录PCR(qRT-PCR)对与水稻淀粉合成相关的10个基因(表2)进行转录水平的表达分析,实验重复3次.
1.5 淀粉粒扫描电子显微镜(SEM)观察
参照孙佳等[15]的方法对编辑突变植株和对照品种水稻植株成熟后的籽粒的淀粉粒结构进行SEM观察,并拍照记录.
1.6 淀粉含量测定
收获的水稻种子干燥后,脱壳保存. 参照孙佳等[15]的方法,利用碘染色法进行表观直链淀粉的检测分析,并拍照记录. 取适量糙米处理成粉末状,过100目(孔径150 μm)筛后保存留用. 每份样品称取20 mg,使用Megazyme公司(爱尔兰)淀粉试剂盒,测定其直链淀粉以及总淀粉含量,实验重复3次.
1.7 蛋白含量测定
稻谷成熟干燥后,脱壳保存. 利用FOSS近红外谷物品质分析仪(Infratec TM1241)测定其粗蛋白含量,实验重复3次.
1.8 米粒外观与农艺性状调查
水稻材料均种植于同一实验田,水肥等栽培条件相同. 待水稻植株成熟后,调查各类型水稻材料的各项农艺性状指标,观察植株形态和糙米外观,并拍照记录,实验均重复3次.
2 "结果与分析
2.1 CRISPR/Cas9敲除表达载体的构建与编辑突变水稻的获得
根据曾栋昌等[17]的方法构建敲除表达载体,成功构建了在Wx基因的启动子、5'UTR、第2外显子以及第3外显子区域内的4个表达载体(图1). 分别侵染野生型水稻“嘉花1号”的愈伤组织,经组织培养过程,获得转基因T0代幼苗后再利用引物HPT-F/R进行检测,共获得携带HPT基因的转基因T0代植株41株. 其中,在核心启动子区域编辑的突变体8株(7株为纯合型);在5'UTR区域发生编辑的突变体7株(均为纯合型);在第2外显子区域编辑的突变体"21株(20株为纯合型);在第3外显子区域发生编辑的突变体5株(3株为纯合型). 进一步利用载体特异性引物"HPT-F/R扩增T1代突变株来剔除携带HPT基因的植株,获得了不含HPT基因,且在启动子,5'UTR,第2、3外显子区域内编辑的纯合突变体,分别命名为wxpro,wxutr,wxex2和wxex3,与野生型“嘉花1号”的序列比对发现,它们分别发生了5对碱基替换,以及3个对碱基、282个对碱基与27对碱基缺失(图2).
2.2 编辑突变体中淀粉合成相关基因RNA表达水平分析
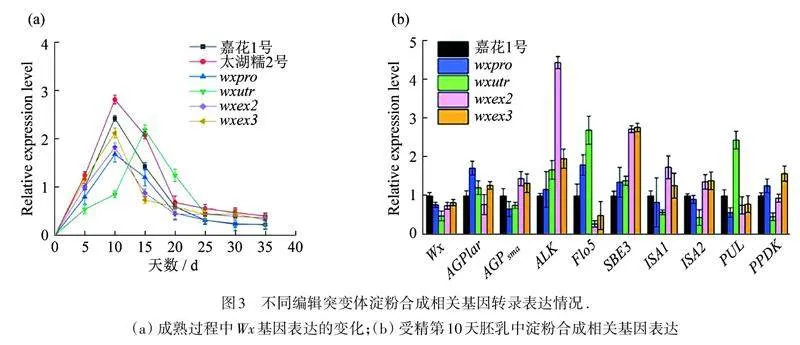
为了明确编辑突变体(wxpro,wxutr,wxex2,wxex3)灌浆期籽粒中的Wx基因表达是否受到影响,对开花授精后第5,10,15,20,25,30,35天的籽粒中Wx基因进行转录水平分析,结果如图3(a)所示,Wx基因的高峰表达期为授精后10~15 d,除了wxutr峰值出现在第15天以外,其余峰值均在第10天,由此推测5'UTR端突变会导致Wx基因表达推迟. 更有意思的是,出现表达量最大的2个品种恰恰是未被编辑过的“嘉花1号”和“太湖糯2号”,这也说明编辑突变体中Wx的转录表达或多或少都受到抑制,其中,启动子编辑的突变体(wxpro)影响最明显.
进一步对授精第10天的胚乳进行淀粉合成相关基因的转录水平分析,如图3(b)所示,发现在启动子编辑的wxpro突变体中AGPlar(腺苷二磷酸葡萄糖焦磷酸化酶大亚基编码基因)和Flo5(可溶性淀粉合酶Ⅲ编码基因)出现了明显上调,而PUL(异淀粉酶编码基因)下调. 在5'UTR端编辑的wxutr植中,ALK(淀粉合酶Ⅱ编码基因)、Flo5(可溶性淀粉合酶Ⅲ编码基因)和PUL(异淀粉酶编码基因)出现了不同程度的上调,而ISA1(淀粉脱分支酶1编码基因)、ISA2(淀粉脱分支酶2编码基因)和PPDK(丙酮酸磷酸双激酶编码基因)表达下调. 在第2、3外显子区域编辑的突变株(wxex2和wxex3),对淀粉合成相关基因的表达水平的影响几乎完全一致,ALK和SBE3(淀粉分支酶Ⅱb编码基因)表达明显上调,而Flo5则出现了明显的下调. 由此可知,Wx基因的突变会影响与淀粉合成相关基因(包含自身Wx基因)的转录表达.
2.3 淀粉颗粒显微结构变化
对野生型“嘉花1号”"“太湖糯2号”以及编辑突变体的稻米横切面进行SEM观察. 结果如图4所示:野生型“嘉花1号”和“太湖糯2号”稻米的淀粉粒均呈现不规则的多边形状,淀粉粒排列紧凑,边缘棱角分明;wxpro突变体的淀粉粒稍显疏松,也呈现不规则多边形状,有棱角,出现少量圆形淀粉颗粒;wxutr突变体的淀粉粒为松散的圆形状颗粒;wxex2和wxex3突变体的淀粉粒结构与野生型“嘉花1号”和“太湖糯2号”相似,呈现不规则多边形,排列紧密.
2.4 淀粉与蛋白含量的变化
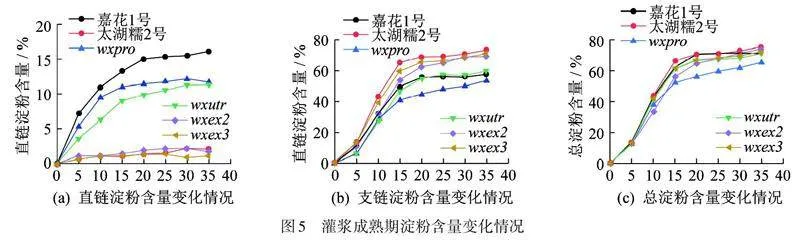
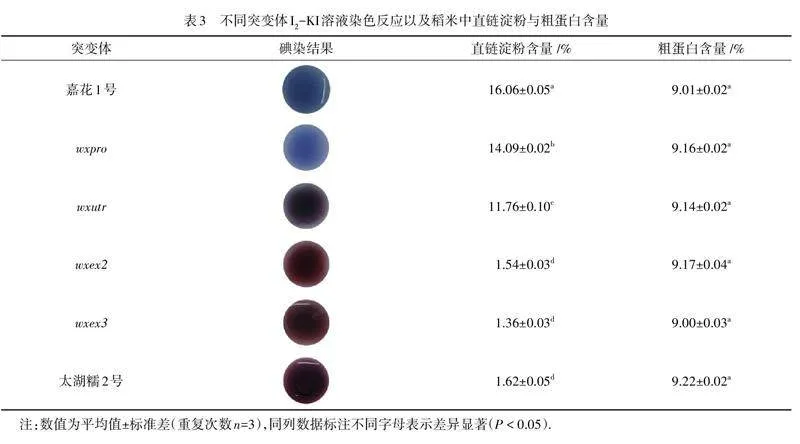
对水稻灌浆期各阶段的淀粉含量进行测定分析,结果如图5所示:所有测试材料的直链淀粉(图5(a)),支链淀粉(图5(b)),以及总淀粉含量(图5(c)),都随着生育期的持续而升高,在成熟期的第35天到达峰值. 值得一提的是,受精后第15~20 天,它们的含量也基本接近峰值,与转录表达基本相一致,如图3(a)所示. 成熟后(表3),wxpro (14.09%)和"wxutr(11.76%)籽粒中的AC均明显低于“嘉花1号”(16.06%);与糯稻品种“太湖糯2号(1.62%)”一样,wxex2(1.54%)和wxex3(1.36%)米粒中的直链淀粉合成几乎完全被阻止. 此外,I2-KI溶液染色反应结果与AC从高到低的结果相一致(表3). 此外,4种编辑突变体中的粗蛋白含量(9.00%~9.67%)与“嘉花1号”(9.01%)相比变化不大. 由此可见,Wx基因的突变对淀粉合成的影响较大,且影响程度与突变位点相关.
2.5 米粒外观与农艺性状
水稻成熟后,对糙米外观及农艺性状进行调查,结果如图6和表4所示. “嘉花1号”是典型的粳稻特征,呈现出短圆粒,半透明胚乳;“太湖糯2号”为白色不透明胚乳,是典型的糯稻特征;wxpro外观与“嘉花1号”相似,呈半透明胚乳;wxutr的胚乳出现了不同程度的白色不透明状;wxex2和wxex3的胚乳完全为白色不透明状,与“太湖糯2号”一致. 和野生型嘉花1号相比,除了wxutr之外,剩余编辑突变体的株高普遍有所下降;穗长除wxpro外,均变短;每株穗数除wxex3外,均变少;而结实率均降低,wxex3下降幅度最大;千粒重除wxutr增加之外,其他类型均出现下降趋势. 由此可知,Wx基因的突变不仅会改变米粒外观,也会影响一些农艺性状.
3 "讨"论
稻米中的淀粉是人们所需碳水化合物的主要来源,可分为直链淀粉和支链淀粉两种. 其中,AC高低与米饭的色泽、硬度、弹性和黏度等特性相关[19]. CRISPR/Cas9技术因其成本低、操作简便、效率高等优点,已广泛应用于生命科学领域[20],目前已有许多利用CRISPR/Cas9技术对水稻品质性状进行改良的成功案例[12-15]. 本研究利用CRISPR/Cas9技术对“嘉花1号”Wx基因编码区的2个外显子区域和基因调控区域(启动子与5'UTR端)分别进行编辑,成功获得了4种在启动子(wxpro),5'UTR(wxutr)"以及第2(wxex2)、第3(wxex3)外显子区域编辑且对淀粉品质有影响的水稻突变植株.
3.1 启动子与5' UTR的编辑改变对Wx基因的表达有调控作用
本研究获得了在Wx基因核心启动子区域编辑的突变体wxpro,其籽粒外形与农艺性状没有发生明显变化,但造成了淀粉粒的松散排列,总淀粉含量和直链淀粉含量下降,稻米品质发生变化. 虽然wxpro的Wx基因在灌浆成熟期的表达模式与野生型一致,但表达量有所下调. 然而,与支链淀粉合成相关的AGPlar基因表达量出现明显上调. 推测原因是:(1)"核心启动子区域的变化影响了与转录因子的结合,使得形成的转录复合体结构不稳定,GBSSⅠ蛋白表达下调,导致AC下降;(2)"由于基因反馈作用,直链淀粉合成下降,促使支链淀粉合成增加. HUANG等[21]也对Wx基因启动子TATA盒区域进行编辑,获得了6种AC微调的突变体类型,与本结果相类似. 同样,在5'UTR端编辑的突变体wxutr,其胚乳出现了不同程度的白色不透明状态,粒形也变得更加圆润. 穗长和结实率与野生型无明显差异,千粒重明显增加,AC降至11.76%,其Wx基因表达量在整个灌浆成熟期明显下降,峰值推迟了5 d;而与支链淀粉合成相关的Flo5,PUL等基因却出现明显的上调. 推测5'UTR区域结构的变化会影响前体mRNA的剪接,使得mRNA的稳定性下降,导致翻译的GBSSⅠ 蛋白减少,最终使得直链淀粉的合成量下降. 由于直链淀粉的合成受阻,而异淀粉酶合成相关基因的表达量增加,淀粉合成向异淀粉粒方向偏移,导致形成的淀粉粒出现异常,淀粉粒排列松散且有不同程度的白色不透明胚乳出现. ZENG等[16]也利用CRISPR/Cas9技术对Wx基因的5'UTR区域进行定向编辑,获得白色不透明状胚乳且AC下降的突变体,与本研究结果相吻合. 因此,认为非编码区的启动子与5'UTR的编辑突变对Wx基因的表达有调控作用,可以改变其胚乳合成直链淀粉的能力.
3.2 Wx基因编码区的编辑使颗粒结合淀粉酶I(GBSSI)失活
本研究中获得了在Wx基因编码区的第2(wxex2)、第3(wxex3)外显子的纯合编辑且淀粉品质改变的2种水稻突变体,这2种突变均造成Wx基因编码的蛋白质翻译提前终止,三级结构变得简单、疏松(图7). 从籽粒外观分析,两类突变体都变得更加细长,胚乳为不透明状,与糯稻“太湖糯2号”一致,其产量也出现了下降趋势,因此可以判断,编码区的变化对淀粉品质和农艺性状的影响较大. wxex2"和wxex3的淀粉粒结构均与“嘉花1号”类似,排列紧密,呈现不规则多边形状. 虽然Wx基因在灌浆期的表达模式与糯稻“太湖糯2号”和“嘉花1号”相一致,但表达量出现了微量下调. 籽粒中的总淀粉含量无明显差异,但其AC却出现了大幅度下调情况,仅有1.54%和1.36%,比糯稻品种“太湖糯2号”还低,可能是一种新型糯稻类型. 虽然Wx基因在wxex2和wxex3中的表达量仅有微量下调,但与支链淀粉合成相关的ALK和SBE3基因却出现了明显上调. 因此可以推测,虽然Wx基因的转录表达水平正常,但由于翻译提前终止,使得GBSSⅠ蛋白功能结构域缺失,以致直链淀粉的合成受阻,进而转向合成支链淀粉,最终出现典型糯稻的表型特征.
4 "结"论
本研究共获得4种靶向Wx基因不同位置编辑的突变植株,通过对其农艺性状、淀粉品质性状和转录水平分析发现:无论是编码区还是启动子与5'UTR这类非编码区的变化都会影响Wx基因的表达,使AC出现不同程度的下调. 启动子与5'UTR这类非编码区突变后影响较小,能微量调节其直链淀粉含量变化,而编码区的变化对其影响显著,出现直链淀粉合成受阻、AC大幅下调的情况. 本研究的结果不仅为水稻食味品质改良提供新的突变体材料,也再次验证了利用CRISPR/Cas9技术靶向Wx基因不同位置对AC进行精准调节的现实可行性,为水稻品质改良育种提供更加广阔的思路.
参考文献:
[1] TIAN Z X,"QIAN Q,"LIU Q Q,"et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities [J]. Proceedings of the National Academy of Sciences,"2009,106(51):21760-21765.
[2] BISELLI C,"CAVALLUZZO D,"PERRINI R,"et al. Improvement of marker-based predictability of apparent amylose content in japonica rice through GBSSI allele mining [J]. Rice,"2014,7(1):1.
[3] HIGGINS J A. Resistant starch:"metabolic effects and potential health benefits [J]. Journal of AOAC International,"2004,87(3):761-768.
[4] TENG B,"ZENG R Z,"WANG Y C,"et al. Detection of allelic variation at the Wx"locus with single-segment substitution lines in rice (Oryza sativa"L.)"[J]. Molecular Breeding,"2012,30(1):583-595.
[5] ZHANG C Q,"YANG Y,"CHEN S J,"et al. A rare Waxy"allele coordinately improves rice eating and cooking quality and grain transparency [J]. Journal of Integrative Plant Biology,"2021,63(5):889-901.
[6] ZHANG C Q,"ZHU J H,"CHEN S J,"et al. Wxlv,"the ancestral allele of rice Waxy"gene [J]. Molecular Plant,"2019,12(8):1157-1166.
[7] LIU Y,"ZHANG A N,"WANG F M,"et al. Development and validation of a PCR-based functional marker system for identifying the low amylose content-associated gene Wxhp in rice [J]. Breeding Science,"2019,69(4):702-706.
[8] SATO H,"SUZUKI Y,"SAKAI M,"et al. Molecular characterization of Wx-mq,"a novel mutant gene for low-amylose content in endosperm of rice (Oryza sativa L.)"[J]. Breeding Science,"2002,52(2):131-135.
[9] YAO S,"ZHANG Y D,"LIU Y Q,"et al. Effects of soluble starch synthase genes on eating and cooking quality in semi waxy japonica rice with Wxmp"[J]. Food Production,"Processing and Nutrition,"2020,2(1):22.
[10] HUANG L C,"SREENIVASULU N,"LIU Q Q. Waxy"editing:"old meets new [J]. Trends in Plant Science,"2020,25(10):963-966.
[11] MA X L,"ZHANG Q Y,"ZHU Q L,"et al. A robust CRISPR/Cas9 system for convenient,"high-efficiency multiplex genome editing in monocot and dicot plants [J]. Molecular Plant,"2015,8(8):1274-1284.
[12] ZHANG H,"XU C X,"ZONG J,"et al. Mutation in CSA"creates a new photoperiod-sensitive genie male sterile line applicable for hybrid rice seed production [J]. Proceedings of the National Academy of Sciences of the United States of America,"2013,110(1):76-81.
[13] BARMAN H N,"SHENG Z H,"FIAZ S,"et al. Generation of a new thermo-sensitive genic male sterile rice line by targeted mutagenesis of TMS5"gene through CRISPR/Cas9 system [J]. BMC Plant Biology,"2019,19(1):109.
[14] 周优,"林冬枝,"董彦君. CRISPR/Cas9技术定点编辑水稻谷蛋白基因GluA3"[J]. 上海农业学报,"2019,35(1):22-28.
ZHOU Y,LIN D Z,"DONG Y J. Targeted editing of rice gluten gene GluA3 by CRISPR/Cas9 technology [J]. Acta Agriculturae Shanghai,"2019,35(1):22-28.
[15] 孙佳,"吴清清,"周文昊,"等. 利用CRISPR/Cas9技术编辑水稻丙酮酸磷酸双激酶基因PPDK"[J]. 上海农业学报,"2021,37(3):1-6.
SUN J,"WU Q Q,"ZHOU W H,"et al. Editing rice pyruvate phosphodouble kinase gene(PPDK)by CRISPR/Cas9 technology [J]. Acta Agriculturae Shanghai,"2021,37(3):1-6.
[16] ZENG D C,"LIU T L,"MA X L,"et al. Quantitative regulation of Waxy"expression by CRISPR/Cas9‐based promoter and 5'UTR-intron editing improves grain quality in rice [J]. Plant Biotechnology Journal,"2020,18(12):2385-2387.
[17] 曾栋昌,"马兴亮,"谢先荣,"等. 植物CRISPR/Cas9多基因编辑载体构建和突变分析的操作方法"[J].中国科学:生命科学,"2018,48(7):783-794.
ZENG D C,"MA X L,"XIE X R,"Construction and analysis of plant CRISPR/Cas9 multi gene editing vector [J]. Scientia Sinica (Vitae),"2018,48(7):783-794.
[18] 张向前,"邹金松,"朱海涛,"等. 水稻早熟多子房突变体font5的遗传分析和基因定位"[J]. 遗传,"2008,30(10):1349-1355.
ZHANG X Q,"ZOU J S,"ZHU H T,"et al. Genetic analysis and gene mapping of an early flowering and multi-ovary mutant in rice (Oryza sativa"L.)"[J]. Hereditas,"2008,30(10):1349-1355.
[19] ZHANG L,"ZHAO L L,nbsp;ZHANG J,"et al. Relationships between transparency,"amylose content,"starch cavity,"and moisture of brown rice kernels [J]. Journal of Cereal Science,"2019,90:102854.
[20] MA Z F,"WEN M Y,"ZHANG Y X,"et al. Development of CRISPR/Cas9 genome editing system and its application in rice molecular breeding [J]. In Vitro Cellular amp; Developmental Biology-Plant,"2021,57:700-708.
[21] HUANG L C,"LI Q F,"ZHANG C Q,"et al. Creating novel Wx"alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system [J]. Plant Biotechnology Journal,"2020,18(11):2164-2166.
(责任编辑:顾浩然)
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29 01:57:52
中国生殖健康(2020年4期)2021-01-18 02:58:10
中国粮油学报(2019年4期)2019-07-12 09:06:32
中国生殖健康(2018年4期)2018-11-06 07:12:16
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
中国塑料(2016年2期)2016-06-15 20:29:57
山东医药(2015年40期)2015-02-28 14:28:45
湖北农业科学(2014年11期)2014-09-10 18:06:07
食品科学(2013年23期)2013-03-11 18:30:11
