
宁德三都澳海岛马尾松林群落特征及物种多样性
2023-12-26 02:44:56肖集泓赖敏英王春晓钟址非温清燕钱志军王迎雪邓清雅邓传远
生态学报 2023年23期
肖集泓,赖敏英,李 媚,王春晓,钟址非,温清燕,钱志军,罗 真,王迎雪,邓清雅,邓传远
福建农林大学风景园林与艺术学院,福州 350002
因地理位置特殊,海岛常年风速较大,受台风浪潮侵袭,土壤瘠薄盐分含量高,且因规模有限和空间隔离,海岛生态系统具有明显的独立性与资源短缺性,属于典型的脆弱型生态系统,在独特的自身环境与复杂的干扰下,表现出易受损性和难恢复性[1]。森林生态系统作为组成结构复杂且物种资源最为丰富的生态系统类型,不仅提供了重要的自然资源,而且对保护脆弱的生态系统具有重要的意义。大量的原始天然林被破坏后,许多海岛通过人工林种植和生态保护政策管理等人为调控方式进行植被重建和修复,并借此减弱和消除海岛生态脆弱性[1—3]。海岛人工林多以单一树种进行种植建设。单一树种种植的人工林群落结构简单,抵抗病虫害和自然灾害的能力较弱,林下植被和其他地被植物及微生物等相对缺乏,因此常表现出林分物种多样性极低,森林群落生态服务功能性差,地被植物及地表枯落物少,土壤养分等肥力较低,保持水土及通养水源等方面能力极弱等缺点[2—4]。且目前仍然缺乏相关技术和研究资料以支撑我国海岛植被修复,导致野生植物资源衰退严重,修复选用的物种也因不适应海岛或群落的环境而被淘汰[5]。因此,对相对稳定的海岛半自然林进行相关研究,不仅能够丰富海岛植物资料,了解人工林向自然林演替过程中植物组成和变动,同时能够了解演替过程、植被稳定性、群落多样性等特点,及了解限制群落演替进程的各种影响因子,这对海岛生态和植被的修复等具有十分重要的指导意义。
群落的结构特征和物种多样性是生态学研究的热点之一[6]。目前,国内外学者多集中于对内陆和外海岛屿的植被进行研究[7—10],对内海岛屿的相关研究较为缺乏。三都澳海岛属内海岛屿,该区域也是南亚热带和中亚热带的过渡区域,地理位置十分重要,但目前对于该区域植物研究仅见对苔藓植物多样性的调查[11]。马尾松(Pinusmassoniana)是松科(Pinaceae)松属常绿针叶大乔木,适应性强,耐干旱瘠薄,是恢复退化土地的优秀先锋树种[12]。马尾松因适应性强,在三都澳地区,特别是环境较内陆恶劣的海岛上广泛分布,占据着当地最大的森林面积,是当地最具代表性的森林类型,具有不可替代的生态价值。三都澳地区的马尾松林为上个世纪飞机撒播造林或人工造林形成,受当地政策的制约,人为影响相对较小,属于半自然群落。自然林的保护和恢复是目前我国实施的一项庞大而复杂的社会工程,自然林资源的合理开发和研究逐渐成为社会关注的热点问题[4]。自然或半自然林林下植物丰富多样,伴随着群落的演替更新和植物的优胜劣汰,适合在群落中生存的优势物种逐渐体现,共同构成了相对稳定的群落结构。因此,对三都澳半自然马尾松林进行群落特征和物种多样性研究,有利于了解其群落中的主要物种组成及相对稳定的群落结构特点,这对当前海岛植被修复及海岛人工林建设具有一定的指导意义。且当地的马尾松林存在植株老化死亡、自然更新困难等问题,其未来的持续经营和管理也亟需相关研究提供基础资料参考。因此本研究通过野外调查三都澳6个海岛的马尾松林,分析其物种组成和植物功能性状,同时划分其植被类型,探究其演替规律,并分析物种多样性及其与环境因子的相关性,旨在摸清三都澳海岛马尾松林群落特征、演替状况,及其物种多样性与环境因子的相关关系,丰富当地植被基础资料,弥补内海岛屿相关研究领域的空缺,为海岛植被修复、海岛人工林建设及当地马尾松林的合理经营和管理提供理论基础。
1 材料和方法
1.1 研究区概况
三都澳位于福建省宁德市东南部,为中国1.84万km黄金海岸线的中点,距宁德市区30 km,是世界级天然深水良港。共有岛屿126个,有居民岛屿17个,其中最大的岛屿是三都岛,总面积29.3 km2,最高海拔461 m,青山岛、长腰岛、白匏岛、鸡公山岛和斗帽岛面积分别为9.48 km2、2.22 km2、1.12 km2、0.89 km2和0.6 km2[11]。研究区属典型的亚热带海洋季风气候,地形以丘陵为主,土壤类型以红壤和黄壤为主,岛上以次生马尾松(Pinusmassoniana)针叶林和马尾松针阔混交林分布最广。
1.2 样地设置与调查
基于实地踏查,于2022年6—8月采用典型选样的方法对三都岛、青山岛、长腰岛、鸡公山岛、白匏岛、斗帽岛6个面积较大和森林覆盖率较高的有居民海岛进行样地设置,6个海岛分别设置有20、6、5、4、3和2个样地(图1),共设置40个20 m×20 m的森林样方。以20 m×20 m作为乔木层样方面积,森林样方4个角设置4个面积为5 m×5 m的灌木样方,4个角及中心位置设置5个面积为1 m×1 m的草本样方。记录乔木层(株高≥ 5 m或胸径≥ 3 cm的乔木、竹类、大型灌木、附生植物和藤本植物)、灌木层(株高<5 m且胸径<3 cm的灌木、攀援灌木、乔木幼树和竹类,统计时剔除以攀援状依附植物生长的攀援灌木植株)和草本层(含攀援草本和蕨类,统计时剔除以攀援状依附植物生长的攀援草本植株)每一棵植株的种名、胸径、株高、冠幅(或盖度)、数量等信息[13]。采用全球定位系统(GPS)和森林罗盘仪记录样方的经纬度、海拔、坡向、坡度和坡位。将坡位划分为上、中、下坡;坡向划分为阳坡(157.5°—247.5°)、半阳坡(112.5°—157.5°、247.5°—292.5°)、阴坡(0°—67.5°、337.5°—360°)、半阴坡(67.5°—112.5°、292.5°—337.5°)[14]。采用林冠数字图像Photoshop分析法[15]测定群落郁闭度,样方基础信息见表1。

表1 样方概况

图1 三都澳6个海岛区位
1.3 数据处理与分析
根据《中国植物志》[16](http://www.iplant.cn/foc)和《福建植物志》[17]整理研究区植物名录,记录分布区域,并区分乡土植物和外来植物,采用最新修订的拉丁名和中文名。由于亚热带地区群落结构和物种组成复杂,应通过数量分类方法(聚类分析)进行植物群落分类[13],因此,根据群落的物种重要值(剔除重要值<5的偶见种),采用目前植物群落分类中较为常用的组平均聚类法[9—10, 18]划分植物群落类型。利用冗余分析(RDA)解释环境因子对物种多样性的影响。利用二次函数拟合分析灌木层鹅掌柴(Heptapleurumheptaphyllum)与群落郁闭度的关系。重要值和物种多样性计算公式[19—20]如下:
重要值=(相对多度+相对频度+相对显著度)/3
(1)
Shannon-Wiener指数(H):
(2)
Simpson指数(D):
(3)
Pielou指数(E):
(4)
Patrick指数(R):
R=S
(5)
式中,各层片相对显著度分别采用乔木胸高(1.3 m处)断面积和灌草层植物盖度计算。S为各样方中出现的物种数,N为所有物种个数之和,Pi=Ni/N,Ni为第i个物种的个体数(i=1,2,3...)。
群落多样性指数=0.5D1+0.3D2+0.2D3
(6)
式中,D1、D2和D3分别表示乔灌草3个层片的多样性指数[9, 21]。利用SPSS软件的单因素方差分析计算各群系、各植被型的多样性指数,同时选择最小显著性差异法(LSD)进行事后方差比较不同群系、及各层片之间的多样性指数差异水平,由于植被型只有两种,因此选择t检验进行多样性指数差异性水平分析。通过方差分析比较本研究地与平潭海岛[9]森林植被物种多样性的差异。
采用五点取样法采集各样方25 cm深的土壤样品,研磨过筛后重复3次测定其9项指标:pH值、全钾、速效钾、全磷、有效磷、全氮、碱解氮、有机质和水溶性盐总量[22],土壤信息见表2。

表2 土壤化学指标
分析群落中种子植物的生长型、生活型、对光的耐受性3项定性功能性质,具体划分依据为:生长型分为乔木、灌木、草本、藤本;生活型划分为9类,即高位芽植物(含大型>30 m、中型8—30 m、小型2—8 m和矮小型0.25—2 m共4类)、地上芽、地面芽、藤本附生和一年生植物;植物对光的耐受性分为阳性、中性和阴性[9, 23]。
2 结果与分析
2.1 物种组成和植物功能性状
统计40个样方调查数据,发现三都澳海岛马尾松群落内维管束植物共计255种(含5变种),隶属于86科168属,属种系数为66.80%。其中以被子植物为主,共计72科150属231种(含5变种),蕨类植物10科16属22种,裸子植物仅2科2属2种,即马尾松和杉木(Cunninghamialanceolata)。研究区植物来源统计,乡土植物78科159属244种,外来植物7科12属12种,其中含1级恶性入侵植物2种,即钻叶紫菀(Symphyotrichumsubulatum)和小蓬草(Erigeroncanadensis),但在林中重要值占比很低。
对255种维管束植物的3个定性功能性状进行统计分析(见表3)。由表3可知,在生长型上,以乔木和灌木植物种类最多,分别占总物种数的37.25%和30.20%;在对光的耐受性上,以中性植物数量最多,占总物种数的69.02%,其中阳性树种都为木本植物;在生活型上,以小型、矮小型和中型高位芽植物为主,分别占总物种数的29.80%、27.84%和24.71%,大型高位芽植物和一年生植物占比极小,分别占比1.57%和2.35%。
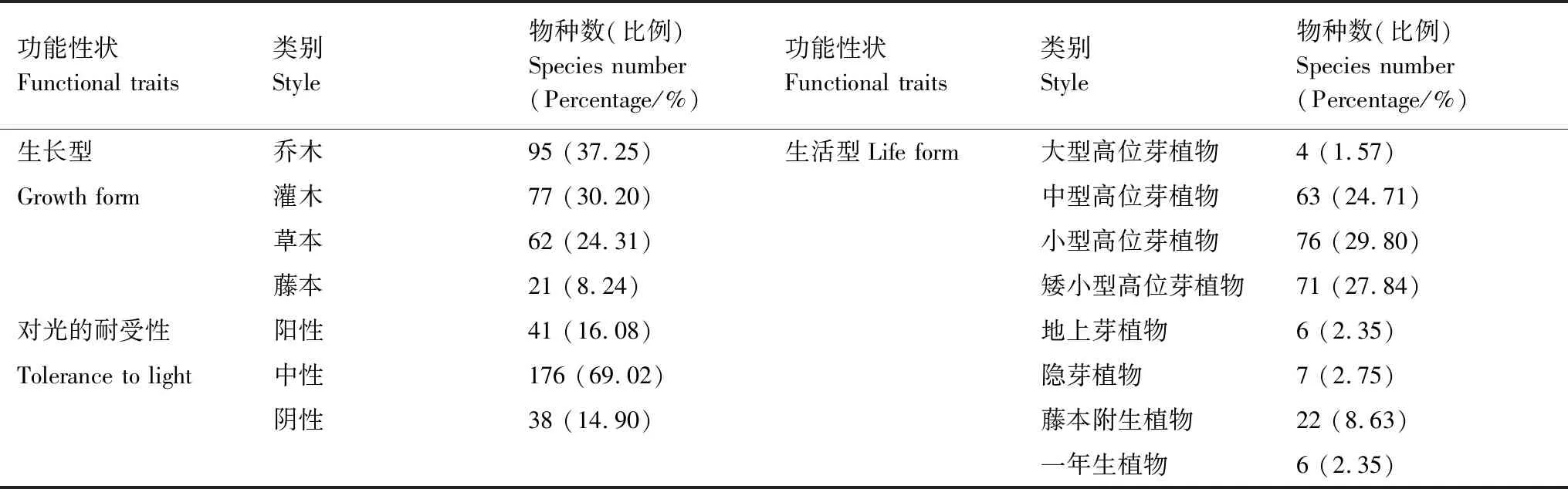
表3 三都澳6个海岛马尾松群落维管束植物功能性状统计
2.2 植物群落类型划分
参照《<中国植被志>研编内容与规范》有关植被分类原则和群落命名原则[13],选取40个样方中重要值>5%的物种,利用组平均聚类法对各个样方的物种重要值进行聚类分析(图2)。由图2可知,当欧氏距离为20时,由上至下将三都澳6个海岛的马尾松群落划分为5类群系:马尾松林、马尾松+檵木(Loropetalumchinense)林、马尾松+鹅掌柴林、马尾松+杨桐(Adinandramillettii)林、马尾松+茶竿竹(Pseudosasaamabilis)林,最终划分为33类群丛(见表4)。统计40个样方各层片重要值占比最大的优势种重要值和数量(见表5),由表5可知,40个样方的乔木层优势种均为马尾松,灌木层优势种主要有:鹅掌柴、杨桐、九节(Psychotriaasiatica)、桃金娘(Rhodomyrtustomentosa)、玉叶金花(Mussaendapubescens)、豺皮樟(Litsearotundifoliavar.oblongifolia)等,草本层最主要优势种为芒萁(Dicranopterispedata),此外还有团叶鳞始蕨(Lindsaeaorbiculata)、扇叶铁线蕨(Adiantumflabellulatum)、狗脊(Woodwardiajaponica)等优势种。

表4 三都澳6个海岛马尾松群落植被类型

表5 三都澳6个海岛马尾松林植被特征及物种多样性指数

图2 植物群落组平均聚类
2.3 植物群落物种多样性与环境因子的相关性
根据调查数据计算40个样方各层片及群落整体的物种多样性指数(包括Patrick指数、Simpson指数、Shannon-Wiener指数、Pielou指数),见表5。由表5可知,在群落各层片上,灌木层的物种多样性指数均远高于同样方的乔木层和草本层。对不同群系、植被型的样方进行多样性指数统计计算,并标记显著性差异(见表6、表7)。由表6、表7和方差分析及t检验结果可知,5类群系和2种植被型的灌木层多样性指数(除E外)均显著(P<0.05)高于乔木层和草本层,表明研究地群落以灌木层物种最丰富、群落最稳定、结构最复杂。从植被型差异来看,马尾松常绿针阔混交林仅乔木层D和群落整体E显著(P<0.05)高于马尾松常绿针叶林,其他指数差异不显著(P>0.05)。从群系差异来看,5类群系乔木层多样性指数差异均不显著(P>0.05),II群系的灌木层R显著(P<0.05)低于其他4类群系,I群系的草本层R显著(P<0.05)高于其他群系,II群系的草本层H、D、R均显著(P<0.05)低于其他4类群系,II群系的群落整体H、D、R显著(P<0.05)低于其他3类混交林群系,V群系的群落整体H、D均显著(P<0.05)低于除II外的其他混交林群系。

表6 不同群系的物种多样性指数

表7 不同植被型的物种多样性指数
对40个样方的物种多样性指数(含群落整体和各层片的Patrick指数、Simpson指数、Shannon-Wiener指数、Pielou指数,见表5)和14个环境因子(含表2的9个土壤因子、表1的4个地形因子(海拔、坡向、坡度、坡位)和群落郁闭度)进行冗余分析(RDA)分析,结果见图3。分析结果筛选出2个显著影响(P<0.05)物种多样性的环境因子:土壤pH和速效钾(AK)。土壤pH与群落Patrick指数、Shannon-Wiener指数及灌草层物种多样性指数呈正相关,与群落Pielou指数、Simpson指数及乔木层物种多样性指数呈负相关;速效钾与群落整体、乔木层、灌木层和草本层的物种多样性指数均呈正相关。而土壤pH和速效钾这两个环境因子共同对物种多样性的总解释量为27.6%,表明还有72.4%的未知因素影响了三都澳海岛的马尾松群落物种多样性。

图3 物种多样性指数与环境因子RDA分析图
鹅掌柴是研究地灌木层重要值最大的物种,属半阳树种,能于全日照、半日照、半荫庇的环境中生存,以半阳或者半阴的环境生长最佳。由于RDA分析反映的是线性回归分析结果,不能反映出非线性相关模型,因此对灌木层鹅掌柴的重要值与群落郁闭度进行二次函数拟合(剔除群落中未出现鹅掌柴的4个样方S2、S6、S8、S18),结果见图4。可知,灌木层鹅掌柴的重要值与群落郁闭度符合二次函数关系,二者呈显著(P<0.05)相关关系。当郁闭度过高或者过低时,鹅掌柴的重要值均占比极小,符合鹅掌柴对光照的适应性结果。

图4 鹅掌柴重要值与群落郁闭度二次函数拟合
3 结论与讨论
3.1 三都澳6个海岛马尾松群落分类和演替
在前人的研究中发现,自然或近自然的亚热带森林植被以中型和小型高位芽植物占比为主,一年生植物相对较少,而次生群落则表现出灌木和(一年生)草本植物占比优势明显,高位芽植物生态位丧失[9,24—25]。本研究区以小型、矮小型和中型高位芽植物为主,一年生植物占比极小,乔木树种占比最大,表现为近自然状态的植被类型,而矮小型高位芽植物占比大可能是受海岛环境影响,植物的适应性选择[10]。但生境异质性会导致同一植物物种表现为不同的生活形态[10],受海岛环境的限制与影响,研究区内的大型高位芽植物种类占比最小,研究地记录的最高物种为马尾松,属于大型高位芽植物(30m以上),但群落中最高统计记录不超过25 m,计算其平均高度仅10 m左右,且本研究地记录的95种乔木树种中,乔木层仅见81种,其他14种以灌木状分布于灌木层,具体体现为灌木层多样性指数(除E外)均明显高于乔木层(表5、6和7),表现出了次生群落的特征。因此,可以将研究地的植被类型划分为具次生性质的自然植被,或称之为半自然植被[26]。
出现在乔木层的竹类、大型灌木、附生植物和藤本植物是乔木层中稳定的组成成分,因此要统计为该层的物种,而根据需要,乔木层可进一步划分为大乔木层(高度≥25 m)、中乔木层(8 m≤高度<25 m)和小乔木层(5 m≤高度<8 m)3个亚层[13]。研究地记录的最高物种为马尾松,高度未超过25 m,且计算每个样方的物种平均高度发现,马尾松平均高度9.1—12 m不等,而乔木层中重要值较高的鹅掌柴、杨桐、茶竿竹、檵木平均高度为5.5—7.5 m不等,乔木层几种优势植物平均高度差异不明显,从实际生长高度来看都属于中小型乔木,因此乔木层不做进一步划分。根据各群落中物种重要值的组平均聚类,将马尾松群落划分为5类群系,分属于2种植被型(见图2和表5)。由于灌木层结构复杂,且部分样方的灌木层和草本层优势种不明显,表现出同一层片多优势种共存的现象,符合亚热带群落结构和物种组成复杂多样的特征[13],因此最终划分为33类群丛(见表5)。5类马尾松群落类型中,乔木层重要值最高的都是马尾松,是绝对优势树种。大多数样方草本层重要值较高的物种较为相似,以芒萁重要值占比较大,表明三都澳海岛马尾松群落林下草本层重要值较高的物种种类受林分类型影响不大,但不同林分类型中,林下灌木层各物种重要值存在较大差异,这与张维伟等[27]的研究结果类似。在群落中,物种之间的相互作用会影响群落的物种组成及其个体数量[28],同时外界的干扰也会导致物种组成差异[29]。由图4可知,当群落郁闭度过高或者过低时,鹅掌柴重要值均较小。根据重要值数据,在S10—12、S19和C2—3这6个郁闭度>0.85的样方中,灌木层中鹅掌柴的重要值仅有5.28、1.32、3.82、0.57、0.72和0.54,鹅掌柴属半阳树种,以半日照或者半阴环境生长最好,过高的林分密度则不利于鹅掌柴的生长。在S11—12两个马尾松-鹅掌柴混交林中,鹅掌柴虽在乔木层占据极大的重要值,但在高郁闭度的林下灌木层仍然难以生存,长此以往,这些样方的鹅掌柴将逐渐被其他物种淘汰,使得林分结构发生改变,群落稳定性受到影响。同理,马尾松属阳性植物,在灌木层仅在S9和C5两个极低郁闭度的马尾松林中出现,在林分结构复杂且郁闭度过高的群落中明显存在自然更新问题。因此,本研究5种类型马尾松群落灌木层之所以出现物种组成及其重要值差异,原因之一极可能是受郁闭度的影响。
从演替方向看,研究地以人工马尾松常绿针叶林作为起点,起初受人为砍伐破坏的影响,群落演替速度较慢,后经当地政策的干预,人为干扰逐渐减弱,在群落恢复过程中表现出次生性质。随着时间的推移,群落正逐渐恢复至近自然状态,但是目前仍然存在次生群落的特征。群落由常绿针叶林逐步向常绿针阔叶混交林演替的过程中,强阳性针叶树种马尾松将逐步被其他阳性阔叶树种取代,如鹅掌柴、杨桐等,阔叶树种将逐渐占据群落的主要地位,群落郁闭度也将逐渐增加。当郁闭度高达一定的程度时,不耐荫蔽的阔叶树种如鹅掌柴等,在高郁闭度的林下将难以生存,自然更新能力下降,生态位逐渐降低,将被逐步淘汰,而强耐阴的阳性树种如杨桐等,将逐步成为该区域稳定群落的优势种或建群种。最终,研究地群落将自然演替为亚热带顶级群落-常绿阔叶林,同时针叶林林下草本层最主要优势种芒萁也将随着群落从针叶林向阔叶林的演替而逐渐丧失优势地位。
3.2 三都澳6个海岛马尾松群落物种多样性特征及环境因子解释
Shannon-Wiener指数(H)越高,群落越复杂,生物种类越多;Simpson指数(D)越大,群落内物种数量分布越不均匀[7]。本研究中,不同林分类型物种多样性有所不同,总体而言,马尾松林的群落整体H、D均显著(P<0.05)低于除II外的其他混交林(表6)。主要原因是,混交林中乔木层植物种类多,生态位差异大,对资源利用更充分[27]。而单一马尾松林乔木层形成的单一胁迫,会降低林下植物的数量,进而影响其物种多样性指数,且植物数量的减少难以提供足够的凋落物供给微生物分解,使得马尾松林内的生境条件比其他混交林差,进一步降低了物种多样性指数。同时,根据表2土壤数据可知,速效钾含量最高的10个样方中,除C4属于马尾松林外,其他都是混交林,从RDA分析结果也发现,速效钾与各层片物种多样性指数均呈正相关性,随着植物生长所必须的营养元素含量的增加,使得各层片植物能获取的营养更多,多种植物生长状况越好,群落就越复杂,从而逐渐导致优势种的地位越下降,最终群落表现出各层片多个优势种共存的现象。反之,各层片营养物质匮乏,能够适应最低限度的营养物质的物种才能存活并繁衍,各层片的优势种明显,占据着群落分布中的优势地位,因此,较之混交林,马尾松针叶林各层片的结构表现就更为简单,从而导致其群落整体H、D均显著(P<0.05)低于除II外的其他混交林群系。何雅琴等[9]对大练岛的森林群落多样性研究也表明,台湾相思+木麻黄林的群落整体H、D均显著(P<0.05)高于台湾相思林和木麻黄林。表明混交林较于纯林,更能增加群落的生物种类,并提高Shannon-Wiener和Simpson指数的大小。
Pielou指数(E)越高,植物群落结构越稳定[30]。本研究地马尾松常绿针阔混交林群落整体E值显著(P<0.05)高于马尾松常绿针叶林(表7),与大练岛[9]的方差分析比较结果也表明,大练岛的台湾相思林和台湾相思+木麻黄林的群落整体E值显著(P<0.05)高于本研究地的马尾松常绿针叶林,这与群落演替规律相符,群落演替越后期,群落越稳定[31]。Patrick指数(R)直接反映群落物种丰富度[7]。II群系(马尾松+杨桐林)的灌木层丰富度指数R显著(P<0.05)低于其它4类群系,主要原因可能是,II群系的6个样地郁闭度均较大(0.8,0.8,0.85,0.88,0.93和0.93),研究地有41种阳性树种(都为木本植物)和鹅掌柴等不耐荫庇的半阳性树种,鹅掌柴在阳光不足的高郁闭度林下灌木层占比较低,甚至难以存活(图4),而马尾松林树冠较之更窄,更有利于光照进入林下,使得林下植物,特别是阳性木本植物的丰富度更高。与大练岛[9]的方差分析比较结果发现,本研究地马尾松林丰富度指数R显著(P<0.05)高于大练岛的所有森林群落类型(台湾相思林、木麻黄林及2种物种的混交林)。引起这一结果的主要原因在于,马尾松林乔木层和灌木层丰富度指数R均显著(P<0.05)高于大练岛的所有森林群落类型的乔木层和灌木层R值。由于马尾松较于木麻黄和台湾相思,生长高度更高,与其他乔木物种存在较大的高度差,在资源竞争时能形成垂直方向的互补[32],同时其林冠密度更低,更有利于阳性树种的生长,因此其乔木层物种更丰富;而灌木层计算乔木幼树,因此丰富的乔木层物种也一定程度上促进了灌木层物种的丰富度,因此马尾松林乔灌层指数R更高。但马尾松林草本层指数R显著(P<0.05)低于大练岛的所有森林群落类型的草本层R值。其主要原因在于,马尾松林草本层多以芒萁为优势物种,芒萁对杂草密度、种子萌发和幼苗生长的具有化感效应[33—34],严重减少了草本层物种,同时,竹类在马尾松林乔木层、灌木层均分布较广,导致草本层光照严重不足,且竹类的凋落物也降低了草本植物的生存空间[35],因此导致马尾松林草本层R指数最低。I群系(马尾松+鹅掌柴林)的草本层R值均显著(P<0.05)高于本研究的其他群系,II群系的草本层H、D、R均显著(P<0.05)低于其他4类群系,主要原因是II群系所有样方草本层均以芒萁为单一优势种,III—V群系的草本层也以芒萁作为优势种居多,但I群系有一半的样方草本层优势种不是芒萁,以芒萁作为优势种的Q1、J1样方芒萁的重要值不高,没有形成致密层。芒萁在林下形成致密层时会大量截留林冠乔木的凋落物,进而影响其凋落物分解和养分循环过程[36],加之芒萁具有化感效应[33—34],因此抑制了草本层其他物种的生长,物种丰富度也大大降低。同时,这些样方的土壤pH和速效钾较于其他群系样方的更低,土壤pH和速效钾与灌草层的物种多样性指数均呈正相关性,pH过低,植物对大量元素的吸收减弱[37],而钾含量过低,会降低植物酶的激活、蛋白质的合成和养分的运输等[38],从而制约了植物的生长发育。而土壤pH与乔木层的物种多样性呈负相关性,主要原因可能是乔木层的主要物种如马尾松、鹅掌柴、茶竿竹等都喜欢生活在偏酸的土壤中,因此pH越高,越不利于乔木层植物的生长。
4 建议
马尾松混交林相较于马尾松林物种丰富度更高,群落稳定性更强,但相对的,混交林过高的林分郁闭度严重制约了林下灌木层植物的生存环境,物种丰富度明显降低,马尾松、鹅掌柴等建群种的自然更新受到严重制约,不利于群落的长期稳定发展。因此未来马尾松森林经营中要结合考虑这两个问题,通过人为抚育间伐,促进马尾松林向混交林转换,同时控制林窗面积,增大林内的辐射强度,也通过补植、控制土壤pH和速效钾的含量等,促进群落长期稳定发展。
研究地中已见1级恶性入侵植物钻叶紫菀和小蓬草2种,且在样线踏查时,发现岛上也分布有其他1级恶性入侵植物,如假臭草(Praxelisclematidea)、鬼针草(Bidenspilosa)、马缨丹(Lantanacamara)等。研究地草本层以芒萁为最高优势种,芒萁的化感效应不仅抑制了恶性入侵植物的入侵,也抑制了草本层一些植物的生长,导致大部分样方草本层多样性指数最低。且芒萁单优草本层片虽然广泛分布于亚热带低山丘陵区的多种退化群落,但消退于常绿阔叶森林群落。若芒萁消退,对入侵植物的化感作用也将减少,而1级恶性入侵植物的入侵性极强,会严重威胁本土植物的生态位,因此需要通过人为干预手段控制林下芒萁密度,既能增加草本层物种丰富度,也能有效抑制入侵植物的扩散。
研究地分布有国家一级保护植物野生建兰(Cymbidiumensifolium)、国家二级保护植物野生茶(Camelliasinensis)和花榈木(Ormosiahenryi),在对研究地进行人为抚育时,应注意加强对这些植物的保护力度,防止人为采摘、毁坏,必要时应采取圈地、引种等保护手段。
猜你喜欢
发明与创新·中学生(2024年6期)2024-05-29 00:00:00
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
林业勘查设计(2022年1期)2022-02-15 05:35:12
意林(2021年19期)2021-10-27 14:22:57
安顺学院学报(2021年4期)2021-09-16 08:11:08
食品安全导刊(2021年21期)2021-08-30 08:22:08
少儿美术(2020年8期)2020-11-06 05:21:38
林业调查规划(2020年3期)2020-06-03 07:02:36
故事作文·高年级(2019年8期)2019-08-20 17:53:42
