
西南地区六种魔芋属植物基于cpDNA序列的遗传多样性研究
2023-12-19 02:12:44陆飞东
广西植物 2023年11期
殷 斯, 郝 转, 陆飞东, 高 永*
( 1. 曲靖师范学院 生物资源与食品工程学院, 云南 曲靖 655011; 2. 渭南师范学院 环境与生命科学学院, 陕西 渭南 714099 )
探究栽培作物野生资源的遗传多样性及遗传分化机制对种质资源的收集与品种改良具有重要的应用价值和科研意义。魔芋是天南星科(Araceae)、魔芋属(AmorphophallusBlume)的多年生草本植物(白立伟等,2016),主要分布于西非、亚洲中南半岛、中国西南山地和东南亚国家的热带及亚热带地区,全球约200种,我国约有20种(刘佩瑛,2004)。魔芋在我国具有悠久的栽培利用历史并且野生种质资源丰富,其地下球状块茎富含葡甘聚糖,具有重要的经济价值(李恒和龙春林,1998)。近年来,我国的魔芋种植面积不断扩大,自2017年起,西南各省的总种植面积已达到世界第一(Srzednicki &Borompichaichartkul, 2020)。但是,由于长期无性繁殖,魔芋种质出现了品质下降、产量降低、病害流行等问题,因此,要推动魔芋产业的进一步发展,加快野生种质资源的收集与遗传多样性评价已刻不容缓(宣慢,2010)。
20世纪80年代起,我国对魔芋种质资源的起源与进化、种质的亲缘关系和分类等进行了研究并取得一定的成果(牛义等,2005)。李恒研究整理了中国魔芋属植物19种,包括中国特有的8个种,后续又发现命名了3个新种(李恒,1988)。一些分子生物学研究利用ISSR、RAPD、AFLP等分子标记,探讨了魔芋部分栽培品种之间的亲缘关系(张盛林和孙远航,2006;任盘宇和潘明清,2013;Pan et al., 2015)。但是目前,我国在魔芋野生资源的亲缘关系特别是分子系统进化方面的研究还较为欠缺,各地命名混乱,同种异名和同名异种的现象较为突出,特别是对野生魔芋资源的遗传变异缺乏系统研究(牛义等,2005)。
作为西南地区的特色经济作物,魔芋具有重要的经济与生态价值(赵培城等,2015)。但环境污染、生境破坏等人类活动,造成了魔芋野生群体的衰退(牛义等,2005)。作为种质资源改良的优良材料,加快野生种质资源的研究与利用,对魔芋的品种改良具有重要意义。当前对魔芋野生群体的遗传背景缺乏了解,给资源保护和育种工作造成了很大阻碍(Gao et al., 2017)。叶绿体DNA(cpDNA)为单系遗传,遗传过程不经历基因重组,被广泛地应用于植物系统发育、遗传多样性评价等研究中(李娅翔等,2020)。cpDNA 片段trnK-matK、rbcL和trnL在天南星科的系统进化研究中,被证明具有通用性好、突变率高的特点(Grob et al., 2002; Sedayu et al., 2010)。破碎生境对种群动态的影响及演化趋势是保育遗传学研究的热点问题,遗传多样性则是推断物种进化潜力的重要指标之一(王峥峰和葛学军,2009)。为评估西南地区魔芋属物种野生群体的遗传多样性,探究代表物种的系统发育地位,本研究采用以上3个cpDNA片段对西南地区6种魔芋属植物的野生群体进行遗传多样性及种间系统发育研究,以期为西南地区魔芋物种的保护提供指导,为今后魔芋的种质创新提供理论依据。
1 材料与方法
1.1 试验材料
2019—2021年,本研究采集南方五省(区)(云南、广西、贵州、湖南、江西)的魔芋属6个种,即花魔芋(Amorphophalluskonjac)、东京魔芋(A.tonkinensis)、西盟魔芋(A.krausei)、滇魔芋(A.yunnanensis)、疣柄魔芋(A.paeoniifolius)、东亚魔芋(A.kiusianus)的野生群体。采样过程中,随机选择植株,每株样本间隔至少5 m,采集叶片置于硅胶中干燥保存(表1)。采用植物基因组DNA提取试剂盒(天根,北京)提取魔芋叶片总基因组DNA,1.5%琼脂糖凝胶进行电泳检测。
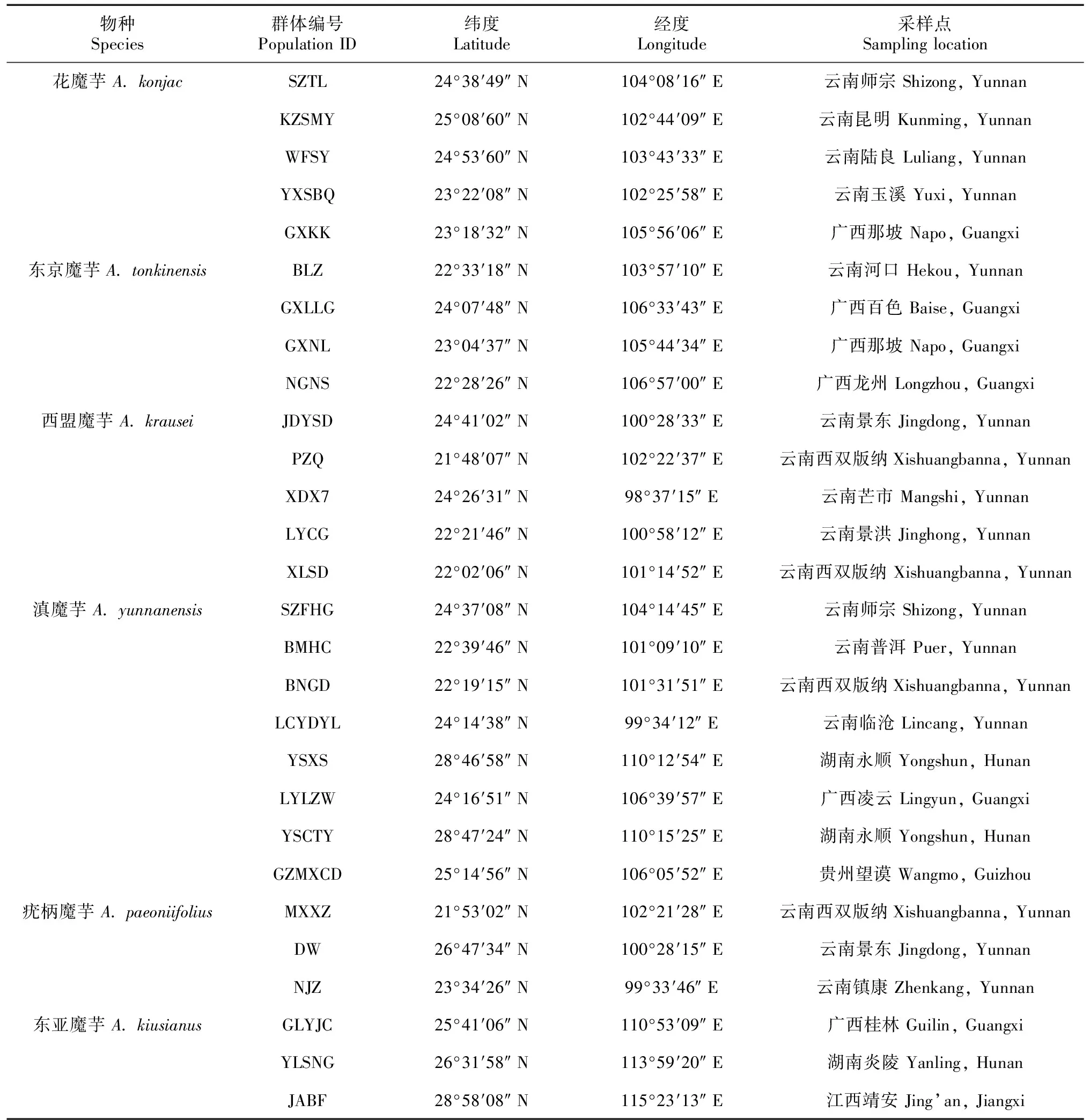
表 1 本研究的魔芋群体样本采集地点信息Table 1 Geographic information of Amorphophallus populations collected in this study
1.2 PCR扩增和测序
利用已发表的引物对trnK-matK、rbcL和trnL 3个cpDNA片段进行PCR扩增(表2)。PCR扩增体系总体积为30 μL,包含20~50 ng DNA模板,正、反向引物(10 μmol·L-1),2×PCR StarMix(GenStar)和ddH2O等。反应程序:预变性,95 ℃,5 min;35个扩增循环,95 ℃变性30 s,56 ℃退火30 s,72 ℃延伸60 s;终延伸,72 ℃,7 min。扩增产物经1.5%琼脂糖凝胶电泳检测,合格产物送生工生物工程(上海)股份有限公司进行双向测序。
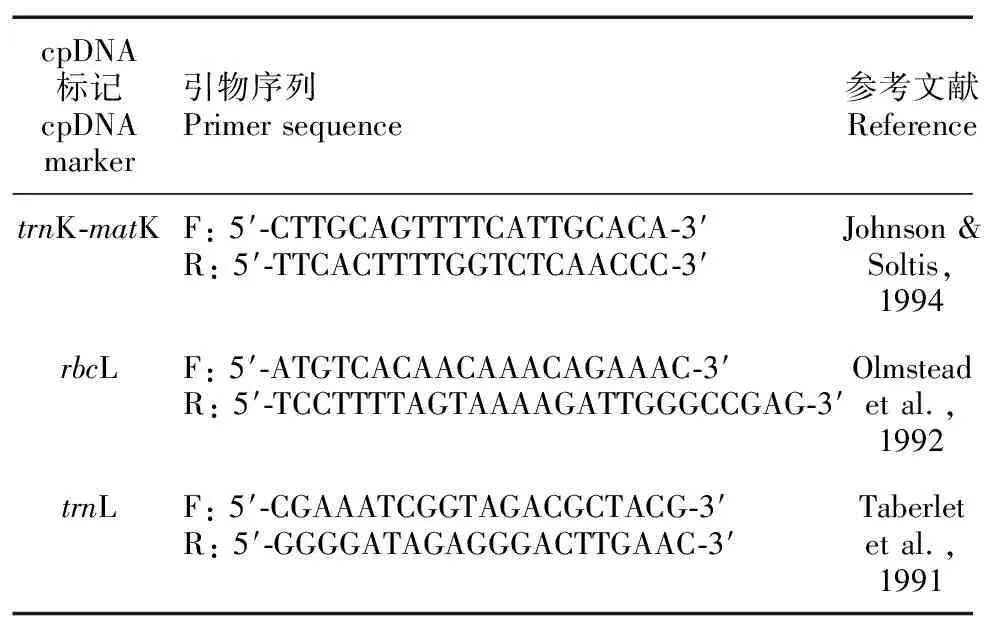
表 2 cpDNA片段扩增所用引物序列Table 2 Primer sequences used in the amplification of cpDNA fragments
1.3 数据分析
采用Lasergene软件的Seqman工具对3个cpDNA片段进行拼接(DNAStar Inc., Madison, WI, USA);使用MEGA v7.0的Clustal W算法对所有样本的DNA序列进行比对,将同一个体的3个cpDNA片段序列串联作为一个整体进行分析(Kumar et al., 2016)。在遗传多样性评估时,将连续多碱基的插入/缺失突变视为单突变事件(Simmons &Ochoterena, 2000)。为减少误差,样本量少于3个的群体(KZSMY、GXKK)不用于群体遗传多样性分析。采用Arlequin软件评估6个物种间的遗传分化系数(FST),并用1 000次模拟运算评估显著性(Excoffier &Lischer, 2010)。
为评估魔芋6个物种之间的亲缘关系及系统进化地位,本研究使用IQ-TREE 1.6.12对cpDNA单倍型序列进行核酸替代模型检测,构建最大似然法(maximum likelihood,ML)系统进化树,运行1 000次bootstrap检测显著性(Nguyen et al., 2014)。此外,采用SplitsTree 4.14.6对所有单倍型序列构建Neighbor-Net树,评估魔芋种间的网状进化关系(Huson &Bryant, 2006)。
2 结果与分析
2.1 cpDNA序列扩增
对来源于28个野生群体的170个样本分别进行cpDNA序列扩增,3个cpDNA片段的电泳检测结果表明,PCR目的产物条带清晰,符合测序要求(图1)。
2.2 基于cpDNA的物种遗传变异与遗传多样性
经样本间的序列拼接和比对,得到的3个叶绿体片段trnK-matK、rbcL和trnL的序列长度分别为719、1 312、315 bp。将同一个体的3条序列串联进行整体分析,序列总长度为2 346 bp。在所有样本序列中检测到128个多态性位点,其中插入/缺失突变位点78个,共得到57个单倍型。遗传多样性分析表明,群体遗传多样性较低,核苷酸多样性(用π表示)在0~0.009 31之间。在部分群体只有一个单倍型(26个群体中的12个),群体DW(疣柄魔芋)和NJZ(疣柄魔芋)具有最高的群体单倍型多样性(Hd=1.0)。在物种水平,每个物种的单倍型数量从4到14不等,其中疣柄魔芋和东京魔芋的遗传多样性较高(表3)。在花魔芋、东京魔芋、西盟魔芋和滇魔芋的种内群体间发现了共享单倍型,但6个物种之间没有共有单倍型存在。物种间的两两遗传分化表明,花魔芋和滇魔芋两个物种间的遗传分化系数最高(FST=0.942),而花魔芋和西盟魔芋的遗传分化系数最低,FST为0.481(表4)。
2.3 魔芋属系统发育关系
为评估魔芋属各物种的系统发育关系,结合GenBank数据库下载的21个魔芋物种的cpDNA序列进行系统发生分析(表5),根据BIC(Bayesian information criterion)评分,HKY+F+R2模型被检测为最优核酸替换模型。最大似然树表明,27个魔芋物种聚成3个主要分支,即非洲分支、东南亚分支和东亚大陆分支。本研究关注的6个物种则以较高的支持率分别被划分在东亚大陆分支和东南亚分支,疣柄魔芋列入东南亚分支。东亚大陆分支又进一步分化为两支(分支A和分支B),分支A包含花魔芋和西盟魔芋,分支B由东亚魔芋、滇魔芋和东京魔芋构成 (图2)。Neighbor-Net网状进化分析的结果与ML树基本一致, 支持东亚魔芋、滇魔芋和东京魔芋构成一个分支且花魔芋与西盟魔芋的亲缘关系较近的结论(图3)。
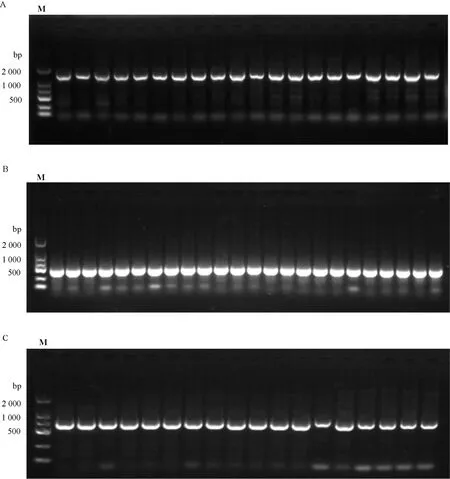
M表示DL2000 Marker。M indicates DL2000 Marker.图 1 部分样本3个cpDNA片段 [rbcL (A)、trnL (B)、trnK-matK (C)]扩增产物电泳图Fig. 1 Electrophoretograms of amplification products of three cpDNA fragments [rbcL (A), trnL (B) and trnK-matK (C)]
3 讨论与结论
遗传多样性与遗传分化不仅能反映物种遗传变异的高低,还能在一定程度上推断群体的演化机制,为制定保护措施提供依据(王峥峰和葛学军,2009)。本研究检测到西南地区魔芋属的6个物种存在低水平的群体遗传多样性,造成此现象的主要原因可能是植物的自身散布能力有限,西南山地的生境阻隔造成了群体遗传多样性降低(Gao et al., 2015)。我国西南山地的多种植物,如苦苣苔、胡黄连等, 也被报道由于隔离造成了低水平的群体遗传多样性(Gao et al., 2015; 李国栋等,2016; Wang et al., 2017)。此外,近年来西南地区野生魔芋的生境由于人为破坏呈破碎化状态,加剧了种群衰退和地理隔离,这是群体遗传多样性降低的人为因素(贺水莲等,2016)。针对该情况,可根据魔芋的遗传分化格局,确定核心种质资源,通过建立群体种源保护地、种质资源圃等多种措施来进行保育(臧润国等,2016)。在物种水平上,滇魔芋的遗传多样性在6个物种中最低,预示着该物种适应环境变化的能力较其他物种更弱,后续应加强对其野生群体的保护。
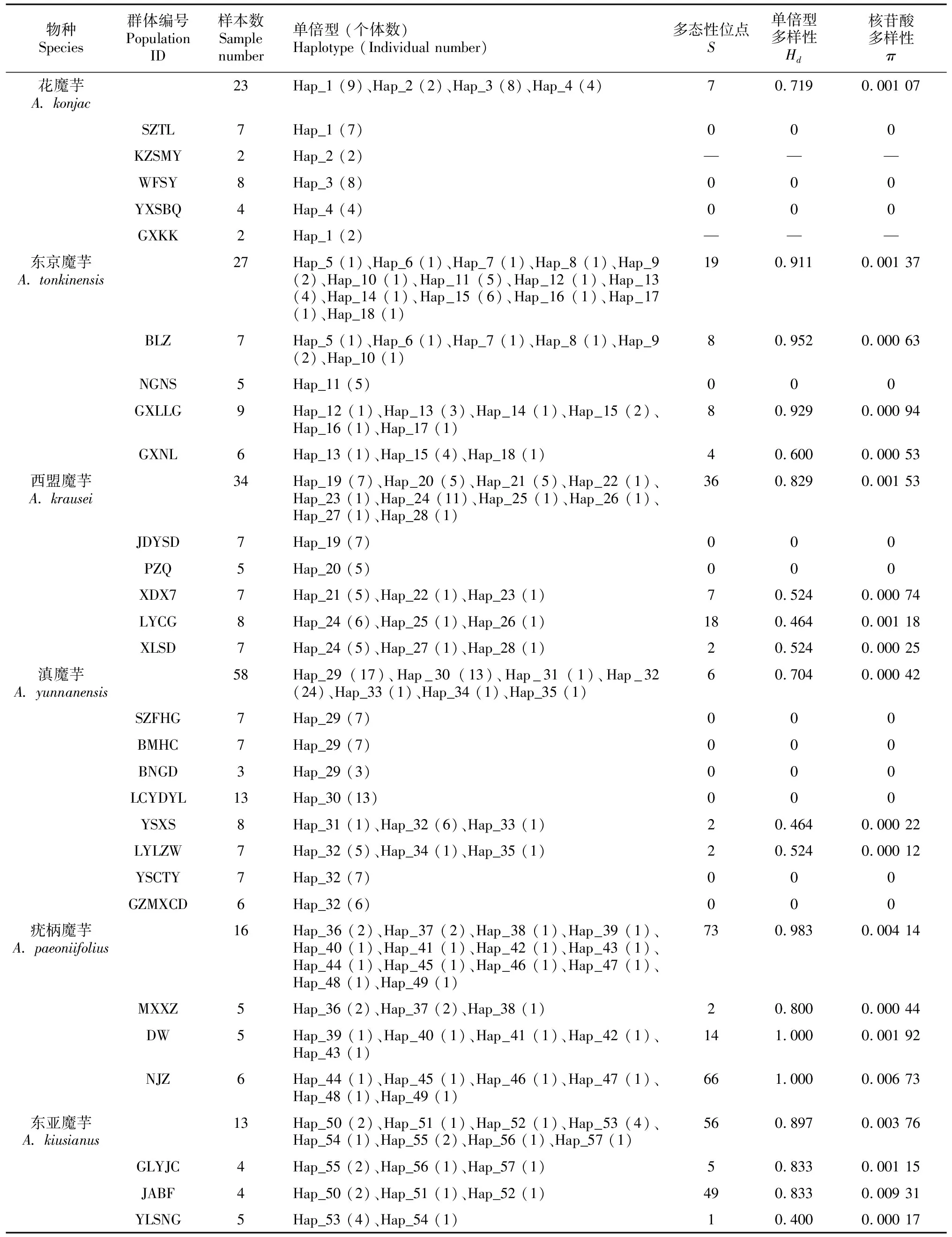
表 3 西南地区魔芋属6个物种的cpDNA序列遗传多样性参数Table 3 Genetic diversity parameters of six Amorphophallus species from Southwest China surveyed for cpDNA sequences
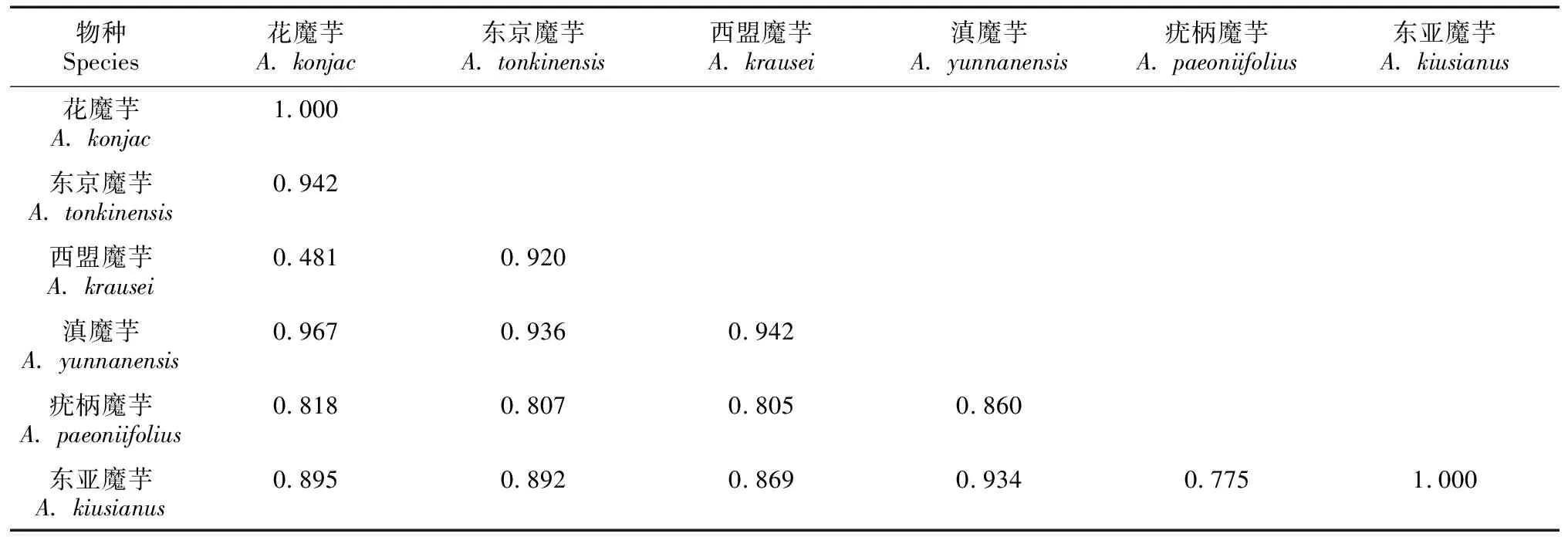
表 4 西南地区魔芋属6个物种的种间遗传分化系数(FST)Table 4 Genetic differentiation coefficient (FST) among six Amorphophallus species from Southwest China

表 5 本研究下载的魔芋cpDNA序列NCBI GenBank登录号Table 5 Accession numbers of cpDNA sequences downloaded from NCBI GenBank in this study
系统发育分析表明魔芋属物种按照地理分布形成3个主要分支,而东亚大陆的类群则进一步分化为两支(Claudel et al., 2017)。目前对魔芋东亚大陆类群分化为两支的原因尚未有明确结论,两个分支之间存在地域重叠,魔芋属早期演化的快速扩张可能导致了该分化格局(Claudel et al., 2017)。形态学研究认为,东亚大陆两个类群间的果实颜色存在差异,可能与吸引相应昆虫进行种子散播有关(Claudel et al., 2017)。鉴于此,生态适应可能也是东亚大陆类群的分化机制之一。细胞学研究也为推断该属的演化历史提供了一些佐证(张风洁,2014)。学者认为2n=26是魔芋属的原始核型,2n=24和2n=28由减数分裂过程配子丢失或增加1条染色体造成,而2n=39则可能是由未正常减数分裂的配子(n=2x=26)与正常减数分裂的配子(n=x=13)杂交形成(Zhao et al., 2021)。针对本研究的物种,东南亚分支的疣柄魔芋和A.commutatus的染色体数量分别为2n=28和2n=26,东亚大陆分支的花魔芋、西盟魔芋、滇魔芋和谢君魔芋的染色体数目均为2n=26(张风洁,2014)。目前染色体核型研究涉及的魔芋物种较少,后续需要解析更多物种的染色体组型,以期更深入了解魔芋属的演化历史及规律。
本研究鉴定了西南地区魔芋代表物种间的亲缘关系,虽然我国魔芋栽培面积位居世界第一,但育种资源狭窄,当前品种已不能满足种植产业的需要。要解决魔芋的良种问题,杂交育种不失为一种可行的解决方案。目前已有一些研究开展了魔芋的种间杂交实验,但需要注意因远缘杂交成功率低、子代不育等造成经济损失(刘二喜等,2020)。根据本研究得出的魔芋属物种的亲缘关系合理选择杂交亲本组合,可为今后魔芋的种质创新、杂交育种及杂种优势群的构建提供理论依据。
综上所述,为对我国西南地区的魔芋野生资源进行遗传评价,研究利用3个cpDNA标记分析了魔芋属6个物种的遗传多样性与遗传分化,鉴定了各物种之间的系统进化关系。研究结果表明,生境隔离和人为干扰可能导致了魔芋野生群体的遗传多样性降低,东亚大陆类群分化为两支可能与早期演化的快速扩张和生态适应相关。魔芋属种间的亲缘关系鉴定可为后续的野生资源利用奠定理论基础。
猜你喜欢
东疆学刊(2021年4期)2021-02-12 01:50:18
今日农业(2019年14期)2019-09-18 01:21:42
学生天地(2019年28期)2019-08-25 08:50:54
外国问题研究(2018年2期)2018-07-04 06:50:16
数学物理学报(2018年1期)2018-03-26 08:16:36
金色少年(奇趣科普)(2016年9期)2016-10-21 07:26:49
中国酿造(2016年12期)2016-03-01 03:08:27
教学与研究(2016年1期)2016-02-28 01:59:27
山西大同大学学报(自然科学版)(2014年3期)2014-01-23 01:56:30
时代人物(新教育家)(2013年3期)2013-08-21 13:25:34
