
贵州习水杜鹃群落结构和种群动态分析
2023-12-21 05:34王逍李崇清穆璁罗康李云波杨瑞
热带亚热带植物学报 2023年6期
王逍, 李崇清, 穆璁, 罗康, 李云波, 杨瑞
(1. 贵州大学林学院,贵阳 550025;2. 贵州习水国家级自然保护区管理局,贵州 习水 564600)
据最新调查结果,中国有37 699 种高等植物,位列全球第3,其中超1.5×104种种子植物为中国特有,约占种子植物总数50%。2021 年,中国科学家首次完成了3.5 万余种野生高等植物的濒危状况评估, 发现40 物种已经灭绝、野外灭绝或者地区灭绝,受威胁物种达3 879 种,占10.84%,其中不乏大量的特有植物[1],这些特有植物在生物多样性评估中占有重要地位[2–5]。但多数的特有植物由于数量少、分布狭窄等原因,往往得不到关注[2],我国有杜鹃花属特有植物450 种[1],这些特有杜鹃花除了带来生物多样性贡献外,还具有极高的观赏价值、文化价值和药用价值。如杜鹃花可作为抗氧化剂、免疫调节剂、抗炎剂和活力恢复剂[6–9];从杜鹃花中提取的天然色素能够代替合成色素,避免了合成色素的副作用[10];食用杜鹃花有清热功能,有益身体健康,且具传统风味[11]。研究还表明在杜鹃花科植物生长的群落中,杜鹃花有助于预测其相关生态系统对人为因素影响的抵抗能力和恢复能力,包括人为导致的气候变化、氮富集和物种入侵等。这种基础物种随着生态系统状态变化出现的动态,将帮助生态学家把重点放在生态系统过程和基础物种响应,而不是历史上占优势的物种和生态系统状态变化的恢复策略[12]。
群落结构的研究有助于掌握自然植被的组成规律,并应用这些规律来指导林业实践活动[13]。种群动态研究不仅能揭示种群的生存现状和演替规律、阐明种群经历的干扰情况、对环境条件的适应能力,还能有效预测种群未来的发展趋势,为濒危物种的保护和管理指明方向[14–15]。植物种群动态是研究生存在特定环境空间里的植物种群随时间呈现的个体数量消长和分布变迁规律[16–17]。
习水杜鹃(Rhododendronxishuiense)系杜鹃花科(Ericaceae)杜鹃花属植物,2010 年开展综合科考时在杨大老林山顶发现的新物种,目前仅分布于贵州习水国家级自然保护区,该种近似于猫儿山杜鹃(R.maoerense),但其花冠呈漏斗形,子房不被短柄腺体,叶较小[18]。习水杜鹃被发现已逾10 年,但未见进一步的调查研究,调查分析该种的分布现状及动态,能为保护区管理机构科学合理的保护和利用该种群提供决策参考。
1 材料和方法
1.1 研究区概况
贵州习水国家级自然保护区,位于贵州省遵义市,地跨习水县和赤水市,地理位置为105º50′~106º29′ E,28º07′~28º34′ N,保护区总面积51 911 hm2, 其中核心区面积21 231.31 hm2,缓冲区面积11 303.8 hm2, 试验区面积19 375.89 hm2,是贵州省面积最大的国家级自然保护区。保护区地形以山地为主,主要为白垩系发育的丹霞地貌,年均温在10.3 ℃~ 17.2 ℃, 年日照数1 091~1 246 h,年降水量770~1 661 mm。保护区内的植物有331 科1 001 属2 539 种,主要由壳斗科(Fagaceae)、樟科(Lauraceae)、山茶科(Theaceae)、蔷薇科(Rosaceae)、忍冬科(Caprifoliaceae)和桑科(Moraceae)等种类占优势的常绿阔叶混交林组成,主要森林群落有润楠(Machiluspingii)-鹅耳枥(Carpinus turczaninowii)林、厚皮栲(Lanneacoromandelica)-杜英(Elaeocarpusdecipiens)林、杜英-石栎(Lithocarpus glaber)林、丝栗栲(Castanopsisfargesii)-山矾(Symplocossumuntia)林等。保护区内还分布有红豆杉(Taxuschinensis)、南方红豆杉(T.wallichianavar.mairei)、福建柏(Fokieniahodginsii)、伯乐树(Bretschneiderasinensis)、鹅掌楸(Liriodendronchinense)、桫椤(Alsophilaspinulosa)等国家重点保护树种。
1.2 样地(线)设置及调查
2020 年6 月—2021 年9 月,在询问保护区周边群众与实地踏查分析的基础上,结合保护区实际确定调查区域,设置13 条样线,沿山脚至山顶设置样线,每条样线3~5 km,宽度0.1~0.5 km (由视野决定),2 条样线相邻不得少于2 km (图1),调查习水杜鹃在贵州习水国家级自然保护区的分布情况。选择有习水杜鹃的分布地设置样地开展调查,但由于习水保护区地形特殊,且习水杜鹃现分布地在高海拔的山顶,分布面积狭窄,无法开展大样地调查。对发现地(杨大老岭,图2)设置10 m×30 m 的样带开展调查,将样带分成3 个10 m×10 m 的样方进行乔木统计,12 个5 m×5 m 的样方进行灌木和草本的统计。调查内容包括:样地地理坐标、海拔、坡度、坡向、坡位等地理信息;所有草本植物名称、平均高度、盖度、多度等;所有胸径(DBH)小于3 cm 的灌木植物的名称、株数、平均高度、平均地径、盖度等;所有DBH 大于3 cm 的乔木植物的名称、胸径、树高、枝下高、冠幅等。同时,以西南角为坐标原点,记录样地中所有木本植物的坐标位置。
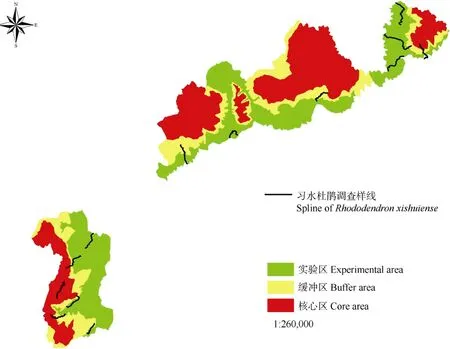
图1 习水杜鹃样线分布图Fig. 1 Sample line distribution of Rhododendron xishuiense in Xishui Nature Reserve

图2 习水杜鹃分布位置示意图Fig. 2 Distribution of Rhododendron xishuiense
1.3 群落结构特征分析
1.3.1 植物地理区系、生活型分析
种子植物和孢子植物地理区系分别参照吴征镒[19]、陈景艳等[20]的划分标准;生活型按照Raunkier 生活型系统统计。
1.3.2 群落物种多样性分析
采用Gleason 丰富度指数、Simpson 多样性指数、Shannon-wiener 多样性指数、Pielou 均匀度指数对群落的物种多样性分析[21–24]。乔木层重要值(I)=(相对密度+相对频度+相对盖度)/3,灌木层重要值(I)=(相对密度+相对频度)/2,草本层重要值(I)= (相对密度+相对频度)/2,Gleason 丰富度指数:R=S/lnA, Simpson多样性指数:, Shannon-Wiener 多样性指数:, Pielou 均匀度指数:J=H/lnS, 式中,Pi为样方中第i个物种的重要值与样地中该层次重要值之和的比值,即相对重要值,S为群落中植物物种数,A为样地面积。
1.4 种群动态
1.4.1 年龄结构和静态生命表编制
采用径阶(地径)代替年龄的方式对乔木杜鹃径级进行划分[25–26],结合调查数据,按0<Ⅰ≤5 cm, 5 cm<Ⅱ≤10 cm,10 cm<Ⅲ≤15 cm,15 cm<Ⅳ≤20 cm,Ⅴ>20 cm 进行径阶划分,将习水杜鹃种群的年龄结构划分为5 个龄级。统计各个龄级内的习水杜鹃株数,按照静态生命表的编制方法编制整个样地的习水杜鹃种群的静态生命表,但静态生命表只是反映了世代重叠的年龄动态历程中的一个特定时间,而不是对同一种群的全部生活史的追踪[27],且考虑到调查中存在误差,在生命表中会出现死亡率为负的情况,因此使用匀滑技术对静态生命表进行处理[28],对Ax进行匀滑修得ax,并据此得到习水杜鹃种群静态生命表。表中参数计算如下:lx=ax/a0×1000,dx=lxlx+1,qx=dx/lx,Lx=(lx/lx+1)/2,,ex=Tx/lx,Kx=lnlxlnlx+1,Sx=lx+1/lx, 式中,Ax为x龄级实际存活数量;ax为修正后x龄级存活数;lx为第x龄级的标准化存活数;lnlx为标准化存活数的自然对数;dx为x至x+1龄级内的标准化死亡数;qx为x至x+1 龄级内的死亡率;Lx为x至x+1 龄级内的平均存活的个体数;Tx为x龄级到超过x龄级存活个体总数;ex为进入x龄级个体的平均期望寿命;Kx为消失率;Sx为存活率。
1.4.2 种群存活曲线、死亡曲线和消失率曲线绘制
以龄级为横坐标,标准化存活个体数的自然对数值(lnlx)为纵坐标,绘制习水杜鹃种群存活曲线,采用指数模型Nx=N0e–bx、幂函数模型Nx=N0x–b对存活曲线进行拟合,若符合指数模型,则为Deevey-Ⅱ型,若符合幂函数模型则为Deevey-Ⅲ型[29]。Nx、N0分别表示x龄级的存活个体数和种群形成初期的个体数,b为死亡率。同样地,以龄级为横坐标,生存函数S(i)、积累死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)等4 个函数值为纵坐标制作习水杜鹃种群的生存率曲线、累计死亡率曲线、死亡密度曲线和危险率曲线。S(i)=S1S2S3……Si,F(i)=1–Si,f(ti)=[Si-1–Si]/hi=Si-1qi/hi, λ(ti)=2(1–Si)/[hi(1+Si)], 式中,Si为种群存活率,qi为种群死亡率,hi为龄级宽度。
1.4.3 种群结构量化动态
采用陈晓德[30]的方法,对习水杜鹃种群结构类型进行量化分析。
种群龄级间动态指数(Vn)为:Vn=(Sn-Sn-1)/max(Sn,Sn-1), 式中,Sn与Sn+1分别为第n和n+1 年龄级种群个体数。
种群年龄结构的数量变化动态指数(Vpi)为:Vpi=,式中,K为最大年龄级。
考虑K和Sn因素的种群结构动态指数(Vˊpi)为:,式中,Vn为种群从n到n+1 龄级间个体数量的变化动态指数,Vpi为忽略外部干扰时整个种群的数量变化动态指数。Sn与Sn+1分别为第n、第n+1 龄级的个体数量,K为种群年龄级数量,max(…)和 min(…)取括号中数列的极大值和极小值。-1≤Vn(或Vpi或Vpi′)≤1,当Vn(或Vpi或Vpi′)取正、零、负值时,表示种群内两相邻龄级间个体数量(或整个种群年龄结构)的增长、稳定、衰退的动态关系。
2 结果和分析
2.1 群落结构特征
2.1.1 习水杜鹃种群分布现状及分布群落物种组成
根据对13 条样线的调查,仅在杨大老岭(105°52′25.941″ E,28°11′30.756″ N,约300 m2)发现习水杜鹃。杨大老岭样地调查有维管植物27科42属65种(表1),其中种子植物24 科39 属62 种,蕨类植物3 科3 属3种。在习水杜鹃群落中,乔木层和灌木层中均以樟科、杜鹃花科、山茶科较多,草本层以禾本科(Poaceae)、百合科(Liliaceae)较多。樟科植物在属的统计上同样在乔木层和灌木层群落中占据优势。从各属物种分析,以杜鹃花属(Rhododendron)、石栎属(Lithocarpus)较多。单科单属植物18 种,占27.69%。群落中有木本植物363 株,其中乔木层以习水杜鹃、岭南石栎(Lithocarpusbrevicaudatus)、桂南木莲(Manglietia chingii)较多。灌木层中杜鹃(Rhododendronsimsii)有179 株,占60.3%,占据绝对数量优势,草本层植物共494 株,箭竹(Fargesiaspathacea) 320 株, 占64.8%,也占据数量上的绝对优势。
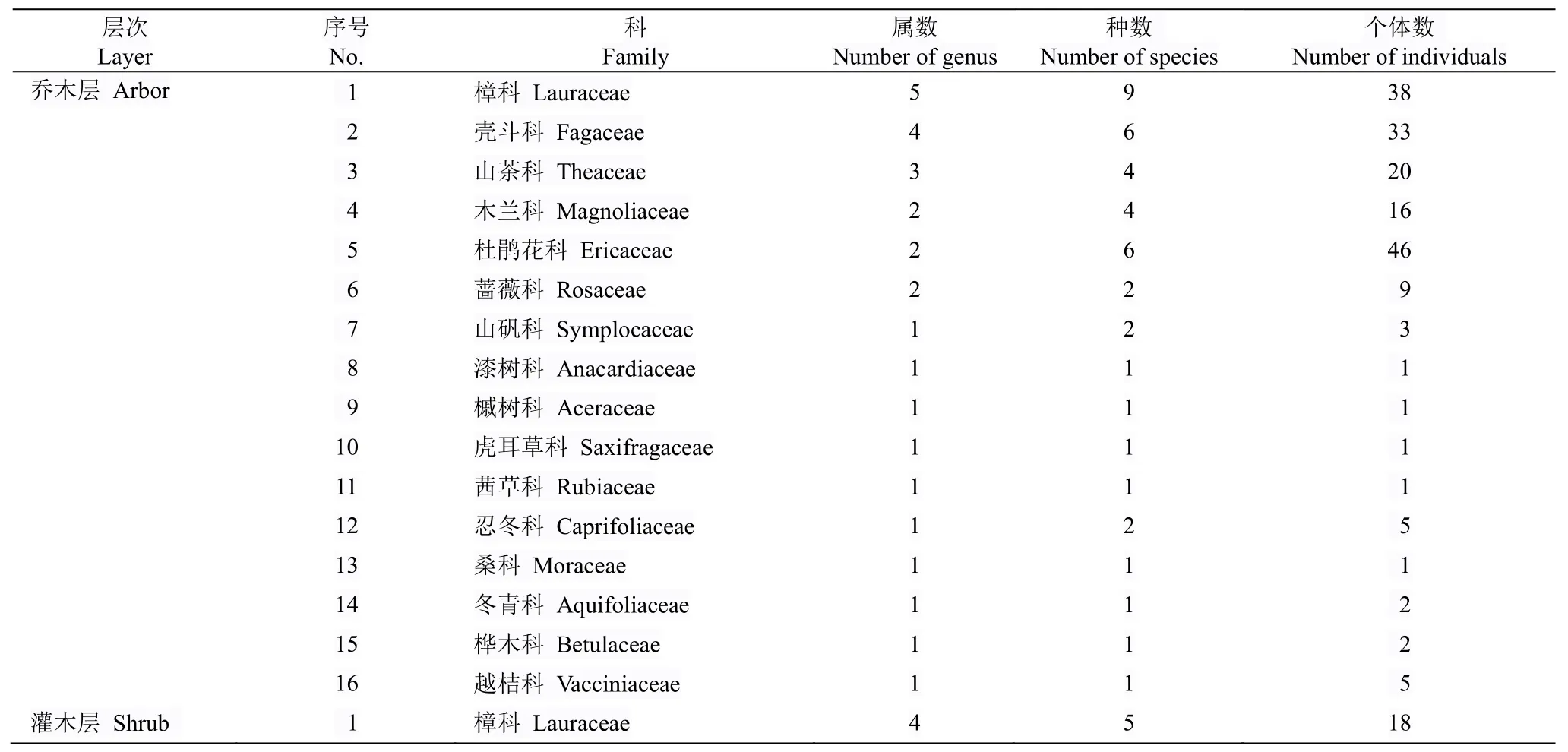
表1 习水杜鹃分布群落的物种组成Table 1 Species composition of Rhododendron xishuiense community
2.1.2 习水杜鹃分布群落的植物地理区系
根据吴征镒对种子植物属的地理分布类型的划分标准,结合陈景艳对贵州蕨类植物地理区系的调查结果,对习水杜鹃群落植物区系进行统计分析(表2)。习水杜鹃群落物种区系分布有14 种分布类型,以泛热带分布、北温带分布、热带亚洲(印度-马来西亚分布)物种数较多。其中泛热带分布有9 科9 属12 种,包含菝葜属(Smilax)、冬青属(Ilex)、厚皮香属(Ternstroemia)、里白属(Hicriopteris)、琼楠属(Beilschmiedia)、求米草属(Oplismenus)、榕属(Ficus)、山矾属(Symplocos)、卫矛属(Euonymus); 北温带分布有6 科6 属11 种,包含杜鹃花属(Rhododendron)、鹅耳枥属(Carpinus)、花楸属(Sorbus)、荚蒾属(Viburnum)、李属(Prunus)、槭属(Acer);热带亚洲(印度-马来西亚分布)有3 科5 属11 种,包含山茶属(Camellia)、含笑属(Michelia)、木莲属(Manglietia) 、 润楠属(Machilus) 和山胡椒属(Lindera)。群落中分布的中国特有植物2 科2 属,分别是香果树属(Emmenopterys)和箭竹属(Fargesia)。热带亚洲至热带非洲分布、热带亚洲和东非或马达加斯加间断分布、热带印度至华南分布、中国-喜马拉雅分布属种均1 种,是习水杜鹃所在群落中地理区系分布最少的类型。
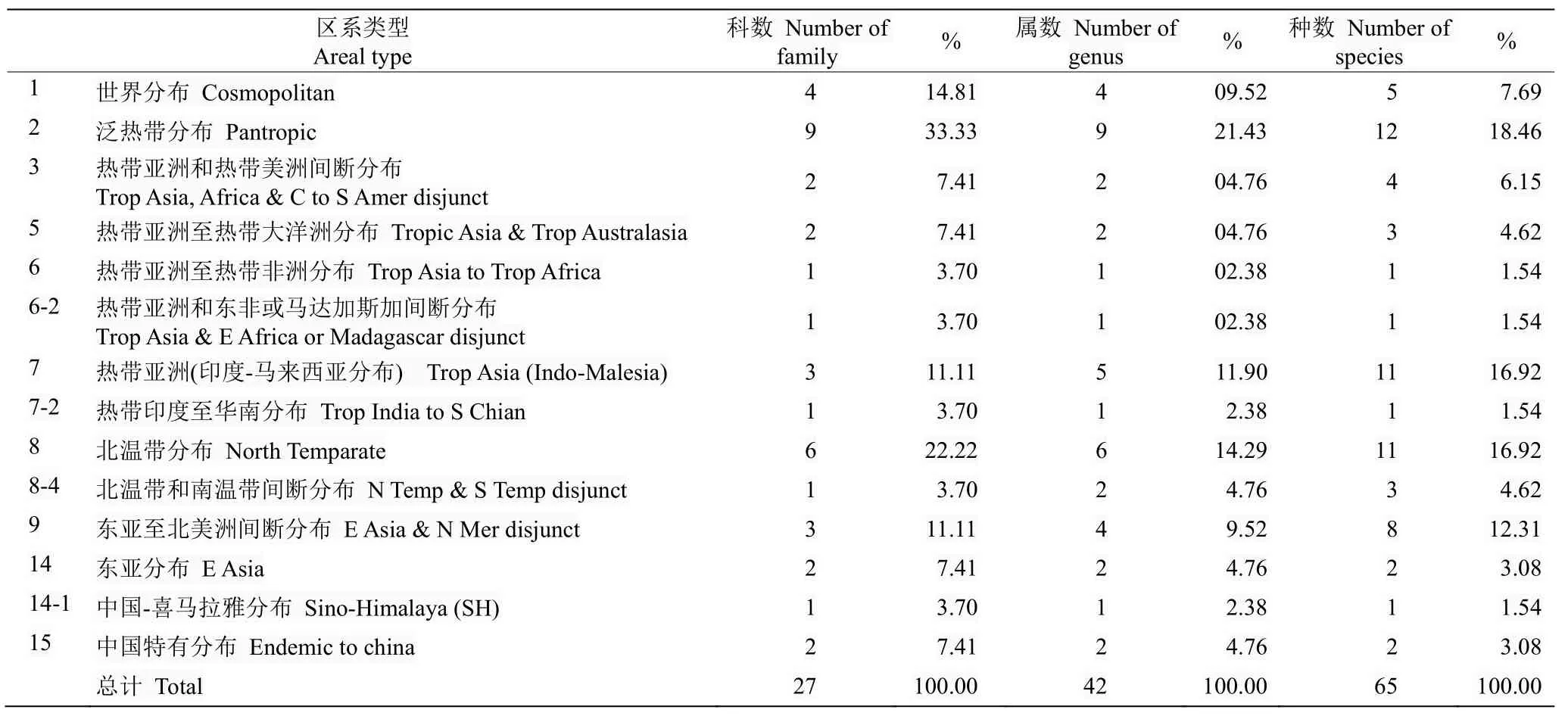
表2 习水杜鹃分布群落植物区系分析Table 2 Areal type of Rhododendron xishuiense community
2.1.3 习水杜鹃分布群落植物生活型
按照Raunkiaer 生活型分类系统,对习水杜鹃群落中65 种植物进行分类,高位芽的物种数最多,为56 种,占总数的86.15%。其中,中型高位芽植物有28 种,占高位芽生活型的50.00%,主要有峨眉含笑(Micheliawilsonii)、厚皮香(Ternstroemia gymnanthera)、桂南木莲、石灰花楸(Sorbusfolgneri)等;小型高位芽植物有26 种,占高位芽生活型的46.43%,主要有习水杜鹃、杜鹃、光枝杜鹃(Rhododendronhaofui)、小果润楠(Machilusmicrocarpa)等;矮高位芽植物有2 种,占高位芽生活型的3.57%。地上芽植物5 种,占总数的7.69%;地面芽植物3 种, 占总数的4.62%;地下芽植物1 种,占总数的1.54%。
2.1.4 习水杜鹃分布群落的优势种和物种多样性
从表3 可见,习水杜鹃分布群落乔木层的物种有43 种,重要值较大的有习水杜鹃、石灰花楸、江南樾桔(Vacciniummandarinorum)、厚皮香、岭南石栎、桂南木莲、小果润楠、石栎、硬壳柯(Lithocarpushancei)、长蕊杜鹃(Rhododendronstamineum)等,其中习水杜鹃的重要值最大,为0.426,是该群落乔木层的最优树种。灌木层共有29 种,重要值较大的物种有杜鹃、小果润楠、习水杜鹃、光枝杜鹃、江南樾桔等,其中以杜鹃的重要值最大,为0.804,虽然习水杜鹃排在第3,但重要值仅为0.362,与杜鹃差距较大,在灌木层中习水杜鹃的优势不明显。草本层物种有11 种,其中以箭竹的重要值(0.824)最大,广泛密布于林下。总的来说, 在乔灌层中均是杜鹃花科杜鹃花属植物为优势种, 表明杜鹃花科植物在该群落中有重要的建群功能。
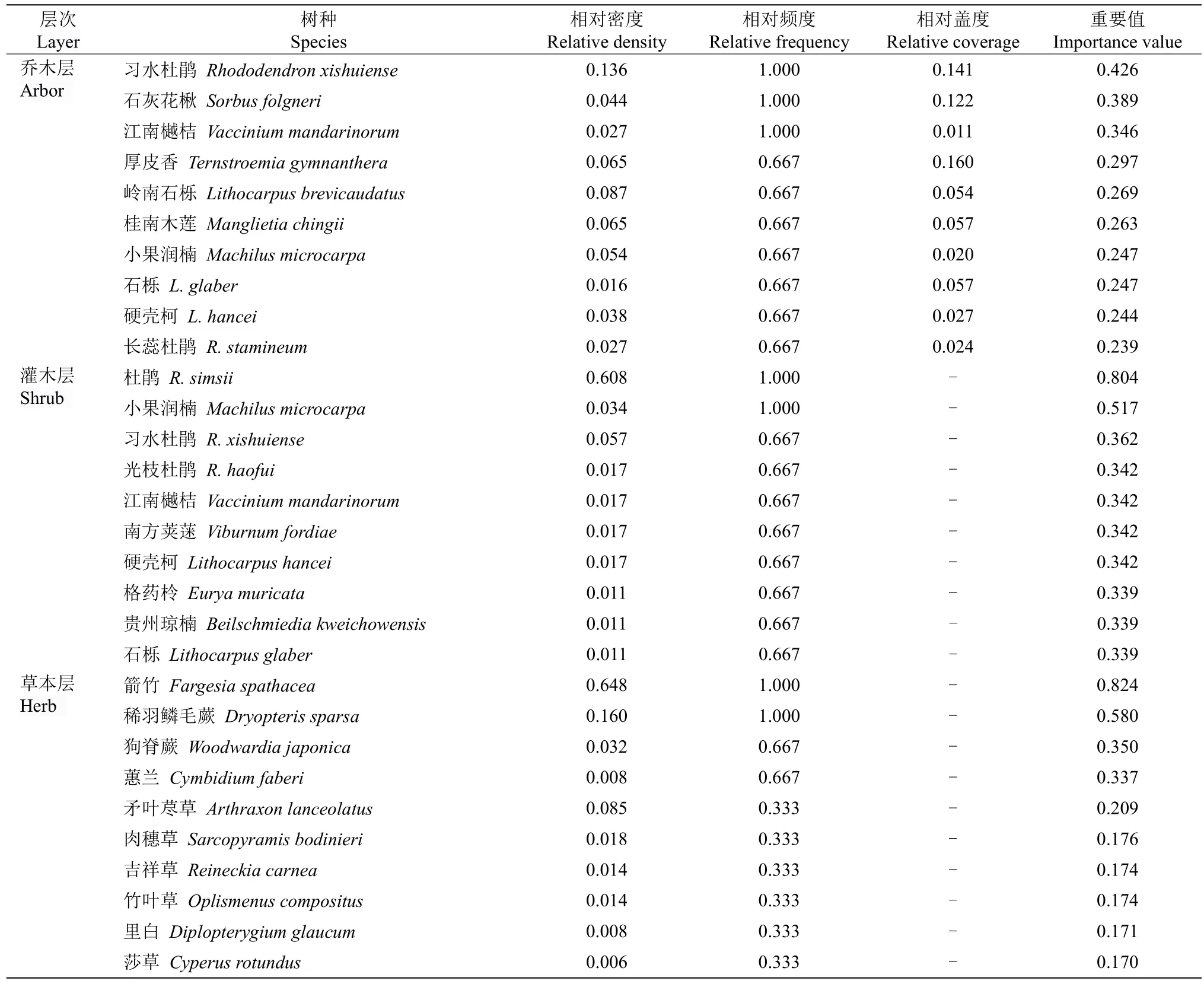
表3 习水杜鹃分布群落物种重要值(前10 种)Table 3 Important value of species in distribution community of Rhododendron xishuiense (top 10 species)
从表4 可见,Shannon-Wiener 多样性指数(H)、Pielou 均匀度指数(J)、Simpson 多样性指数(D)、Gleason 丰富度指数(R)均表现出乔木层>灌木层>草本层。其中除Simpson 多样性指数外,其他多样性指标乔木层均远远大于灌木层和草本层,表明乔木层物种丰富且多样性高,在该群落中具明显优势,主导着群落的发生发展。
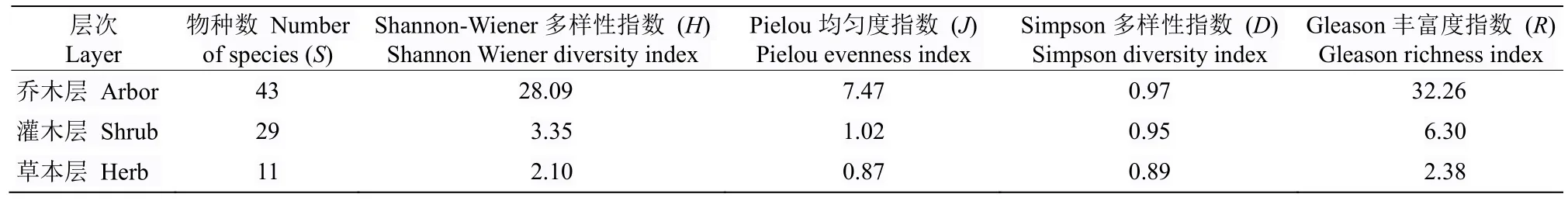
表4 习水杜鹃分布群落物种多样性指数Table 4 Species diversity index at all levels of Rhododendron xishuiense community
2.2 习水杜鹃种群动态
2.2.1 种群结构
由于本次样线调查未能发现其他的习水杜鹃分布地,故只能选择在发现地杨大老岭设置10 m×30 m 的样带开展调查。在样地中共调查到习水杜鹃34 株,年龄结构呈“金字塔”型(图3),其中第Ⅰ龄级数量最多, 有13 株(占38.24%),第Ⅱ龄级次之(11 株,占32.35%),习水杜鹃种群中,幼苗、幼树相对整个种群数量来说较多,大树、老树较少,总体上, 习水杜鹃种群个体数量随着年龄的增长而逐渐减少,在前3 个龄级时种群降低速度较均匀,但在第Ⅳ龄级时突然出现较强波动,甚至在该龄级出现了个体缺失的现象。

图3 习水杜鹃径阶结构Fig. 3 Diameter structure of Rhododendron xishuiense
2.2.2 种群的静态生命表
习水杜鹃的前3 龄级符合种群生命表的一般假设规律,第Ⅳ龄级存活个体急剧减少为0,出现了死亡率为负的情况,因此对Ⅲ、Ⅳ、Ⅴ龄级进行匀滑处理,得到ax。编制种群静态生命表(表5)可以看出,种群个体随龄级增加不断减少,死亡率qx和消失率Kx均也随着龄级增长逐渐上升, 在第Ⅳ龄级时达到最大值,整个种群个体到达第Ⅴ龄级时只有不到第Ⅰ龄级10%的个体存活下来。生命期望值ex在第Ⅰ龄级时最高,在第Ⅲ龄级时最低,第Ⅳ龄级存活率Sx最低,消失率Kx最大,该龄级种群开始衰退。
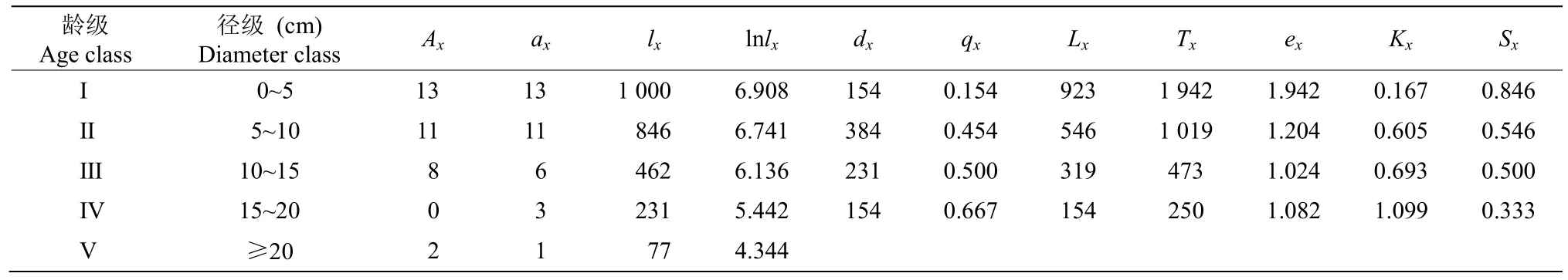
表5 习水杜鹃种群静态生命表Table 5 Static life table of Rhododendron xishuiense population
2.2.3 种群存活曲线、死亡曲线以及消失率曲线
习水杜鹃种群存活曲线可能是Deevey-Ⅱ型或Deevey-Ⅲ型(图4)。应用Hett 等提出的数学模型对习水杜鹃标准化存活个体数的自然对数值(lnlx)拟合,结果表明,指数模型拟合函数为Nx= 8.01563e–0.10485x,幂函数模型拟合函数为Nx=7.28124x–0.07726,且2 个模型的P<0.001 值也都达到极显著水平。相较2 个函数的拟合结果,指数函数模型的调整R2和F值均大于幂函数模型,指数函数模型的拟合效果更好,说明习水杜鹃种群存活个体数与龄级之间符合指数函数模型,种群存活曲线近似于一条直线,属于Deevey-Ⅱ型。拟合结果同样表明习水杜鹃种群随着年龄的增长,存活个体数量逐渐减少,与该种群的年龄结构和生命表的分析结果基本一致。
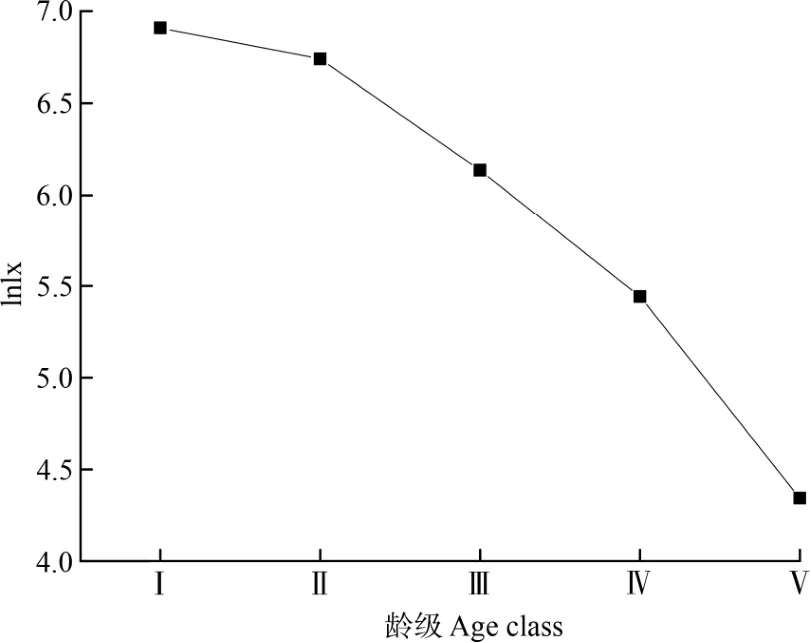
图4 习水杜鹃存活曲线图Fig. 4 Survival curve of Rhododendron xishuiense population
从习水杜鹃种群的死亡率qx和消失率Kx曲线(图5)可以看出,随着龄级增长均呈现逐渐上升, 在死亡高峰期时消失率也出现极大值,在死亡低峰期时消失率也出现极小值,该种群的死亡率和消失率曲线变化趋势基本保持一致,但死亡率曲线的变化幅度相对较小。习水杜鹃种群的死亡率和消失率到第Ⅳ龄级的时候达到最大值(qx=0.667,Kx=1.099),说明该龄级已接近种群生理寿命。
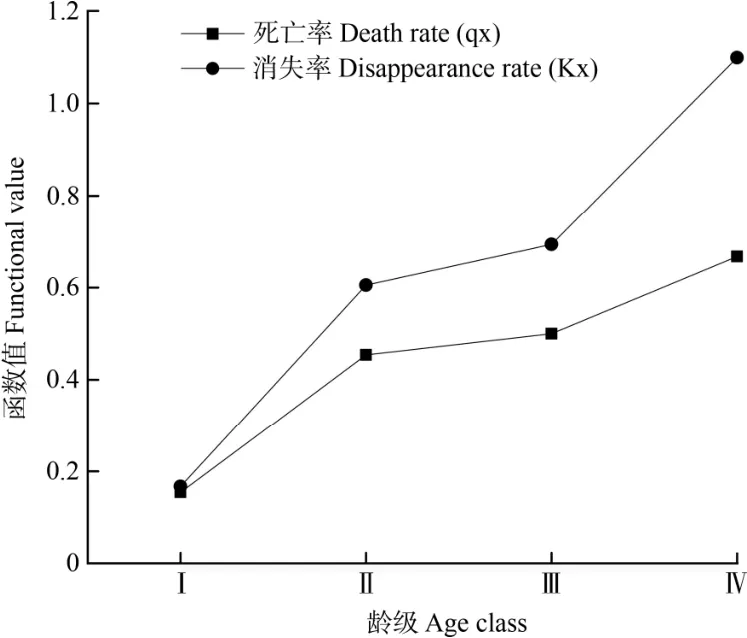
图5 习水杜鹃种群的死亡率、消失率曲线Fig. 5 Mortality rate and vanishing rate curves of of Rhododendron xishuiense population
2.2.4 种群的生存分析
习水杜鹃种群的生存率、累计死亡率、死亡密度、危险率函数值显示(图6),该种群的生存率持续下降,累计死亡率、危险率持续上升,死亡密度在第Ⅱ龄级出现波动,开始由上升转至持续下降,到第Ⅴ龄级时,种群的生存率接近0,而累计死亡率接近1,种群危险率也达到最大值,习水杜鹃种群开始进入生理衰退期。结合生存率、累计死亡率、死亡密度、危险率函数的结果,习水杜鹃种群生存状况展现出全期减少的特点,其中前期减少速度快、中期减少速度较慢、后期缓慢衰退。
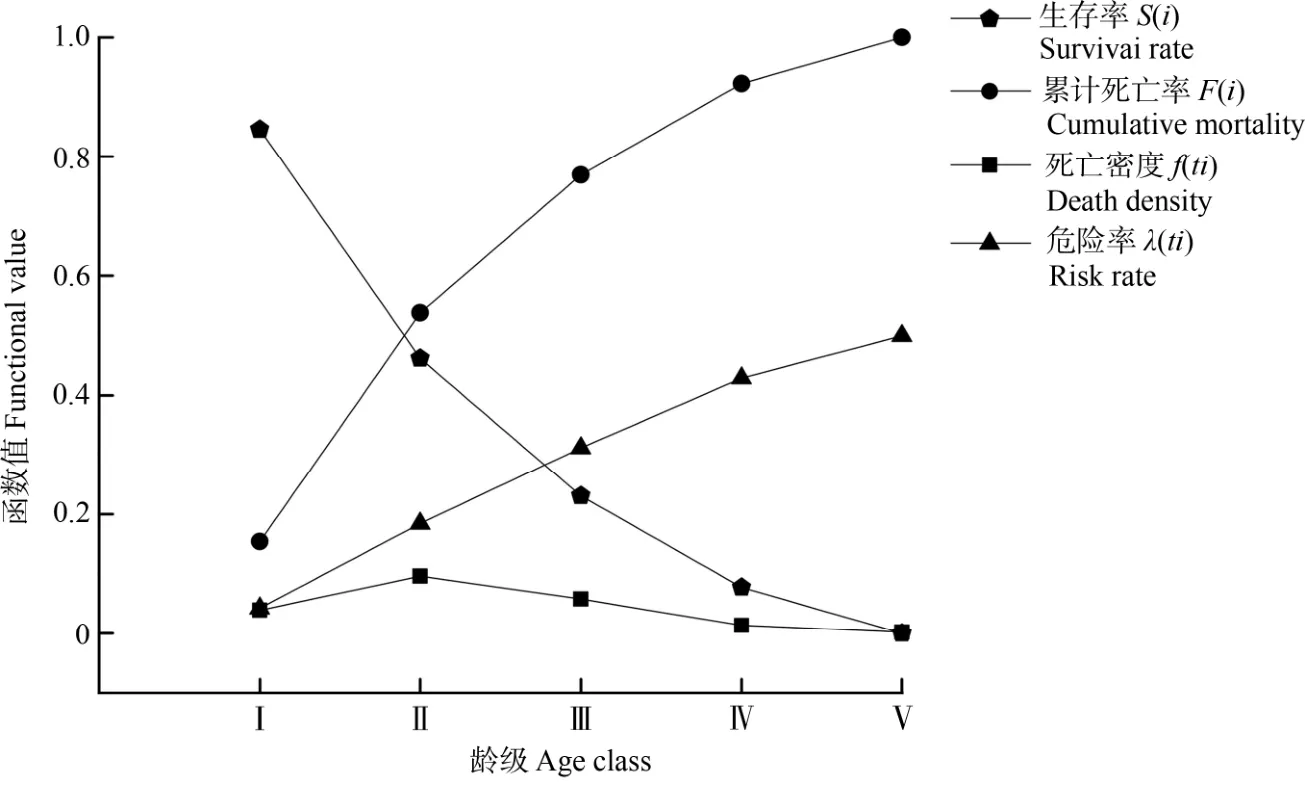
图6 水杜鹃种群的生存率、累计死亡率、死亡密度、危险率函数Fig. 6 Curves of survival rate, cumulative mortality rate, mortality density and risk function of Rhododendron xishuiense population
2.2.5 种群动态的量化分析
对习水杜鹃种群结构动态量化分析结果表明,各龄级的数量变化动态指数中,前3 个龄级均大于0 (其中V1=0.15,V2=0.27,V3=1),第Ⅳ龄级V4=-1,忽略外部干扰时习水杜鹃种群的数量变化动态指数Vpi=34.37%>0,考虑外部干扰影响时Vpi′=3.4%>0。综合习水杜鹃种群的量化分析结果,表明该种群属于增长型种群。虽然个体数量较少,但整个种群在现存的生境中形成了相应的生存策略,一定时间内,在无突发扰动的情况下,能较好的适应当地的环境并保持相对稳定。
3 结论和讨论
本次样线调查未能在贵州习水国家级自然保护区发现其他的习水杜鹃分布地。与五莲杨(Populus wulianensis)、华东黄杉(Pseudotsugagaussenii)等濒危物种相似,稀有物种在广泛的栖息地内有一个自然限制的范围,受特定空间环境的影响,成为了分布狭窄的栖息地的行家,但有限的空间和资源会限制种群发展[17,31]。中国西南部杜鹃分布的多样性受到地形复杂性和温度季节性共同影响[32],与此相同,习水杜鹃分布地海拔高(超1 500 m 山脊)、低温、潮湿等特定的外界环境,较高的异质性限制了习水杜鹃的传播,影响着习水杜鹃的分布,这也可能是在保护区其他区域暂未发现习水杜鹃分布的原因。受习水杜鹃分布范围的限制,本研究的样地较小,但不能因为“人口贫乏”和分布狭窄而忽略对特有植物的保护[3]。且习水杜鹃分布在较为狭窄的山脊上,周围地势陡峭,与山脊外的物种联系较小,较为符合群落的一般特征。群落结构分析中植物地理区系的划分对植物濒危等级、保护珍稀濒危物种与植物多样性具有重要意义[33],习水杜鹃分布群落的地理区系类型有14 个,地理成分多样,其中以热带植物较多,但伴有温带和中国特有植物,这与贵州习水国家级自然保护区属种亚热带的地理位置基本相符。温带植物分布占比较大的原因可能保护区是以丹霞地貌为主,热带植物更多分布在切割较深的沟谷,而本研究样地设置在海拔超1 500 m 的山脊, 综合来看,习水杜鹃所在群落是以热带性质为主,伴有温带性质山地森林植物区系。从系统发育方面看,本区有不少如樟科、木兰科、壳斗科、山茶科等原始或古老的植物。生活型是植物生态习性的体现,高位芽植物在习水杜鹃分布群落中占优势,是典型的亚热带常绿阔叶林的外貌组成特征,与保护区温暖湿润的气候相适宜[34],充分体现了该群落中植物对外界气候环境的适应性。植物优势种对群落内物种组成和结构、生物多样性、生产力、生态过程和功能均有重要影响,根据对习水杜鹃分布群落的重要值比较,杜鹃花科植物在乔灌层占较大优势,发挥着重要的建群功能,其中习水杜鹃对整个群落的建群作用较大,乔木层石灰花楸、江南樾桔,灌木层杜鹃、小果润楠,草本层箭竹等是群落中的优势物种,对群落发生发展也具有重要的影响。本次调查面积不到保护区面积百万分之一,统计到27 科42 属65 种,分别占保护区植物科属种8.7%、4.5%、2.8%,物种多样性较高,其中乔木层远远大于灌木层和草本层,主导着群落演变发展。
受外部环境和植物自身内部因素长期综合性的影响,植物更新个体随着时间推移在发生改变。种群结构的分析有助于推测种群过去和当下的生存状况、预测种群的更新动态[35]。本研究表明习水杜鹃个体极少,种群年龄结构呈“金字塔”型,幼苗幼树数量储备较多,种群属于增长型,但种群随着龄级增长逐渐减少,甚至在第Ⅳ龄级出现了缺少习水杜鹃存活个体的现象,这与苏佳佳等[36]对贵州习水国家级自然保护区内福建柏(Fokieniahodginsii)种群的研究结果相似,由于云贵高原干旱气候的干扰,使得种群个体在特定时间大量死亡或无法繁殖新的个体。但除了环境因素外,习水杜鹃种子萌发障碍机理尚不明确,还有历史上的、病虫害砍伐和极端天气等因素的影响,都有可能是造成第Ⅳ龄级出现个体缺失的原因。但种群现在低龄级储备的个体优势,说明了习水杜鹃在种群发展过程中,逐渐适应了环境的变化,甚至形成了一定的生存策略,以保障种群持续增长。种群静态生命表和存活曲线进一步量化了习水杜鹃种群的生存现状,揭示了习水杜鹃的生存能力和生存策略,对保护濒危植物具有重要意义[37–38]。从习水杜鹃的种群静态生命表得出,习水杜鹃种群的死亡率和消失率逐渐增大,且在第Ⅴ龄级均出现了最大值,这可能是随着年龄增长,资源需求增加,竞争加剧,特别是进入主林层的大树个体,随着冠幅增大,对养分、光照、水分等资源的竞争再次加剧,到老树阶段是生理衰老,老龄个体竞争力减弱,导致第Ⅴ龄级大量个体死亡。同时,生命期望值和存活率在第Ⅴ龄级也最低,反映出习水杜鹃大树个体在乔木层遭受的竞争尤为激烈。调查中也发现,习水杜鹃所在群落为人迹罕至的原始林区,林分郁闭度已接近上限,种群对资源的争夺必定激烈。存活曲线的拟合结果表明习水杜鹃种群属于Deevey Ⅱ型,为稳定型种群,但由于整个种群个体数量较少,特别是大树数量稀少,若不能提高低龄级个体存活率,随时间推移,无法使中龄级个体数得到补充,则种群将有可能朝着衰退的趋势发展,存在灭绝的风险。生存率、累计死亡率、死亡密度、危险率函数的结果进一步补充证实了上述结论的合理性。种群结构动态量化分析可以直观的反映种群发展趋势,习水杜鹃种群结构的量化分析结果表明,种群属于增长型,相较整个种群而言,低龄级个体数量较多,在现存环境未遭到破坏或不遭受巨大扰动的前提下,习水杜鹃能够维持后续龄级的更新,从而保持种群的相对稳定,但习水杜鹃种群个体总量较少,幼苗幼树的更新补充和茁壮成长是保障种群发展的关键因子,当前群落中物种丰富度较高,在垂直结构上,各层次物种之间的互补性也会越高,整个群落有效地利用林下部分的资源[39–40],使得整个群落可被利用的空间和资源极少,竞争激烈的环境中习水杜鹃种群新的个体繁殖生长困难。调查中还发现习水杜鹃的花果易受虫害,结实率不高,习水杜鹃的种群发展趋势不容乐观。就目前调查到的习水杜鹃分布数量,按照目前国际上应用最广泛的IUCN 濒危物种等级标准,习水杜鹃属于濒危(CR)等级植物,亟需开展相应的保护工作。
综上,习水杜鹃种群分布范围狭窄,种群总量极少,且遭受较强的竞争,遭遇环境变化或巨大扰动(病虫害等)时,随时有灭绝的风险。建议将习水杜鹃列为保护区的最高关注级别,采取适当措施促进幼苗幼树生长,使中年个体得到有效补充,以保障种群持续增长。另外,本次调查的结果形成了一个“生态档案”,保护区的工作者可以据此对习水杜鹃进行监测,并根据监测结果实时制定科学的保护方案。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
心声歌刊(2021年3期)2021-08-05
河南农业大学学报(2020年2期)2020-05-22
小聪仔(幼儿版)(2020年12期)2020-02-01
星星·散文诗(2019年15期)2019-12-29
星星·散文诗(2019年5期)2019-07-19
山西林业科技(2019年1期)2019-05-27
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
大众考古(2015年7期)2015-06-26
