
宜宾4 个品种早茶叶可培养内生细菌的比较分析
2024-01-01 00:00:00邓骛远高刚游玲张海英
宜宾学院学报 2024年6期

摘 要:为了比较宜宾不同品种早茶叶可培养内生细菌群落结构,以种植面积较广的4 个早茶品种为材料,采用传统培养方法,分别从其老叶和嫩叶中分离内生细菌,并经16S rDNA 序列分析对其进行鉴定. 结果表明,从4 个品种早茶叶中共分离到内生细菌336 株,归类于18 个属23 个种. 从内生细菌数量和多样性来看,黄芽早gt;福鼎大白gt;中茶302gt;中茶108,且均为嫩叶gt;老叶. 微杆菌属Microbacterium 和黄单胞菌属Xanthomonas 在4 个品种早茶叶中均有分布,且都集中或仅存在于嫩叶,其中Microbacterium 为绝对优势菌群. 各种早茶老叶和嫩叶还有各自的优势菌属短小杆菌属Curtobacterium 等或特异性菌种短小芽胞杆菌B. pumilus、磷酸镰孢菌F. phosphorivorans 等. 说明不同品种早茶叶内生细菌种类组成有明显的相似性,也存在一定的品种或组织专一性. 这些内生细菌及其分布规律可能在较大程度上促成了宜宾早茶的高产优质以及各品种早茶的不同品质特征.
关键词:宜宾;早茶;可培养内生细菌
中图分类号:Q939.11
DOI: 10.19504/j.cnki.issn1671-5365.2024.06.10
茶树(Camellia sin ensis (L.) O. Kuntze)为山茶科、山茶属的木本经济作物. 中国的茶叶种植面积和产销量多年来稳居世界首位,尤其是以宜宾产地为代表的早茶已成我国重要的出口创汇农产品[1-2]. 随着物质生活水平的提高,以及茶文化的深远影响,人们对茶叶的需求量不断增加,且对茶叶的品质有更高的要求. 茶叶的色、香、味等主要品质特征及农残量,是茶叶等级评价的依据和标准,直接影响其消费量[3-4]. 因此,要想提高茶叶市场竞争力,不仅要追求更高的茶叶产量,还需进一步控制农残和改善茶叶品质.
研究表明,茶树的内生菌不仅能促进宿主生长,还能增强其抗病虫害能力等[5-6]. 有文献报道,内生菌对茶叶品质的形成有很大的贡献. 一方面,茶叶内生菌可参与或调节宿主植物茶树叶内活性物质的代谢,从而影响茶叶品质成分的组成;另一方面,在制茶过程中,部分内生菌会参与茶叶的发酵,产生或降解某些滋味物质[7-8]. 不同品种以及同一品种不同生长地的茶树,对环境中营养物质的利用能力与有害因子的抗性,以及茶叶产量和品质等皆有较大差异[9-12]. 由此推知这些差异在较大程度上与茶树内生菌群结构有关. 宜宾市地处四川盆地南缘,金沙江、岷江、长江横穿全市,气候温和,雨量充沛,长年温差不大,无霜期达300 天以上,特别适宜茶树生长,已成为全国最大的生态早茶基地和出口茶生产基地,茶叶种植面积广且品种丰富[2,13]. 得天独厚的自然条件,孕育了世界不可复制的茶业生态,使得宜宾早茶具有“形美、色绿、汤亮、味醇、香雅、早优双绝”特色品质而享誉世界[2]. 因此宜宾地区的茶树可能存在与其他地域环境完全不同的潜在内生细菌物种资源. 而对这些内生菌资源加以开发利用,最首要的一环就是分离鉴定.
茶树内生菌的分离鉴定相关研究起步较晚. 相对较早的是朱育菁等于2008 年从采自福建宁德地区的大白毫和福云六号茶叶中分离到包括红杆菌属、微杆菌属、根瘤菌属、贪噬菌属的16 株细菌和1株真菌[14]. 后来陈百文、汪立群等相继从黄观音、福鼎大白茶、紫鹃、云抗10 号等不同品种的茶树中分离了内生菌[15-16]. 郑世仲,邓晓旭,刘姣姣等分别从茶树叶片、根组织中分离到了对茶树胶孢炭疽菌、腐皮镰刀菌和轮斑病菌有较好抑制效果的内生细菌[6,17-18]. 但针对“中国早茶之乡”——宜宾的不同品种茶树及其不同组织部位展开较为系统全面的内生菌分离鉴定研究还鲜见报道.
本研究立足于“重要经济作物-茶叶”和“新型生物资源-内生细菌”两大要素,以宜宾4 个代表性的早茶品种为材料,比较研究其老叶和嫩叶中可培养内生细菌的群落结构,明确茶叶内生细菌在不同品种茶树和不同组织部位的分布规律,为进一步探究内生细菌对宜宾茶叶产量、品质等的影响提供依据,亦可为优质茶叶生产过程中合理实施必要的微生物调控提供新的微生物来源.
1 材料与方法
1.1 试验材料
试验材料为黄芽早、中茶108、中茶302 和福鼎大白4 个品种早茶树鲜叶,于2022 年夏采自宜宾市金秋湖镇川茶生产基地. 每个试验早茶品种,均随机采集10 株以上无病虫害、生长健壮的茶树鲜叶样品,分老叶和芽叶,每株间隔20 m 以上. 样品存放在采样袋中低温保存,带回实验室后立即开展内生细菌分离.
1.2 试验方法
1.2.1 材料消毒
选取健康茶树鲜叶,用自来水冲洗干净,晾干表面多余水分,用75% 乙醇浸泡1 min,3%~5% 的次氯酸钠消毒6~10 min(不同茶叶略有不同),无菌水冲洗3~4 次,并将最后一次冲洗液涂布于培养基上,26~28 ℃培养2 d,观察平板有无菌落长出,以检测消毒是否彻底.
1.2.2 内生细菌的分离、纯化
将消毒后的鲜茶叶用无菌剪刀剪成0.5~1 cm见方的组织小块,接种于不同培养基上,组织块紧贴培养基表面,每皿均匀放置6~8 块,老、嫩茶叶各接30 个平板,28 ℃恒温培养箱中培养2~3 天. 根据平板上菌落形态特征的差异,对茶叶内生细菌进行初步筛选. 用接种针挑取不同的单菌落,平板划线3~5次,培养至纯菌株.
1.2.3 内生细菌的鉴定
使用TSINGKE DNA 提取试剂盒(通用型)提取和纯化菌株DNA. 具体操作步骤严格按照说明书执行. 以所提DNA 为模板,细菌16S rDNA 通用引物27F: 5'~GAGAGTTTGATCCTGGCTCAG~3'和1492R: 5' ~TACGGCTACCTTGTTACGAC~3'进行PCR 扩增. PCR 反应体系(50 μL):DNA 模板1 μL,引物各2 μL,1×TSE101 金牌mix 45μL.PCR 扩增程序为:98 ℃预变性3 min;循环:98 ℃,10 s;55 ℃,15 s;72 ℃,15 s/kb,39 个循环;7 2 ℃延伸5 min;4 ℃保存. PCR 产物经琼脂糖凝胶电泳检测后,送北京擎科生物科技有限公司完成测序.
1.3 数据处理
用Contigexpress 软件将测序得到的内生细菌株的2 条序列进行校对、拼接,把拼接好的序列提交至NCBI 进行BLAST 分析,比较DNA 的同源性,完成菌株16S rDNA 分子生物学鉴定.
2 结果与分析
2.1 不同品种早茶叶可培养内生细菌的数量
通过分离纯化共得到茶叶内生细菌336 株,其中黄芽早嫩叶109 株、老叶14 株;福鼎大白嫩叶64株、老叶23 株;中茶302 嫩叶60 株、老叶22 株;中茶108 嫩叶38 株、老叶6 株(如图1,因分离到的茶叶内生细菌总的数量较大,文中仅列出来自每种鲜茶叶样品的1 株内生细菌的菌落形态).从可培养内生细菌的数量来看,不同品种茶树及其老叶和嫩叶间均有较大差异,表现为黄芽早gt;福鼎大白gt;中茶302gt;中茶108,在同一品种内,皆为嫩叶gt;老叶.
2.2 菌株的16S rDNA 扩增
提取茶叶内生细菌的基因组DNA,PCR 扩增16S rDNA 序列,经琼脂糖凝胶电泳检测,得到一约1500 bp 长的片段(如图2,因分离到的茶叶内生细菌总的数量较大,对应的PCR 图谱太多,文中仅列出4个品种茶叶部分内生细菌的电泳条带,其余菌株的条带位置与图中一致).
2.3 不同品种早茶叶可培养内生细菌种类及分布
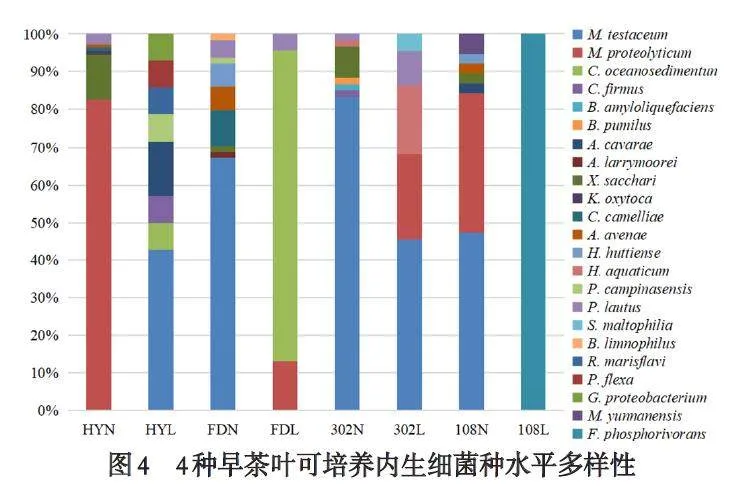
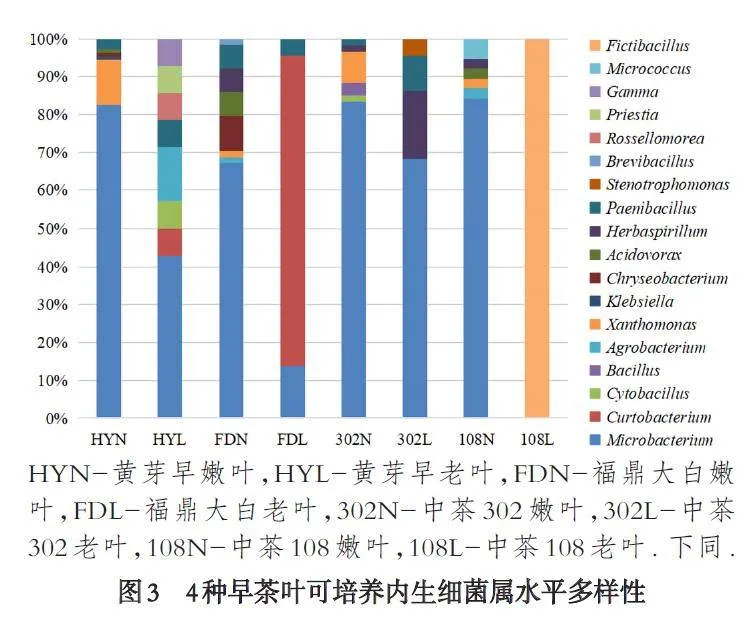
通过测序与同源序列分析,4 种茶树叶中共鉴定到23 种细菌,分别为砖红色微杆菌Microbacteriumtestaceum、微杆菌Microbacterium proteolyticum、短小杆菌Curtobacterium oceanosedimentun、胞囊杆菌Cytobacillusfirmus、解淀粉芽胞杆菌Bacillus amyloliquefaciens、短小芽胞杆菌Bacillus pumilus、卡瓦拉农杆菌Agrobacteriumcavarae、莫氏土壤杆菌Agrobacterium larrymoorei、黄单胞菌Xanthomonas sacchari、产酸克雷伯氏杆菌Klebsiellaoxytoca、金黄杆菌Chryseobacterium camelliae、燕麦食酸菌Acidovorax avenae、赫氏草螺菌Herbaspirillum hut⁃tiense、草螺菌Herbaspirillum aquaticum、坎皮纳斯类芽孢杆菌Paenibacillus campinasensis、灿烂类芽胞杆菌Paenibacillus lautus、嗜麦芽窄食单胞菌Stenotroph⁃omonas maltophilia、沼泽短芽孢杆菌Brevibacillus lim⁃nophilus、蔷薇菌Rossellomorea marisflavi、弯曲芽孢杆菌Priestia flexa、伽马变形杆菌Gamma proteobacterium、云南微球菌Micrococcus yunnanensis、磷酸镰孢菌Fictibacil⁃lus phosphorivorans,其中黄芽早嫩叶鉴定到6 种菌,属水平上分布于微杆菌属Microbacterium、黄单胞菌属Xanthomonas、克雷伯氏菌属Klebsiella、金黄杆菌属Chryseobacterium、食酸菌属Acidovorax、类芽孢杆菌属Paenibacillus,老叶8 种,分布于Microbacterium、短小杆菌属Curtobacterium、胞囊杆菌属Cytobacillus、土壤杆菌属Agrobacterium、Paenibacillus、蔷薇菌属Rossello⁃morea、普里斯特氏菌属Priestia、Gamma;福鼎大白嫩叶9 种,分布于Microbacterium、Agrobacterium、Xan⁃thomonas、Chryseobacterium、Acidovorax、草螺菌属Her⁃baspirillum、Paenibacillus、短芽孢杆菌属Brevibacillus,老叶3 种,分布于Microbacterium、Curtobacterium、Paeniba⁃cillus;中茶302 嫩叶7 种,分布于Microbacterium、Cyto⁃bacillus、芽孢杆菌属Bacillus、Xanthomonas、Herbaspiril⁃lum、Paenibacillus,老叶5 种,分布于Microbacterium、Her⁃baspirillum、Paenibacillus、寡养单胞菌属Stenotroph⁃omonas;中茶108 嫩叶7 种,分布于Microbacterium、Agrobacterium、Xanthomonas、Acidovorax、Herbaspirillum、微球菌属Micrococcus,老叶1 种,假芽孢杆菌属Fictiba⁃cillus(如图3-图4).从物种多样性来看,仍是黄芽早gt;福鼎大白gt;中茶302gt;中茶108,且除了黄芽早的老叶与嫩叶物种多样性比较接近外,其余3 个品种早茶均为嫩叶gt;老叶.
4 个不同品种早茶树的叶有2 个共有内生细菌属,Microbacterium 和Xanthomonas,其中Microbacterium为共有优势菌属,但在不同品种茶叶中存在数量和种的差异,黄芽早嫩叶有M. proteolyticum 90 株,占可培养内生细菌总数的82.57%;老叶有M. testaceum18株,相比嫩叶该属内生菌数量虽不算多,但占老叶的内生细菌总数近一半,42.86%;中茶108 有M. proteo⁃lyticum 14 株,还有同属的M. testaceum18 株,但都存在于嫩叶,该属菌株共占嫩叶内生细菌总数的84.21%;福鼎大白嫩叶有M. testaceum 43 株,占嫩叶细菌总数67.19%,.老叶有M. proteolyticum,仅3 株;中茶302 中只有M. testaceum,嫩叶50 株,占比达83.33%,老叶10株,与黄芽早的老叶相似,也达到该组织内生细菌总数近一半,45.45%. 对Xanthomonas 而言,在4 个不同品种早茶中都仅在于其嫩叶. 此外,每个品种早茶的叶中还存在各自不同种类的优势内生细菌属,如福鼎大白嫩叶中的Chryseobacterium,老叶中的Curto⁃bacterium. 尤其是中茶108 老叶中的优势菌属Fictiba⁃cillus,是在该组织中分离到仅有的1 个属,也是其特有菌属. 相较于其余茶叶品种,黄芽早茶叶同时拥有4 个特异种属,其中Rossellomorea、Priestia、Gamma 分布在老叶,Klebsiella 存在其嫩叶中. 福鼎大白嫩叶有1 个特有属Brevibacillus;中茶302 老叶和嫩叶各有1个特有属,分别为Stenotrophomonas 和Bacillus. 中茶108 嫩叶也有1 个特有属Micrococcus.
3 结论与讨论
本研究从宜宾4 个不同品种早茶的叶中共分离到内生细菌336 株,归类于18 个属,23 个种. 其中Microbacterium 在4 个品种均有分布,且占比最高,为绝对优势菌群,但都主要集中在嫩叶,如在黄芽早嫩叶中的M. proteolyticum,占老叶和嫩叶组织中该属菌株总数的93.75%,福鼎大白和中茶302 的嫩叶中均为M. testaceum,分别占叶内该属菌株总数的93.48%和83.33%;而中茶108 叶中的M. proteolyticum 和M.testaceum,全部存在于嫩叶中,占叶内该属菌株总数的100%. 朱育菁等从福建宁德地区的大白毫和福云六号茶叶也分离到几种较多数量的Microbacterium 属内生细菌,且也都来自嫩叶[14]. 后来汪立群等以栽植于安徽省舒城县德昌苗木有限公司苗圃的3 年生无性系紫娟茶树较嫩的第3 叶为试验材料,也分离到一些Microbacterium 属的内生细菌株[16]. 但两者所分离到的茶叶内生细菌Microbacterium 属在数量和占比上都远没有在宜宾的这4 个早茶品种多,且在同属内的种水平上也存在一些差异. 说明Microbacterium 在茶树中是较普遍存在的内生菌属,且其分布表现出组织差异性,单从叶组织来看,主要分布在嫩叶. 但不同品种或生长在不同地域的茶树,其叶中的Micro⁃bacterium 属内生细菌又存在数量或种类的差异. 推测除了物种本身的遗传因素外,这可能还与茶树生长的地域环境不同有较大关系. 据文献报道,Micro⁃bacterium 菌属可固氮、溶磷以及产植物生长激素,从而能促进植物生长,同时在重金属耐受与植物病虫害防治等也表现出较好的应用潜能[19-21]. 如张志斌等的研究发现分离自东乡野生稻叶组织的内生菌株KlspL18,与M. proteolyticum RZ36T 相似度为97.64%,能产IAA 高达291.7 mg.L-1 菌株发酵液[22].余海忠等从襄麦冬中分离筛选到砖红色微杆菌M.testaceum,还能产生其宿主植物的主要活性成分甾体皂苷,可能存在与宿主植物相同或相似的代谢途径,进而影响着植物代谢产物的含量[23]. 根据以上分析,可以判断宜宾早茶中叶存在的较大比例的M. proteo⁃lyticum 与M. testaceum 对茶树的生长、抗逆性及茶叶品质成分等方面可能产生了重要影响.
此外,X. sacchari 在4 个茶叶品种也皆有分布,且仅存在于各品种早茶的嫩叶中. 从已发现的来自其他物种植物的X. sacchari 内生细菌来看,该种菌也具有较多生物学功能. 如郭鹤宝从水稻种子中分离培养到内生细菌群X. sacchari,能分泌IAA,促进水稻和拟南芥幼苗的生长;除此之外,还对番茄青枯以及水稻白叶枯细菌病原菌具有拮抗作用,并能通过茎叶的侵染定殖水稻体内,且未发现该类群对水稻具有致病性,因此认为该菌具备用于生物肥料生产的基本条件[24]. 据此可以推测宜宾早茶嫩叶中的X. sac⁃chari 在促进茶树生长与芽叶萌发,增强茶树抗病虫害能力,提高茶叶产量方面也许发挥了较大作用.
本研究结果还显示,除了4 种早茶共有的内生细菌种群外,各品种茶树的老叶和(或)嫩叶还有各自的优势菌属或特异性菌属,而这些内生细菌也许是茶叶生长习性、品质特征等形成的又一促成因素,如中茶302 嫩叶中的B. pumilus. 梁静娟等从广西北部湾红树林海洋淤泥中筛选到的B. pumilus PLM4 具有产生抗肿瘤多糖的能力[25]. 孙迅等从34 株细菌中筛选到菌株B. pumilus H-101 高产木聚糖酶[26]. 罗晶晶等的研究结果表示,通过在萎凋过程中添加木聚糖酶等外源酶能改变茶叶中的某些化学成分,如茶多酚、茶氨酸等物质有明显增加,而可溶性糖有减少[27]. KOMERA Irene 等利用重组枯草芽孢杆菌产B. pumilus 来源的γ-谷氨酰转肽酶能高效合成L-茶氨酸[28]. 因此,中茶302 嫩叶中的B. pumilus 极有可能对茶叶中诸如茶氨酸、茶多酚、茶多糖等物质成分的含量产生了一定的影响. 再如108 老叶的特异性菌群F. phosphorivorans,同时也是从中分离到仅有的可培养内生细菌种群. 袁梅等从湖南水稻根部中分离到编号为sd352 的一株F. phosphorivorans 内生细菌,以及郭雨晴等从荒漠珍稀植物根际土中分离到的编号为HD3 的该种菌株,皆有较显著的促生作用[29-30]. 当然除了以上着重分析的内生菌群外,还有本研究中一些较少提及的内生细菌,其对茶树潜在的生物学功能尚不清楚,有待进一步研究.
综上,不同品种茶树叶内生细菌分布规律的不同,但又存有相似,可能在一定程度上造就其在生长特点和茶叶品质方面的共性和差异. 除了物种的遗传背景以外,生态环境与种植条件等是其重要影响因素. 宜宾是世界茶叶原产地之一,土质优良,具有非常适合茶树生长且有别于他地的生态环境,加之茶叶基地的管理标准化、生态化,使得宜宾早茶采摘早、采期长,茶叶香高味爽、病虫害少、卫生安全[2].本研究中4 种宜宾早茶叶均以Microbacterium 为绝对优势菌属,以及X. sacchar 都仅存在于各品种早茶嫩叶的内生细菌分布特点,可能是宜宾早茶具有上述品质的共同原因. 此外,同在宜宾种植的不同品种早茶在生长状况、茶叶产量和香气、滋味等感官品质方面又有各自不同的特点,这可能与各品种的共有内生细菌属的占比不同以及优势或特异性内生细菌种属的差异有关. 由此看来,若对这些来源于宜宾茶叶生态的内生细菌,尤其是那些经推测有促进茶树生长和提高茶叶品质的潜力菌株开展进一步研究,有望从中挖掘出有利于茶叶提质增产的功能菌株,并将在茶产业及其相关领域具有较好的应用前景.
参考文献:
1] 姜仁华, 陈富桥. 高质量发展背景下我国茶产业的业态创新之路[J]. 茶博览, 2020(6): 18-23.
[2] 王太, 彭小元, 辛燕, 等. 宜宾早茶久负盛名饮茶之风浸梁东南[J]. 农产品市场周刊, 2017(18): 34.
[3] 杨益欢, 禹利君, 贺军辉, 等.“ 发花”对不同等级莓茶风味品质的影响[J]. 食品工业科技, 2022, 43(17): 87-94.
[4] 刘腾飞, 张丽, 杨代凤, 等. 气相色谱法测定茶鲜叶中4 种有机磷农药残留量的不确定度分析[J]. 食品工业科技,2023, 44(1): 323-331.
[5] 武警, 陈楠楠, 韩梦琳, 等. 茶树根系耐铝促生内生细菌的分离鉴定及其特性研究[J]. 茶叶科学, 2022, 42(5):610-622.
[6] 郑世仲, 周子维, 陈晓慧, 等. 拮抗炭疽病的茶树内生菌筛选, 鉴定及培养条件优化[J]. 茶叶科学, 2023, 43(2):205-215.
[7] 杨瑞娟, 王桥美, 彭文书, 等. 茶窖中微生物群落分布多样性及普洱茶内生菌和茶品质的研究[J]. 热带农业科学,2021, 41(1): 97-105.
[8] 路伟尧. 普洱茶发酵微生物的溯源分析[D]. 北京: 北京化工大学, 2013. .
[9] 杨霁虹, 周汉琛, 刘亚芹, 等. 基于HS-SPME-GC-MS 和OAV 分析黄山地区不同茶树品种红茶香气的差异[J]. 食品科学, 2022(16): 235-241.
[10] 王君雅, 陈玮, 刘丁丁, 等. 不同品种茶树新梢响应“倒春寒”的转录组分析[J]. 茶叶科学, 2019, 39(2): 181-192.
[11] 疏再发, 郑生宏, 邵静娜, 等. 不同茶树品种(系)对减半施肥的响应研究[J]. 茶叶科学, 2022(2): 277-289.
[12] 郑琳, 刘盼盼, 金孝芳, 等. 不同茶树品种(系)黄大茶的品质分析[J]. 食品工业科技, 2022, 43(21): 310-318.
[13] 唐茜. 生态早茶基地标准化茶园建设的关键技术[J]. 宜宾科技, 2011(2): 42-45.
[14] 朱育菁, 陈璐, 蓝江林, 等. 茶叶内生菌的分离鉴定及其生防功能初探[J]. 福建农林大学学报(自然科学版), 2009, 38(2): 129-134.
[15] ]陈百文, 刘伟, 赵恬欢, 等. 对茶树炭疽病菌具拮抗作用的茶树内生细菌的筛选[J]. 福建农林大学学报(自然科学版),2010, 39(4): 341-346.
[16] 汪立群, 颜小梅, 郭小双, 等. 紫娟、云抗10 号两个茶树品种内生菌多样性研究[J]. 安徽农业大学学报, 2016, 43(1):1-5.
[17] 邓晓旭, 谢夏, 潘娅梅, 等. 茶树腐皮镰刀菌拮抗菌株的筛选鉴定及促生防病特性分析[J]. 茶叶科学, 2023, 43(1):67-77.
[18] 刘姣姣, 严哲伟, 张义菊, 等. 一株沙福芽胞杆菌ZG6 的筛选, 鉴定及其对茶树轮斑病的生防潜力研究[J]. 中国生物防治学报, 2023, 39(1): 212-220.
[19] MAHMOOD A, KATAOKA R. Metabolite profiling reveals acomplex response of plants to application of plant growth-promoting endophytic bacteria[J]. Microbiological Research, 2020,(234): 126421. doi:10.1016/j.micres.2020.126421.
[20] BANACH A, KUŹNIAR A, MARZEC-GRZĄDZIEL A,et a1. Phenotype switching in metal-tolerant bacteria isolated from a hyperaccumulator plant[J]. Biology, 2021, 10(9): 879.doi:10.3390/biology10090879.
[21] 赵晶, 邢志富, 田茂雄, 等. 微杆菌Sneb159 杀线虫活性物质的分离与鉴定[J]. 微生物学报, 2020, 60(10):2341-2349.
[22] 张志斌, 朱晓湉, 刘婷婷, 等. 高产吲哚乙酸东乡野生稻内生微杆菌KlspL18 分离及鉴定[J]. 微生物学报, 2022, 62(11): 4108-4121.
[23] 余海忠, 程旭, 王海燕, 等. 不同生育期襄麦冬内生细菌的分布及产甾体皂苷菌株的初步筛选[J]. 天然产物研究与开发, 2022(6): 1027-1037.
[24] 郭鹤宝. 水稻种子内生Xanthomonas sacchari 系统发育及其植物促生特性研究[D]. 北京: 中国农业科学院, 2020.
[25] 梁静娟, 王松柏, 庞宗文, 等. 海洋细菌Bacillus pumilusPLM4 产抗肿瘤多糖的发酵条件优化研究[J]. 广西农业生物科学, 2006, 25(3): 256-260.
[26] 孙迅, 王宜磊, 邓振旭, 等. 木聚糖酶高产菌株Bacillus pumilus H-101 的筛选及产酶条件的研究[J]. 微生物学杂志, 1997, 17(2): 17-22.
[27] 罗晶晶, 王登良. 不同外源酶添加对夏茶金观音红茶品质的影响[J]. 蚕桑茶叶通讯, 2014(6): 17-19.
[28] KOMERA I, 杨套伟, 张显, 等. 利用重组枯草芽孢杆菌产Bacillus pumilus 来源的γ-谷氨酰转肽酶及其在L-茶氨酸合成中的应用[J]. 食品与生物技术学报, 2019. 38(10): 67-71.
[29] 袁梅, 谭适娟, 孙建光. 水稻内生固氮菌分离鉴定、生物特性及其对稻苗镉吸收的影响[J]. 中国农业科学, 2016, 49(19): 3754-3768.
[30] 郭雨晴, 赵世超, 徐道龙, 等. 3 种荒漠珍稀植物根际促生菌的筛选、鉴定及对高粱幼苗生长的影响[J]. 草地学报, 2020,28(4): 1121-1128.
【编校:许洁】
基金项目:四川省科技厅面上项目(2021YJ0292);固态发酵资源利用四川省重点实验室开放基金(2018GTJ002);香料植物资源开发与利用四川省高校重点实验室开放基金(2018XLZ006)
猜你喜欢
名家名作(2023年34期)2023-03-24 11:05:46
文萃报·周五版(2022年38期)2022-07-04 11:20:55
华人时刊(2022年1期)2022-04-26 13:39:40
海峡姐妹(2020年11期)2021-01-18 06:16:22
云南画报(2020年11期)2020-12-14 07:17:32
文艺生活·中旬刊(2020年12期)2020-04-08 02:44:57
四川省干部函授学院学报(2019年1期)2019-06-25 00:53:36
岷峨诗稿(2017年4期)2017-04-20 06:26:33
中国调味品(2017年2期)2017-03-20 16:18:19
中国茶叶(2016年3期)2016-01-18 04:54:08
