
土壤碳库积累与分配对热带森林恢复的响应
2023-12-25 14:27解玲玲王邵军王郑钧郭志鹏张昆凤张路路樊宇翔郭晓飞夏佳慧兰梦杰杨胜秋
生态学报 2023年23期
解玲玲,王邵军,肖 博,王郑钧,郭志鹏,张昆凤,张路路,樊宇翔,郭晓飞,罗 双,李 瑞,夏佳慧,兰梦杰,杨胜秋
西南林业大学生态与环境学院,昆明 650224
土壤有机碳库作为陆地生态系统中最大的碳储藏库,约占总碳储量的三分之二[1—2],其微小变化导致大气CO2浓度的显著变化[3—4]。土壤微生物量碳库、易氧化有机碳库、惰性有机碳库等有机碳组分,主要来源于动植物残体、腐殖质及微生物体等[5],其在一定程度上决定土壤总有机碳的积累、分配及分解过程,而且能调控土壤腐殖质和团聚体结构的形成、土壤供水供肥能力以及有机物质的分解、转化与循环过程[6]。微生物量碳对土壤有机碳库的贡献仅占1%—4%,但能通过参与土壤生物化学转换与养分循环,调控土壤养分的固定、转化、分解及稳定过程[7]。易氧化有机碳影响土壤有机碳库的时效性和有效性,对外界环境变化反应十分敏感,通常作为表征土壤有机碳库初期变化的主控因子[8],惰性有机碳是土壤有机碳库中相对难分解、较稳定的部分,是土壤对环境长期变化的响应指标[9]。因此,探索土壤总有机碳库与微生物量碳库、易氧化有机碳库、惰性有机碳库组分之间相互关系及其积累与分配特征,是研究生态系统碳过程及全球变化的关键科学问题。
研究表明,土壤有机碳库及其各组分积累与分配动态,取决于气候类型、土地利用变化、土壤环境条件、人类活动干扰等生物与非生物因素的作用[10—14]。其中,森林恢复是促进生态系统修复、土壤有机碳库蓄积与分配的重要驱动因素[15]。随着森林恢复进行,植被覆盖度、植物多样性、地上-地下凋落物输入逐渐增加,促进了土壤微生物及动物主导的生态学过程,从而可能影响土壤有机碳库及各组分的蓄积与分配[16]。张金[17]研究表明,随森林恢复年限增加,土壤有机碳储量呈上升趋势,王卫霞[18]发现重火干扰后北方温带森林恢复过程中土壤有机碳储量表现为后期阶段显著高于前中期阶段。特别是,土壤微生物碳源主要来自于活微生物、死微生物、微生物代谢产物,在恢复进程中,逐渐趋于稳定的森林小气候促进了土壤微生物量碳的蓄积[19],而受凋落物碳输入以及微生物固定、转化等因素的调控,森林恢复进一步促进土壤易氧化有机碳、惰性有机碳含量的积累[20]。然而,目前国内外学者主要集中于亚热带及温带森林土壤有机碳库的时空变化特征研究[21],而关于热带森林恢复过程中土壤碳库组分积蓄与分配动态的研究却相对缺乏。因此,探明土壤有机碳库组分积累与分配对热带森林恢复的响应机制,能为理解热带森林土壤碳沉降过程的调控机制、温室效应减缓和全球碳平衡维持提供关键数据支撑。
西双版纳位于我国大陆热带雨林面积分布最为集中的气候区,由于人口增加、刀耕火种农业活动等人为因素干扰,热带雨林生态系统遭到了严重破坏,形成了一系列处于不同恢复阶段的次生热带森林类型[22]。随着热带森林恢复进程,植物丰富度与多样性、凋落物输入及土壤理化环境可能发生显著改变,进而导致土壤总有机碳储量及微生物量碳库、易氧化有机碳库、惰性有机碳库积累量的变化[15]。本文以西双版纳3种恢复阶段的森林类型(白背桐群落、崖豆藤群落、高檐蒲桃群落)为研究对象,比较不同恢复阶段热带森林土壤碳库各组分积累与分配(微生物量碳/总有机碳储量、易氧化有机碳/总有机碳储量、惰性有机碳/总有机碳储量)的时空特征,揭示土壤总有机碳与碳组分的相互关系,并分析热带森林恢复过程土壤碳库组分积累/分配与主要环境因子(植被丰富度与多样性、土壤温湿度、容重、pH、全氮、水解氮、铵氮、硝氮等)的关系,旨在探明土壤有机库积累与分配对热带森林植被恢复的响应特征,为理解热带森林恢复过程中土壤碳库变化对全球碳平衡的贡献提供基础数据参考。
1 材料与方法
1.1 样地概况
研究区位于中国科学院西双版纳热带植物园(101°16′E,21°55′N),地处东南亚热带北缘,为季风气候区,年平均气温21.5℃,年平均降水量1557 mm,有明显的干湿交替,其中雨季(5—10月)高温多雨,降水量占全年的87%,干季(11—4月)降水主要以浓雾的方式,仅占全年降水量的13%。土壤质地多为壤土,由花岗岩母质风化所形成的砖红壤,呈酸性。地带性植被类型为热带雨林和季雨林,刀耕火种恢复形成了白背桐、中平树、崖豆藤、野芭蕉、高檐蒲桃等一系列群落类型。
在西双版纳中国科学院热带森林植物园实验区内,选择具有代表性的3个处于不同恢复阶段热带森林(白背桐群落、崖豆藤群落、高檐蒲桃群落)为研究样地,3个群落处于连续演替的不同恢复阶段且起源于热带雨林经刀耕火种后形成的裸地,立地条件(包括坡度、坡向与坡位等)及土壤理化性质(pH= 4.5,总有机碳= 36.7 g/kg、全氮= 1.9 g/kg、全磷= 0.5 g/kg)等基本类似,样地之间间隔100—800 m左右(表1)。采用标准地法调查3个恢复阶段植物群落多样性,在群落种类组成分布比较均匀的地段,分别随机选取5个大小为40 m×40 m样方(距离大于200 m),并在各样方内分别随机设置5个灌木(4 m×4 m)和草本(1 m×1 m)样方,测算各样地乔木层及林下植物Margalef丰富度(MR)、Shannon多样性(H′)。其中,MR和H′分别按下式计算[16]:
MR=(S-1)/lnN
(1)
H′=-∑(pi/lnpi)
(2)
式中,S为物种总数;N是群落中所有类群的个体总数,式(2)中pi=ni/N;ni为第i个类群的个体数。
1.2 样品采集与指标测定
于2018年3、6、9、12月在上述样地按三点法分别随机选择3个重复样方,去除地表凋落物后按照三个土层0—5、5—10、10—15 cm分层破坏性采集土壤样品,采用便携式土壤水分温度测量仪(SIN-TN8)同步测定各土层的土壤温度和含水量。将样品带回实验室,剔除树根、残体等杂质后在室内进行自然风干,样品风干后过2 mm筛备用。
土壤有机碳 (SOC)采用油浴加热-重铬酸钾氧化法测定[23];微生物量碳 (MBC)采用液态氯仿熏蒸浸提-水浴法测定[24];易氧化有机碳 (EOC)采用高锰酸钾氧化法测定[25];土壤容重 (BD)采用环刀法测定[26]。参照鲁如坤《土壤农业化学分析方法》[27]测定土壤pH值、全氮 (TN)、水解氮 (HN)、铵氮 (AN)、硝氮 (NN)等土壤指标。
土壤剖面总有机碳储量Cs、土壤微生物量碳储量、易氧化有机碳储量用下列公式计算[28]
Cs=C×D×E×(1-G)/10
(3)
式中,Cs为碳储量、C分别代入为土壤有机碳含量(g/kg)、土壤微生物量碳(g/kg)、易氧化有机碳(g/kg),D为土壤容重(g/cm3),E为土层厚度(cm),G为直径>2mm的石砾所占的体积比例(%)(调查发现土壤粒径均<2 mm,故忽略不计)。代入计算分别得出总有机碳储量(CSOC)、土壤微生物量碳储量(CMBC)、易氧化有机碳储量(CEOC)。惰性有机碳储量(CROC)通过CSOC-CEOC-CMBC得出。微生物量碳、易氧化碳、惰性有机碳占土壤总有机碳储量的比例(MBC%、EOC%和ROC%)分别通过CMBC/CSOC、CEOC/CSOC、CROC/CSOC计算得出。
1.3 数据处理与分析
使用Excel 2010制作不同恢复阶段样地情况、土壤理化性质变化表,土壤有机碳库组分间相互关系采用多项式回归曲线图;单因素ANOVA检验通过SPSS 26.0比较各样地变量之间的差异显著性;采用Origin 2021制作土壤碳库积累与分配时空特征图;运用Mental检测植物及土壤性质对土壤碳库积累/分配的相关性及解释率;通过Canoco 5软件进行冗余分析(RDA)分析,确定影响土壤碳库积累与分配的主要环境因子。
2 结果与分析
2.1 不同恢复阶段热带森林土壤碳库组分积累的时空特征
西双版纳热带森林恢复显著促进了土壤有机碳的积累(图1—4,P<0.05)。CSOC、CMBC、CEOC、CROC大小顺序为:恢复后期高檐蒲桃群落(37.60 t/hm2、1.69 t/hm2、12.46 t/hm2、23.51 t/hm2)>恢复中期崖豆藤群落(34.12 t/hm2、1.42 t/hm2、9.96 t/hm2、22.43 t/hm2)>恢复前期白背桐群落(30.71 t/hm2、1.12 t/hm2、8.65 t/hm2、20.53 t/hm2),相较于恢复前期CSOC、CMBC、CEOC、CROC在恢复后期分别提高了20.54%、50.84%、44.11%、14.51%,恢复中期分别提高了11.12%、25.99%、15.12%、9.25%。不同恢复阶段CSOC、CMBC、CEOC、CROC均随时间变化显著,月动态呈“单峰型”变化,6月达到峰值,12月达到最低水平,总体呈现出湿季(6,9月)大于干季(3,12月)的季节变化特征。恢复前期,6月份CSOC、CMBC、CEOC、CROC比12月分别增加了26.02%、102.78%、139.37%、19.70%;恢复中期增加了37.98%、78.85%、107.09%、21.77%;恢复后期增加了42.26%、80.65%、122.12%、24.86%。
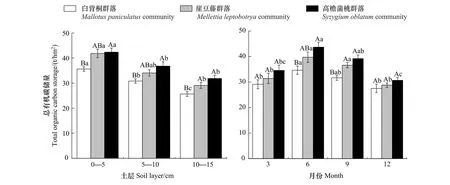
图1 不同恢复阶段总有机碳储量时空变化

图2 不同恢复阶段微生物量碳储量时空变化
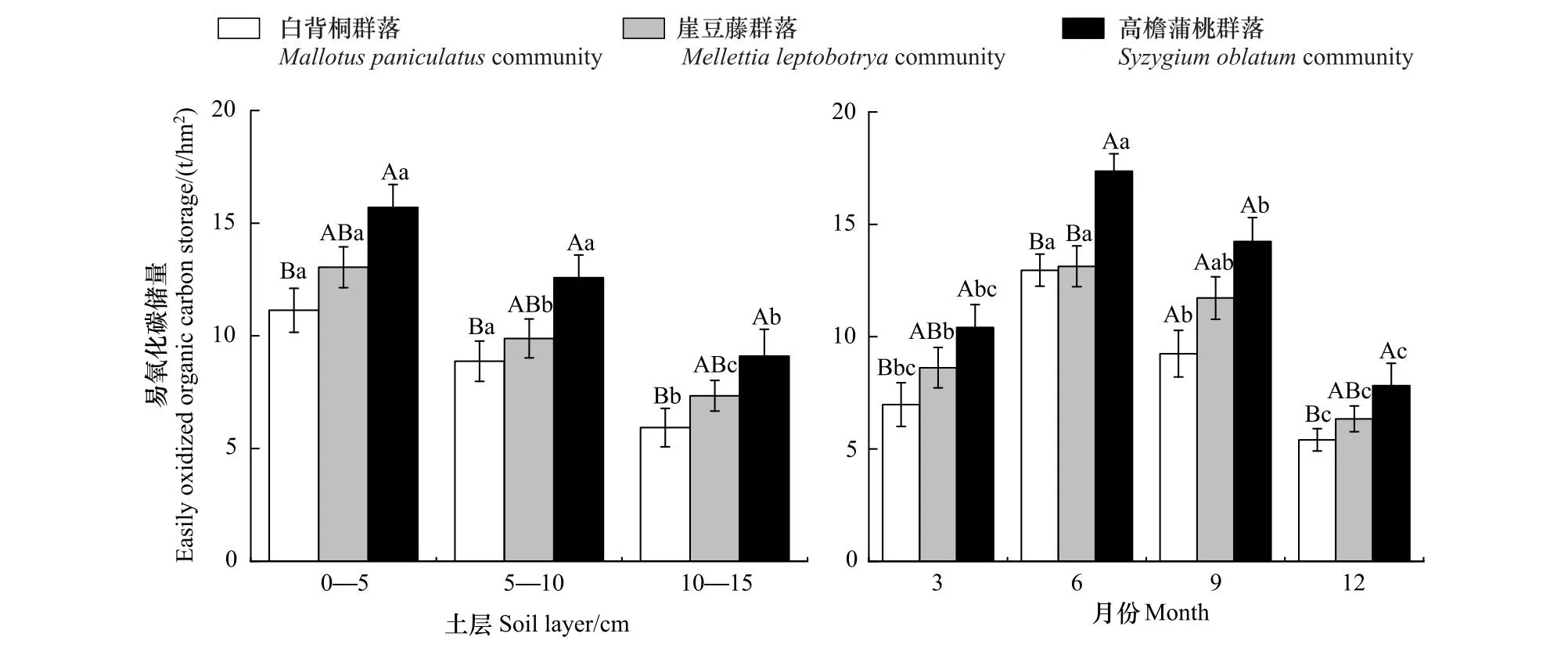
图3 不同恢复阶段易氧化碳储量时空变化

图4 不同恢复阶段惰性有机碳储量时空变化
热带森林恢复CSOC、CMBC、CEOC、CROC均具有随土层加深而减少的垂直变化特征(图1—4,P<0.05)。在恢复前期,0—5 cm土层CSOC、CMBC、CEOC、CROC比5—10 cm土层分别增加了1.15、1.44、1.25、1.14倍,比10—15 cm土层分别增加了1.38、1.82、1.87、1.28倍;在恢复中期,0—5 cm土层CSOC、CMBC、CEOC、CROC比5—10 cm土层分别增加了1.22、1.33、1.32、1.15倍,比10—15 cm土层分别增加了1.42、1.95、1.77、1.32倍;在恢复后期,0—5 cm土层CSOC、CMBC、CEOC、CROC比5—10 cm土层分别增加了1.15、1.24、1.25、1.13倍,比10—15 cm土层分别增加了1.32、1.74、1.72、1.30倍。CSOC、CMBC、CEOC、CROC在每一土层中均呈现出恢复后期>中期>前期的变化趋势,可见森林恢复及土层对土壤碳库组分影响显著。
2.2 不同恢复阶段热带森林土壤碳库组分分配的时空特征
热带森林恢复显著提升了MBC%、EOC%(图5—6,P<0.05),表现为后期(4.47%、32.77%)分别是前期(3.58%、26.14%)、中期(4.08%、30.07%)的1.14、1.25,1.15、1.25倍;不同恢复阶段ROC%均值分别为前期(71.83%)、中期(70.10%)、后期(68.69%),热带森林恢复对ROC%的影响不显著(图7,P>0.05)。土壤有机碳组分分配具有明显的时间变化特征(图5—7,P<0.05),MBC%和EOC%在6月达到最大值(2.43%—5.07%),12月达到最小值(18.40%—39%)。恢复前期,12月份MBC%、EOC%比6月分别减少了41.46%、43.75%;恢复中期减少了30.43%、48.64%;恢复后期减少了30.00%、46.15%,MBC%、EOC%在同一月份表现为恢复后期>恢复中期>恢复前期,湿季(6、9月)分配大于干季(3、12月)的变化特征。ROC%表现出相反的特征,在12月达到峰值(58.61%—70.37%),6月达到谷值,在恢复前期、中期、后期6月比12月分别减少了9.12%、11.73%、14.83%。

图5 不同恢复阶段微生物生物量碳分配时空变化

图6 不同恢复阶段易氧化有机碳分配时空变化
热带森林不同恢复阶段MBC%、EOC%具有随土层深度增加而减少的垂直变化特征(图5—7,P<0.05)。MBC%在恢复前期、中期、后期0—5 cm分别是5—10 cm、10—15 cm的1.26、1.32倍,1.16、1.31倍,1.06、1.31倍;EOC%在恢复前期、中期、后期0—5 cm分别是5—10 cm、10—15 cm的1.10、1.37倍,1.17、1.35倍,1.08、1.33倍。ROC%具有随土层深度呈增加的垂直变化趋势,但无显著差异,ROC%在恢复前期、中期、后期0—5 cm分别是5—10 cm、10—15 cm的0.97、0.91倍,0.96、0.93倍,0.97,0.96倍。
2.3 土壤总有机碳与各碳组分储量的相互关系
通过对CSOC、CMBC、CEOC、CROC进行多项式回归分析,结果表明CSOC随CMBC、CEOC、CROC的增加而显著增加(图8,P<0.05)。土壤有机碳组分对总有机碳储量变化的解释率不同,表现为CMBC(83.71%)>CROC(82.17%)>CEOC(78.54%)。

图7 不同恢复阶段惰性有机碳分配时空变化

图8 土壤总有机碳储量与碳组分储量相互关系
2.4 土壤有机碳积累/分配与植物、土壤环境因子之间的关系
随着热带森林恢复的进行,乔木物种丰富度(TSR)、林下物种丰富度(USR)、乔木Shannon多样性(TSD)、林下植物Shannon 多样性(USD)、HN、AN均显著呈现出恢复后期>恢复中期>和恢复前期的变化趋势(表2),在恢复后期,TSR、USR、TSD、USD、HN、AN分别比恢复前期提高了490.82%、240.92%、42.78%、50.04%、18.94%、20.18%;TN、NN呈现出恢复后期显著大于恢复前期、中期的变化趋势(表2),相较于恢复前期,TN、NN在恢复后期分别提高了12.73%、25.51%;ST、BD、pH并无显著变化。
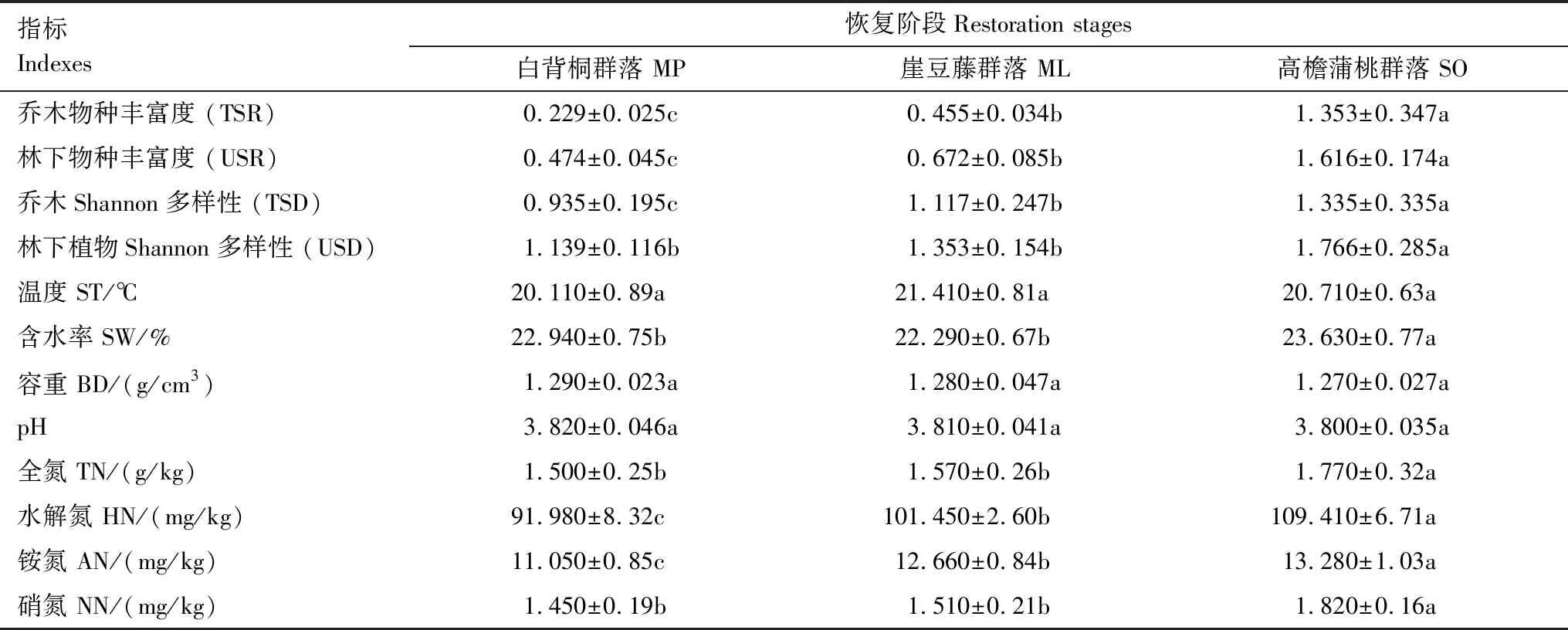
表2 不同恢复阶段植物多样性及土壤理化性质的比较
通过Mantel分析得出土壤有机碳库积累(CSOC、CMBC、CEOC、CROC)/组分分配(MBC%、EOC%、ROC%)与环境因子(TSR、USR、TSD、USD、ST、SW、BD、pH、TN、HN、AN、NN)之间的相关性和解释值(图9)。结果表明,TN均与土壤有机碳库积累(CSOC、CMBC、CEOC、CROC)与分配(MBC%、EOC%、ROC%)弱相关(Pearson′sr=0.4—0.6;P<0.05);土壤有机碳库积累与TSR、USR、BD、pH、HN、NN中等相关,土壤有机碳库分配与SW、AN、NN中等相关(Pearson′sr=0.6—0.7;P<0.05);土壤有机碳库积累与TSD、USD、SW、ST、AN较强相关,土壤有机碳库分配与TSR、USR、TSD、USD、ST、BD、HN较强相关(Pearson′s=0.7—0.9;P<0.05)。pH、AN是土壤有机碳库积累的主要影响因子,而TN、HN对土壤有机碳库分配具有重要影响(Mantel′sr≥50%,Mantel′sP<0.05)。

图9 土壤有机碳积累/分配与植物、土壤环境因子Mantel分析
RDA结果显示(图10):第一、二坐标轴对土壤有机碳库积累(CSOC、CMBC、CEOC、CROC)贡献了91.59%,对土壤有机碳库分配(MBC%、EOC%、ROC%)贡献了82.42%。SW、ST均与CSOC和CMBC的夹角较小,是CSOC和CMBC主控因子;HN、USR则分别是CEOC和CROC的主要影响因子;TSD与MBC%夹角较小,SW与EOC%夹角较小,故TSD、SW是MBC%和EOC%的主要驱动因子;BD与夹角ROC%较小,是ROC分配变化的主控因子。

图10 土壤有机碳积累/分配与植物、土壤环境因子冗余分析
3 讨论
3.1 热带森林不同恢复阶段土壤碳库组分积累时空变化特征
本研究发现热带森林恢复显著促进土壤有机碳库各组分的积累。这是热带森林恢复能够引起植物覆盖、凋落物输入和植物多样性增加,从而导致土壤有机碳库各组分储量均呈增加的趋势[29]。在恢复初期,植被群落组成、结构及多样性较为单一,森林内小气候不稳定,且凋落物数量较少,输入土壤的养分低,不利于土壤微生物对土壤微生物量碳、易氧化有机碳和惰性有机碳的转化和固持[30—31]。在恢复中后期,森林小气候逐渐趋于稳定、群落结构与物种组成趋于复杂,生物量及凋落物积累增加,促进了土壤有机碳库组分的蓄积。徐红伟[32]亦研究表明黄土高原地区森林恢复(8a—45a)促进土壤有机碳的积累,但其增幅(66.67%)大于西双版纳热带森林(18.91%),可能是热带高温高湿的水热条件刺激土壤微生物对有机碳的分解,致使土壤碳组分积累量较小[33]
热带森林恢复对土壤有机碳库组分积累季节动态的影响表现为湿季显著高于干季。这与样地土壤温湿度变化呈现出一致规律,说明土壤微气候对土壤碳蓄积具有显著影响。湿季中,6月土壤有机碳库组分积累量最高,可能由于湿季较高的土壤温湿度有利于微生物的碳固持。曹润[7]和李少辉[34]研究表明,西双版纳湿季适宜温度与充沛降水,使得植被新陈代谢旺盛,促进了枯枝落叶和根系分泌物向土壤的碳输入,增强了土壤微生物活性从而促进土壤有机碳的积累。干季低温少雨,植被生长及凋落物输入速率缓慢,土壤微生物碳固持及碳库组分积累均较慢,从而呈现出土壤碳库组分积累湿季大于干季的变化规律。热带森林恢复对土壤有机碳积累的季节波动具有样地差异性,恢复后期土壤有机碳储量、微生物量碳储量、惰性有机碳储量的季节波动大于恢复前期,此现象可能与立地条件、土壤有机碳矿化速率等因素有关[35]
不同恢复阶段热带森林土壤有机碳库组分积累的垂直变化表现为沿土层深度的增加而减小。土壤有机碳储量主要取决于植物根系、分泌物、微生物数量,由于凋落物的碳归还量随土层加深而降低导致了土壤有机碳的表聚现象[36]。表层凋落物输入数量较多,水热条件与透气性也较高,能够促进土壤微生物对表层有机碳的固持[37—39]。另外,陈小梅[40]研究得出的南亚热带森林土壤有机碳库积累的垂直减少幅度(46—55%)与西双版纳热带森林(15—87%)具有一定的差异性,这与不同研究区样地的林龄、植被多样性、凋落物输入及土壤理化环境的差异密切相关[41]。热带森林恢复对土壤有机碳积累的垂直变化幅度有所不同,土壤易氧化有机碳储量在恢复前期的垂直变化幅度大于恢复中后期,猜测可能与土壤表层输入下层的土壤有机碳质量等因素相关[42]。
3.2 热带森林不同恢复阶段土壤碳库组分分配变化特征
热带森林恢复促进了土壤微生物量碳和易氧化有机碳的分配,但对惰性有机碳分配无显著影响。这由于在恢复初期,通过地上植被进入土壤中的有机碳较少,导致恢复初期微生物量碳及易氧化碳的分配较低。而随着热带森林的恢复,植被组成逐渐稳定且多样化,凋落物输入量、根系分泌物及细根量增加,微生物活动增强,加快了土壤有机碳的周转速率,导致活性有机碳的分配量增加。而土壤惰性碳组分是由活性有机碳通过微生物转变碳形成,研究表明,森林恢复初期土壤惰性碳组分可能不会产生显著的增加,因此,本研究经51年的恢复未能导致惰性有机碳分配比例的显著增加。一般到60—100年后,活性有机碳在微生物作用下大量转化为惰性有机碳,从而使得惰性碳储量及惰性碳分配显著提高[6]。热带森林不同恢复阶段土壤碳库组分分配(MBC%、EOC%)表现为湿季显著高于干季,且均在恢复后期配比最高,而ROC%呈现出相反的变化规律。Sinsabaugh[43]研究结果亦表明,恢复后期稳定的微环境和较高的凋落物输入,导致微生物对易分解有机碳的转化与积累量大于其分解量,从而促进了微生物量碳及易氧化有机碳在总有机碳库中占比的增加,相应地导致了惰性有机碳占比的减少。习丹[44]研究表明,湿季高温高湿样地环境及丰富碳源,有利于土壤微生物对活性有机碳的固持作用,并相应地减少了惰性有机碳,使其在总有机碳库中的占比降低;干冷的环境不利于微生物的活性,可能降低其对土壤有机碳的积累、分解与转化,影响土壤易氧化有机碳的产生与积累,从而使热带森林干季微生物量碳、易氧化有机碳在土壤碳库的分配比湿季少,干季干冷的土壤环境抑制了微生物对惰性有机碳的分解,导致其配比湿季高。与本研究结果不同,陈小梅[40]研究表明南亚热带恢复演替序列EOC%呈现恢复早期高于后期的变化趋势,可能由于不同气候区样地物理化学环境的差异而导致。
热带森林不同恢复阶段MBC%、EOC%呈现随土层深度的增加而减少的垂直变化特征,而ROC%随土层无显著变化。Sinsabaugh[45]、韩琳[46]等研究表明,表层凋落物层和根系的集聚,能够促进土壤微生物的生长、繁殖与活动,而深层土壤透气差、温度低、可利用的反应底物少,土壤微生物活性较弱,导致土壤微生物的碳固定能力较弱,降低了土壤易氧化有机碳的分配,同时易氧化有机碳向惰性有机碳的转化在不同土层中可能差异不显著,从而导致惰性有机碳配比无显著垂直变化[47]。另外,本研究中恢复前期MBC%、EOC%沿土层减少幅度高于恢复中后期,由于恢复前期还未形成稳定的林内小气候,植物所产生的碳源较少且更多在维持自生所需的生长发育,导致分配到下层的较少,从而减少最高。
3.3 植物与土壤环境因子对碳库储量及组分分配的影响
热带森林恢复显著提高了乔木与林下植物物种丰富度、Shannon多样性以及土壤氮库含量,从而影响土壤碳库储量及组分分配。胡靓达等[48]研究表明,植物群落作为土壤碳积累的主要驱动力,其类型、组成、结构、起源的不同决定了植物生物量和凋落物输入量的差异性,并影响土壤微生物对有机碳的固持,森林恢复过程中植物丰富度、多样性及凋落物数量的显著增加,能够促进土壤碳的蓄积过程[49—51]。热带森林恢复提高了植被丰富度及多样性,促进有机碳输入、土壤微生物丰富度及功能多样性的增加,提高了对碳库的分解与转化,刺激易氧化有机碳形成与积累,从而显著促进了土壤有机碳库对微生物量碳及易氧化碳的分配[52—53]。惰性有机碳主要存在于土壤黏粒及细粉粒中,具有较强的稳定性,一般在土壤中不易被微生物分解或矿化,大多数平均周转年限超过一百年,所以热带森林恢复对惰性有机碳分配的影响并不显著[54]。
土壤氮库组分(全氮、水解氮、铵氮、硝氮)显著促进了热带森林土壤碳库储量及组分分配,而对ROC%的影响不显著。李昌珍[55]研究得出土壤氮库含量主要通过影响植被生长、枯枝落叶的生产及凋落物的质量,而调控土壤碳库各组分的积累与分配。Huang等研究[56]得出植物多样性及丰富度将导致枯落物碳氮比(C/N)降低,枯落物作为土壤-植物营养物质传输的主要桥梁,较高的凋落物质量(低C/N),能提高枯落物的周转速率,有利于微生物对有机碳的分解、转化与积累,同时促进有机质向深层土壤运输的能力[57—59];氮的增加能够提高微生物量和土壤酶活性,提高土壤氮素利用率、抑制土壤中有害物质的产生,为微生物营造良好生长环境,促进土壤有机碳库积累及组分分配[54]。在本研究中恢复后期C/N(20.01)小于恢复中期(24.03)和前期(25.22),恢复前期,植被丰富度、多样性低,C/N较高,土壤微生物活性弱,对凋落物及根系残留物的分解转换速率较慢,易氧化碳的含量较低,导致有机碳库对微生物量碳及易氧化碳的分配低,而恢复后期,植被组成逐渐稳定、丰富度、多样性显著增加,C/N逐渐降低,刺激了土壤微生物的生长、繁殖活动,进而促进了土壤碳库对微生物量碳及易氧化碳分配[60]。
本研究中,土壤容重、pH对热带森林土壤碳库储量及组分分配具有负向调控作用,对ROC%具有正向调控作用。这与贾御夫[60]、张静静等[61]研究结果相似。一般而言,土壤容重小,肥力高、透气性好、土质疏松,有利于水分渗蓄,而随径流冲刷的碳氮养分为植被根系的生长发育提供了养分,进一步促进了植被的生长发育,有利于土壤微生物的生长繁殖,从而提高土壤有机碳库的积累及分配[62]。土壤pH过高过低都会影响土壤性质及养分有效性,抑制植物生长发育和土壤微生物活动,进而影响土壤碳库积累及组分的分配过程[63]。在恢复初期,土壤容重相对较高,其黏粒含量高,土体较为紧实,土壤水热循环水平低,有机质含量低,微生物活性低,不利于易氧化有机碳的积累及惰性有机碳的分解,所以初期微生物量碳及易氧化碳分配低,而惰性碳分配高。随着热带森林恢复的进程,森林的生长发育,为土壤提供了大量有机质,土壤容重逐渐降低,提高了土壤保肥能力,为大量微生物的生长繁殖提供了保障,土壤微生物量的积累促进稳态碳转化为活性碳,即促进了惰性有机碳的分解及易氧化有机碳的形成,致使土壤有机碳库对微生物量碳及易氧化碳分配的提高,对惰性有机碳分配的降低。
4 结论
热带森林次生恢复进程,通过引起林下物种丰富度和乔木香农多样性的增加以及土壤物理环境(温湿度、容重)及氮库(水解氮)含量的变化,从而刺激热带土壤有机碳库组分积累与分配。研究结果有助于理解热带森林恢复过程中土壤有机碳积累与分配特征及其调控机制,能为正确评估热带森林恢复中土壤碳库变化在全球碳平衡和温室效应缓解中的地位与作用提供关键数据参考。
致谢:感谢中国科学院西双版纳热带植物园园林园艺中心及杨效东研究员、周文君副研究员、李少辉博士生为该研究土壤样品采集和测定提供的帮助。
猜你喜欢
核科学与工程(2021年4期)2022-01-12
新世纪智能(数学备考)(2020年9期)2021-01-04
绿色科技(2020年20期)2020-11-20
家庭医学(下半月)(2020年7期)2020-08-24
科学大众(2020年12期)2020-08-13
江苏农业科学(2017年6期)2017-05-11
林业勘查设计(2017年1期)2017-03-11
浙江农业学报(2016年7期)2016-06-15
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
