
猪精液冻融技术及其影响因素研究进展
2024-01-01 00:00:00范振港刘鑫苗义良张霞
华中农业大学学报 2024年5期

关键词 猪; 精液冻融; 冷冻保护剂; 冻精解冻剂; 精液理化性质
猪精液冷冻是指以液氮为冷冻介质将精液保存于超低温环境(-196 ℃)下,进而抑制精子的生理代谢活动,实现对精液长期保存的一种方法。精液冷冻流程主要包括精液采集、冻前处理、冻前降温平衡、添加冷冻保护剂、程序性冷冻,精液解冻流程主要包括加热解冻、添加解冻稀释液、解冻后精液质量评估等。虽然猪精液冻融技术存在解冻后精子活力低的问题,但若该技术能在生产上规模应用,不仅可提高优秀种公猪利用率,而且可降低生物安全风险,提高生猪的生产效率。因此,本文针对目前猪精液冻融技术进行综述,旨在为从事猪精液冻融技术研发的科研人员了解该方向研究的最新动态提供参考,进而为猪精液冻融技术的完善提高并实现在养猪生产上的规模应用提供科学依据。
1 猪冷冻精液发展概况及主要技术流程
人工授精技术变革了猪的繁殖方式并促进了生猪养殖业的发展,如今93% 以上的生猪生产都采用人工授精技术。猪精液的冷冻研究始于20 世纪50年代,国内外研究人员主要对冷冻稀释液、冷冻保护剂、平衡时间、冷冻剂型、冷冻方法等开展了大量研究。1970 年,Polge 等 [1]在英国首次成功使用冻融后的精液进行人工授精并获得健康仔猪。1973 年,Sal⁃amon 等[2]采用颗粒冻精进行人工授精,成功使5 头母猪妊娠,产仔数平均为7.6 头。20 世纪80 年代后,猪冷冻精液的主流剂型和方法由颗粒形式、干冰制备逐渐改为细管形式、液氮熏蒸。我国广西地区在20 世纪80 年代使用冻融精液进行人工授精配种母猪头数达6 万头以上, 受胎率接近76%,平均窝产仔数达9 头以上[3]。日本在1991 年使用冻融精液进行人工授精,配种分娩率为72.3%,平均窝产仔数8.0头[4]。虽然猪精液冷冻技术经过长期的研究已经取得了很大进展,但仍存在冻融后的精子损伤大、活力低及受精能力弱等问题,导致该技术的使用率较低[5]。
猪精液冷冻的流程包括精液的采集、冻前处理、冻前降温平衡、添加冷冻保护剂和冷冻处理。冷冻处理的方式有缓慢冷冻和快速冷冻(玻璃化冷冻)。缓慢冷冻方法包括干冰冷冻、液氮熏蒸和程序冷冻仪冷冻。缓慢冷冻过程中,精子在含有冷冻保护剂的稀释液中,通过逐步降温的方法进行冷冻。快速冷冻是使用高浓度的冷冻保护剂,让精子和冷冻溶液在冷冻时呈现黏稠而不产生结晶的玻璃化状态。解冻过程主要分为低温慢速解冻和高温快速解冻。在解冻剂中可添加促精子复苏物质、抗损伤物质来提高解冻后精子的活力。目前解冻程序主要有低温解冻(0~5 ℃)、中温解冻(30~40 ℃)、高温解冻(50~70 ℃),解冻时间有20、16 和8 s 等。
在猪精液预处理降温过程中,不同猪个体的精子对“低温打击”敏感度存在明显差异,这反映了不同猪个体的精子耐冻性(freezability)存在差别。根据冻精解冻后质量评定的高低可将精液来源分为好(good freezers)和差(bad freezers)2 种,简称为GFE(good freezability ejaculates)和PFE(poor freezabilityejaculates)[6],或GSF(good semen freezability)和PSF(poor semen freezability)[7]。造成精液来源存在质量差异的机制尚不清楚,推测遗传差异可能是其耐冻性差异的基础。
2 影响猪精液冻融后精子活力的关键因素
2.1 猪精液冷冻稀释液和冷冻保护剂
常见的猪精液冷冻稀释液有2 种,一种是乳糖-蛋黄体系(lactose egg yolk, LEY),一种是贝尔茨维尔体系(beltsville freezing extender 5, BF5)[8],这2 种稀释液均需添加新鲜蛋黄。精液在稀释时有一步稀释法和两步稀释法,一步稀释法是直接将精液与含有冷冻保护剂的稀释液按比例进行混合,两步稀释法是先将精液与不含冷冻保护剂的稀释Ⅰ液按比例混合,后经降温平衡再与含冷冻保护剂的稀释Ⅱ液进行混合,再经降温平衡后进行冷冻。
在两步稀释法中,稀释Ⅱ液通常添加冷冻保护剂(cryoprotectant agents, CPA),CPA 分为渗透性冷冻保护剂和非渗透性冷冻保护剂。渗透性冷冻保护剂最常用的是甘油,此外还有二甲基亚砜、乙二醇、甲醇、丙二醇和二甲基乙酰胺等,该类物质通过与氢键相互作用高度溶于水并能透过质膜。甘油在高于5 ℃时会干扰细胞的代谢,因此在使用甘油时需在低温条件下将甘油与精液混合。研究表明甘油添加量为2% 和4% 时会破坏猪精子的核膜[9],高于4% 时会影响质膜流动性,高于6% 则会影响猪精子的运动性和顶体完整性[10],因此,目前精液冷冻过程中甘油添加量一般以3% 为宜。
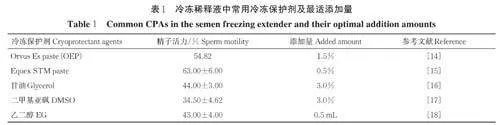
非渗透性冷冻保护剂最常用的是新鲜蛋黄,此外还有牛奶、羟乙基等聚合物,如右旋糖酐、聚乙二醇和聚乙烯吡咯烷酮等。相较于甘油,蛋黄物质起到的冷冻保护效果更佳,但蛋黄物质对蛋黄的新鲜程度有很高要求,且其生物安全性很难得到保障。目前可使用从蛋黄中提取的低密度脂蛋白(low den⁃sity lipoprotein, LDL)代替蛋黄作为冷冻保护剂,研究表明,添加0.09 g/mL LDL 可具有很好的精液冷冻保护效果[11]。大豆卵磷脂同样富含LDL,有研究证实大豆卵磷脂可替代蛋黄作为冷冻保护剂,当添加5% 大豆卵磷脂进行精液冷冻时可达到最佳的冷冻效果,这些替代方法可减少使用蛋黄所存在的潜在生物安全性问题[12]。Equex STM 也是一种常用的冷冻保护剂,它是基于十二烷基硫酸钠和三乙醇胺的合成物质,因Equex STM 和蛋黄结合时可从蛋黄中提取更多的脂质或脂蛋白到冷冻液中,进而可显著提高精子的冻融质量和受精能力[13-14]。笔者总结了近些年来在猪冷冻稀释液中添加不同冷冻保护剂对猪精子冻融后活力的影响,如表1 所示。
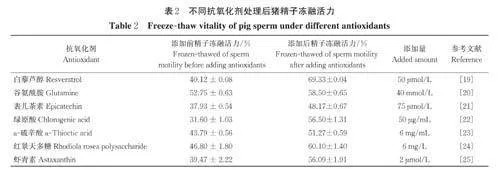
除上述常用的冷冻保护剂外,抗氧化剂常被添加到冷冻稀释液中,如白藜芦醇、谷氨酰胺、表儿茶素、绿原酸、α-硫辛酸、红景天多糖、虾青素等,但冷冻稀释液中添加不同的抗氧化剂提高冻融后精子活力的水平不同( 表2)。
2.2 猪精液冻前平衡和冷却速度
冻前平衡指精液进行程序化冷冻前要经历一段较长时间的缓慢降温平衡过程,该过程需要经过2 个重要的温度节点:17~15 ℃和5~4 ℃,猪精子经历最明显的脂质相变是在15~5 ℃。在降温平衡过程中,当由常温降至10 ℃以下时精子会发生冷休克现象,当继续降温至1~2 ℃时,冷休克现象仍然存在,并会导致更多的精子丧失生存能力。因此,精子在程序冷冻前需在15 ℃平衡2~3 h,以使精子逐渐获得对抗冷休克的能力。在冻前平衡过程中,可通过每间隔40 min 轻微摇晃精液来防止精子发生冷休克现象。有研究表明降温平衡过程可通过维持质膜的脂质结构来增加精子的冷冻耐受性,为达到猪精液最佳冷冻效果,通常冻前降温平衡时间为17~20 ℃保存24 h[26]。平衡降温效果还与其他多种因素有关,如稀释液的种类,当稀释液为AHP(Androhep®Plus)或ASP(Androstar® Plus)时,在10 ℃下平衡24h 的精子表现出更好的低温耐受性,这主要是因为该稀释液包含非动物来源的细胞膜保护成分,会降低精子体外保存时对低温的高度敏感性。当乳糖-蛋黄( 不含甘油)用作稀释介质时,在5 ℃下平衡4 h就能够取得更好的冷冻效果[27]。
除冻前平衡外,程序性冷冻速度也对冷冻效果有很大影响。若冷却速度过快,细胞内的水分不能完全流出,导致水分在细胞内冻结形成冰晶,冰晶将引起精子冷冻损伤;若冷冻速度过慢会导致脱水过多,使得细胞器及膜体积收缩,影响细胞膜脂质-蛋白质复合物、致使大分子变性、诱导不可逆的膜融合,从而造成精子损伤。一般认为猪精液最佳冷冻速度为30~50 ℃/min。采用0.5 mL 冷冻细管进行程序性冷冻时,速度为20、40 或60 ℃/min 和液氮熏蒸均能提高猪精液的冻融效果[28-29]。
2.3 猪冷冻精液解冻
解冻剂种类较多,物质成分各不相同。因此冻精使用不同解冻剂进行解冻时,解冻时间和温度没有统一的标准。解冻剂的研究主要集中在精浆和抗氧化剂方面。精浆作为精液的组成部分,是自然条件下输送精子的必需介质,主要由蛋白质组成,还含有一系列无机离子、盐、糖、柠檬酸、前列腺素和电解质,精浆还为精子存活提供了适合的生理条件,如酸碱度和渗透压等。此外,精浆含有清除小分子自由基的功能,包括维生素C、尿酸、酪氨酸、还原型谷胱甘肽和亚牛磺酸等[30]。因此,精浆在维持精子生存能力及代谢、运动和获能调节中起着关键作用。
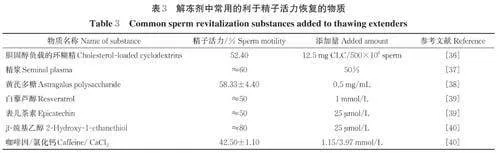
在解冻剂中添加精浆是否有利于精子活力的恢复仍存在争议。众多学者认为冷冻前应通过离心尽可能去除精浆,只留下富含精子的部分进行冷冻[31-33]。在解冻时添加精浆可防止质膜不稳定及其他类似获能的变化,因而添加精浆作为解冻介质有利于维持质膜的完整性,且添加精浆作为解冻介质可维持精子活力,有助于精液在体外保存过程中维持由快速和线性运动精子组成的亚群。研究表明,添加50% 精浆可以最有效地维持精子的活力[34]。但也有研究发现在精子体外保存时,添加10% 的精浆并没有对精子保护起到实质性作用,与精子体外储存能力丧失相关的因素是精液稀释程度而非精浆的有无,精液稀释倍数越高精液活力丧失越快[33]。另有研究认为精浆在冻精解冻后的体外保存过程中对精子的存活是有害的,精浆中含有的内毒素(脂多糖)已被确定为损害公猪精子耐冻性的原因之一,当内毒素(脂多糖)与精子表面存在的Toll 样受体4 结合后可破坏质膜的稳定性[32]。精浆在冻精中的作用存在不同看法的原因可能与精浆的来源、精浆添加量及与精子接触的时间、培养温度等因素相关。除添加一定比例抗氧化剂可提高猪冻精解冻效果外,其他能够激活精子的物质还包括咖啡因,但咖啡因作为解冻剂成分来添加时会造成严重的多精受精现象。有报道发现,使用肌酸和β-甲基环糊素联合处理可以使体外受精双原核率达到70% 左右[35]。笔者总结了近些年在解冻剂中添加不同类型解冻保护物质对精子冻融后活力的影响,详见表3。
此外,解冻速度对精子活力有很大的影响,因而寻找合适的解冻平衡点至关重要。解冻温度包括低温解冻(0~5 ℃)、中温解冻(30~40 ℃)和高温解冻(50~70 ℃),且不同的解冻温度对应不同的解冻时间,如37 ℃解冻20 s、50 ℃解冻16 s、70 ℃解冻8 s 等。低温解冻可能会导致重结晶现象,对精子造成进一步的损伤,但损伤更多是由于渗透性冷冻保护剂不能快速排出所导致。有研究指出70 ℃解冻8 s 后,精子曲线运动速度(curvilinear velocity,VCL)、直线运动速度(straight-line velocity,VSL)、平均运动速度(average path velocity,VAP)和头部摆动振幅(ampli⁃tude of lateral head displscement,ALH)等指标均优于37 ℃解冻20 s[41]。
3 猪精液冻融过程中冷冻损伤及抗冻机制
3.1 猪精液冻融过程中物理结构损伤对精子质量的影响
细胞在冷冻和解冻过程中的物理结构损伤除了低温损伤,还包括中等低温范围(-15 ℃和-5 ℃)的致死性。冷冻流程涉及几个重要温度节点:15~5 ℃冷冻冲击和-5~-50 ℃冰晶形成。当溶液被快速降温至-5 ℃和-15 ℃时会产生自发性的冰核,一旦冰核产生,随着温度的继续降低将向所有方向快速增大,导致精子和溶质都将被限制在未冻结的部分中,从-15 ℃到-160 ℃会促使一些小冰晶和液态水转化为较大的冰晶,随着冰晶的不断形成,精子细胞只能不断堆积到狭窄的未冻结溶液通道中,不可避免地对细胞造成机械损伤[42]。
冷冻保护剂可防止冰晶对细胞的损伤,如甘油可通过氢键相互作用实现高度溶于水,并能透过细胞的质膜,随着冰晶的不断形成,冷冻液中未结晶部分中甘油的浓度逐渐升高,从而降低了这部分未冻结溶质的冰点,进而抑制了冰晶的形成。甘油与水之间的氢键相互作用还可以抑制冰晶的扩散,随着未冷冻部分甘油的浓度不断升高,溶质玻璃化逐渐形成,有效减少了冰晶对精子细胞造成的机械损伤。因此在冷冻过程中,控制冰晶形成的大小和数量,维持冰晶在微晶状态,可以更好地保护精子。
3.2 猪精液冻融过程中功能性损伤对精子质量的影响
1) 氧化应激对精子冻融的影响。氧化应激会引起DNA 损伤,电子呼吸链和线粒体转录系统都会被过量的活性氧(reactive oxygen species,ROS)破坏,而向精液中添加抗氧化剂可抑制ROS 引起的氧化损伤[43]。研究发现,在冷冻过程中产生的ROS 并不多,低温保存可降低线粒体产生ROS 的能力[44]。ROS 产生的时间主要在精液采集后稀释及降温平衡过程和解冻过程,精子ROS 的形成受精子内源性抗氧化物介导,如位于细胞质和线粒体中的超氧化物歧化酶,但致使超氧化物歧化酶浓度升高的确切机制尚不清楚。
与氧化应激关系密切的生物化学反应是脂质过氧化,ROS 和精子功能之间的负相关关系涉及脂质过氧化级联反应,冷冻精子比新鲜精子表现出更高的脂质过氧化水平,而脂质过氧化水平与4-羟基壬烯醛(4-hydroxynonenal, 4-HNE)有关。4-HNE 是一种细胞毒性醛类物质,同时是精子衰老的标志物[45]。4-HNE 是一种强氧化剂,可以与蛋白质、DNA 和脂质等生物分子发生共价结合,从而引起细胞损伤。由于ROS 的生成,精子线粒体基因编码的ETC 蛋白(MT-ND1,MT-ND6)以及MT-TFA 和POLRMT 被4-HNE 修饰,这类蛋白质修饰被认为是氧化应激的关键诱因,造成线粒体DNA 片段降解,转录系统和ETC 呼吸链均受到影响,进而影响精子功能[46-47]。冷冻保存过程中精子内蛋白和DNA结构的变化是冻融精子活力下降的原因,冻融会影响精子头部核蛋白的功能,诱导DNA 结构发生显著变化及鱼精蛋白磷酸化水平发生降低,从而改变精子核中鱼精蛋白-1 与DNA 的相互作用,并破坏核蛋白半胱氨酸基团之间的二硫键,进而影响核蛋白和DNA 之间的结合。因此,精子在冻融过程中,ROS升高会导致DNA 片段损伤和精子核内染色质的变化,从而影响相关基因的转录与翻译,最终导致精子损伤。
2)质膜变化对精子冻融的影响。细胞质膜一般由磷脂、可变数量的甾醇如胆固醇、一定数量的蛋白质等构成,磷脂赋予细胞膜流动性,胆固醇维持刚性和稳定性。随着温度降低,膜脂质会经历物理相的变化,由流体相变为凝胶相,甾醇的存在可抑制这种脂质相的变化,但猪精子质膜上胆固醇与磷脂比例低(猪:0.26,牛:0.45),且胆固醇分布不对称,导致精子质膜对低温引起的脂质物理相的变化更敏感,造成猪精子冻融效果差[48]。猪精子质膜含有高比例的多不饱和脂肪酸,极易受到自由基的攻击,这使得它对氧化应激以及随后的脂质过氧化更加敏感。不饱和磷脂比其他磷脂更早胶凝并发生相分离。因此,在冷冻过程中完整的蛋白质无法进入到膜内结晶凝胶结构域内,造成蛋白质聚集到剩余的液体脂质结构域中,导致许多蛋白质-脂质相互作用丧失,最终导致完整膜蛋白通过脂质相分离变得不可逆地聚集,膜脂质被重组,部分胆固醇分子被释放。目前有研究使用环糊素负载胆固醇处理精液,通过增加精子膜的胆固醇含量提高精子对低温的耐受性,这可能是提高冻融精子质量的一种较好的策略[49]。
膜冷冻损伤会对离子通道的功能造成影响,这也是冻融精子受精能力下降的原因。冷冻会导致精子蛋白质丰度变化,Chen 等[50]通过比较新鲜和冻融的猪精子蛋白质组发现多达41 种蛋白质的数量发生了改变,其中有6 种蛋白质在冷冻保存后含量下降,35 种蛋白质含量上升,这41 种蛋白质可分为10 个不同类别,涉及到多种生理过程,如精子的过早获能、粘连、能量供应以及精子-卵母细胞的结合和融合等,用蛋白质印迹法进一步证实,在冻融精子中,激酶锚定蛋白(3AKAP3)、超氧化物歧化酶1(SOD1)、磷酸丙糖异构酶(TPI1)和外周致密纤维蛋白2(ODF2)的表达增加。另有研究证实TPI1 可作为猪精子冷冻保存前的生物标志物,TPI1 与解冻后精子活力具有高度相关性,被认为能够预测精子耐冻性的高低[7]。
3)酪氨酸磷酸化对精子冻融的影响。精子蛋白的酪氨酸磷酸化发生在输卵管部位,是精子获能的标志。在冷冻过程中由于质膜的相变以及脂质与蛋白质之间相互作用被破坏,导致质膜不稳定并失去选择渗透性的能力,从而增加细胞外离子如Ca2+和HCO3-的流入水平。Ca2+和HCO3-的流入会导致冻融精子出现冷冻获能或类似获能的变化,这也是精子在冻融后受精率发生降低的原因之一。研究证明精子体外保存会导致酪氨酸磷酸化的总体增加,造成一种功能性的、过早发生的获能样状态,这种低温诱导的非调节性获能被称为“低温获能”[51]。
通常酪氨酸磷酸蛋白沿着鞭毛分布,可能参与精子的运动调节和超活化过程,而猪精子中与获能相关的酪氨酸磷酸蛋白定位于头部而不是鞭毛,表明精子头部在获能和顶体反应中起到重要作用。同时,冷冻保存降低了猪精子中胆固醇的含量,又因猪精子质膜的胆固醇、磷脂比例本来就比较低,导致抗冻能力进一步下降,从而使猪精子更容易诱发这种获能样变化。与获能同时出现的是一种酪氨酸磷酸蛋白复合物p32,这是一种前顶体蛋白酶结合蛋白sp32 的酪氨酸磷酸化形式,与顶体蛋白成熟相关,p32 的升高与精子Ca2+水平提高和顶体丢失相关。因此p32 可以作为顶体反应的信号分子,在猪精子冷冻过程中,p32 出现可认为精子发生顶体反应,猪精子冷却至5 ℃比仅冷却至15 ℃产生更大程度的低温获能和自发顶体反应,蛋黄的存在可以明显减少自发的顶体反应,提高解冻后精子的活力。
4 猪精液冻融过程中其他因素对精子质量的影响
4.1 包装形式对精子冻融的影响
冷冻精液常见的包装储存形式有安瓿瓶、颗粒型冷冻保存、铝箔(塑料)袋和麦管灌装。当灌装形式具有较大表面积与体积比时,精液冻融过程中温度变化更加均匀,冷冻过程中有更大的表面来散热,解冻过程中能够迅速升温,冻融效果更好。例如2 mL 扁平细管或5 mL 塑料细管,被称为扁平包装形式,在精子冻融后活力较0.25 mL 细管效果要好,并且其可容纳更多精液,在实际生产时减少解冻细管的根数,减少操作的复杂性。与0.5 mL 细管相比,扁平灌装形式在精子冷冻保存过程中可更加均匀地脱水,但其体积大,所需精液剂量大也是该方法的弊端。
4.2 细菌对精子冻融的影响
精液中细菌的来源包括内源性和外源性2 种。内源性细菌通常包括公猪包皮、尿道、外生殖器,或泌尿生殖道感染导致的细菌,内源性细菌通常对精液质量影响较小。外源性细菌主要与采精过程相关,包括环境条件、采集手法、采精器具等。细菌对冻融精子的影响主要体现在对精子理化性质的影响,如顶体完整性、质膜性质、蛋白质变化等[52]。此外,细菌的存在会造成冻融精子发生凝集现象,进而使精子的活力迅速丧失。为了避免细菌对冻融精子的影响,一般的处理方法是在冷冻液中添加抗生素,但长期使用各种抗生素会增加细菌的耐药性。因此,为尽量避免细菌造成的影响,最好的解决办法是优化收集精液的卫生条件,在采精和冷冻保存过程中尽量减少细菌的污染。
4.3 其他因素对精子冻融的影响
温度和光周期会影响公猪附睾中存储精子的进一步成熟。有研究证明,精液耐冻性取决于精子收集的季节,冬春两季采集的精子比秋、夏两季具有更高的耐冻性,更能耐受冷冻保存[53]。此外,采精的前半部分,尤其是最初的10 mL,比后半部分具有更高的耐冻性,可能的原因是前半部分含有较低水平的碳酸氢盐,或者精液后半部分的精浆会产生负面影响[54]。
精液运输过程也会对精液质量造成一定影响,有研究人员使用特定的模拟设备模拟猪精液运输过程中受振动带来的影响,结果显示,精液在300 r/min频率下、持续进行6 h 圆形水平振动处理,会导致精液介质碱性化,碱性条件会损伤精子的质量,在之后7 d 的储存期间,持续振动处理会进一步导致精子线粒体活性、顶体和质膜完整性降低[55]。因此在精液冷冻前收集和运输过程中应该注重每一步细节,尽量降低对精子造成的不必要损害。
5 提高猪精液冻融效果新方法
5.1 基于纳米技术提高猪精液冻融效果
近年来,纳米技术被逐渐用于畜禽精液净化选择,即根据物质的特性对其进行纳米技术处理,改变物质状态、赋予物质新功能。有研究证实添加新型冷冻保护剂纳米化红景天多糖进行精液冷冻,当添加体积比为15% 时,可将精子冻融后的活力提高11%,达到69%[56]。另有研究在冷冻过程中添加40μg/mL 纳米载体包含的抗氧化剂-白藜芦醇,可减少精子冷冻引起的损伤[57]。此外,纳米技术结合分子标记技术可用于精子生存能力和形态(如头部和尾部异常)检测。通过将生物标志物附着到用于选择性靶向精子的纳米颗粒上,可针对性筛选优质精子,如将磁性氧化铁纳米颗粒(magnetic iron oxidenanoparticles, MNP)与联膜蛋白V 或凝集素结合,再同精液孵育30 min 后使用磁场分离,当纳米颗粒质量为87.5 μg(约218 750 粒纳米颗粒),精子数目为1.6×109~2.0×109 时,可有效去除3% 异常精子,保留97% 优质精子[58]。因此,纳米技术的发展为开发新型猪精子筛选技术、精液选择、精子冷冻提供了新的方向。
5.2 添加耐冻生物标志物提高猪精液冻融效果
蛋白质组学和代谢组学可在特定条件下识别和量化相关分子,检测出的分子可作为精子功能的潜在生物标志物。目前较多研究聚焦分子生物标志物预测精子低温耐受性、减轻冷冻和解冻对精子的不利影响。通过凝胶色谱分离技术将精浆分为2 类不同质量组分:SP1(gt;40 ku)、SP2(lt;40 ku),将这2 种不同质量等级的精浆分别添加在解冻液中,与不添加精浆组相比,这2 种精浆均可显著(Plt;0.01)提高精子运动特性、膜完整性和活力[37]。有研究使用UHPLC-QTOF-MS 技术分析不同冻融能力公猪精浆代谢水平的差异,结果表明D-天冬氨酸、N-乙酰-L-谷氨酸和肌苷的含量存在显著差异[6]。这些发现为探索不同冻融能力的精子代谢生物标志物提供了研究方向。除此之外,精浆中C 型2 类尼曼-匹克蛋白(Niemanne Pick C2 , NPC2)的含量也可预测精子的冷冻能力,这种蛋白对胆固醇有较高的亲和力,在公猪精浆中发现NPC2 的16 ku 和19 ku 2 种亚型在PFE 中的相对水平高于GFE 中,PFE 中NPC2 的含量高于GFE,会导致胆固醇的外排,进而影响质膜的流动性和冷冻效果[7]。针对不同个体猪精液体外长时间保存效果存在差异性进行研究,可筛选出颗粒蛋白前体(granulin precursor)、豆荚蛋白前体(legu⁃main)和AWN 前体蛋白(AWN precursor)3 种候选蛋白,并表明不同个体猪精液体外长期保存能力间的差异可能与精子对自身SP 的耐受性不同相关,而不同个体SP 之间的差异又体现在所筛选出的3 种候选蛋白丰度间的差异。因此,3 种候选蛋白可作为不同个体精子体外保存的标志蛋白,这有助于筛选适宜做精液冻存的公猪个体[59]。总之,应用蛋白组学进行猪精子冷冻损伤机理研究,有助于揭示精子冷冻损伤的分子机理,寻找合适的与精子冷冻性相关的生物标志物,为实现猪精液高效的冷冻保存提供重要依据,笔者总结了猪精子的分子生物标志物及其主要生物学功能(表4)。
6 展望
猪精液冷冻保存是猪精子长期保存最有效的方法,但冷冻过程会对精子造成损害,使得解冻后精子活力、受胎率和窝产仔数方面比鲜精效果差。近年来,国内多家企业开展了冻精技术的探索,相继研发了猪精液冷冻保存技术,并与相关生猪企业进行冻精配种合作,取得了一定成果。有报道称国内某些企业冻精解冻后活力可达70%~80%,且受胎率实现了与鲜精配种同等效果的繁育水平,达90% 以上,使得我国冷冻精液解冻后活力及配种效果达到世界领先水平[62]。但我国目前在猪精液冷冻方面规范化和标准化的技术体系建设上并不完善。因此,应当对猪精液冻融生产工艺中各个流程进行优化组合,删繁就简后制定一套规范化、标准化的猪精液冷冻体系,并建立快速、简便、准确的冻融精子质量评估方法,以实现在生猪生产过程中大规模使用猪冷冻精液,从而降低种公猪引种和饲养管理的成本,减少不必要的生物安全风险。
