
浙江九龙山香果树径级结构和空间分布格局
2023-12-13 08:35郑子洪郑伟成杨晓君唐战胜郑蓉骆争荣
浙江林业科技 2023年6期
郑子洪,郑伟成,杨晓君,唐战胜,郑蓉,骆争荣
(1. 浙江九龙山国家级自然保护区管理中心,浙江 遂昌 323300;2. 遂昌县生态林业发展中心,浙江 遂昌 323300;3. 浙江师范大学 生命科学学院, 浙江 金华 321004;4. 丽水学院 生态学院,浙江 丽水 323000)
香果树Emmenopteryshenryi为茜草科Rubiaceae 香果树属Emmenopterys落叶大乔木,是第四纪冰川孑遗植物之一[1]。香果树主要分布在我国陕西南部和甘肃南部、华中、华东、华南和西南地区15 个省区海拔430 ~ 1 630 m 处岩石裸露度高、土层浅薄的山谷林中[1-3]。它是已知的香果树属中唯一现存种,其同属近亲在北美和欧洲都已灭绝,目前香果树已经成为了香果树属系统演化上的一个盲枝[4]。由于人类过度开发森林和土地资源,生境片段化加剧,香果树的生境受到了极大的破坏,加上香果树种子和幼苗在野外的萌发率和存活率低,使得其天然更新能力差,种群数量越来越少,正濒临灭绝,已被列为国家二级保护植物[5-6]。过去40 多年,学者们从种子萌发、生理特性、物候特征、种群动态及遗传多样性、群落结构和繁育技术等方面对香果树进行了广泛的研究,取得了重要的研究成果[2,4]。然而,对于香果树在野外的空间格局以及与其它物种的空间关系影响香果树在群落中的生存和更新仍知之甚少。
种群的径级结构能很好地反映种群的更新状况和动态变化。径级结构分析一直是香果树研究的重点领域。然而,来自不同地区的有关香果树径级结构的研究结果存在不少差异,影响香果树种群结构的因素也一直没有得到清晰的揭示[2]。另一方面,种群的空间分布格局不仅对于其利用资源而且作为其他物种资源被利用及其繁殖都非常关键[7]。不同种群的空间关系决定了它们之间的竞争势及其空间生态位,在很大程度上决定了群落的稳定性和发展方向[8]。任何在森林群落构建中起作用的生态学过程都会在空间上留下可以被检测的印迹,空间格局分析可以用来推断很多潜在生态学过程[9]。分析种群时间动态和空间过程,使两方面信息相互印证和补充,有利于揭示影响香果树生存繁衍的重要因素。
本研究采用香果树种群径级结构分析结合空间点格局分析,探讨香果树种群的动态变化、空间分布和香果树与伴生树种的空间关系,以期从群落学上揭示其濒危的原因,为香果树的有效保护提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
浙江九龙山国家级自然保护区介于28°19′10″ ~ 28°24′43″ N、118°49′38″ ~ 118°55′03″ E 之间,处于浙、闽、赣的交界处,遂昌县西南部。气候属中亚热带湿润季风气候,四季特征明显不同,年均温约17 ℃;最冷月为1月,极端最低温为-17 ℃;最热月为7 月,极端最高温为37 ℃;年均降水量约为1 855 mm,年均相对湿度83%,夏季高温多雨,冬季寒冷潮湿。因地形复杂,九龙山保护区的气候具有垂直地带性,高海拔区域气候近似温带。土壤属富铝土纲,红壤土类,包括老红壤、红壤、黄红壤和红黄壤等亚类,随着海拔的变化,各亚类土壤按序替代。植被以亚热带常绿阔叶林最为典型,分布最为广泛[10]。
1.2 样地调查
2021 年7 月12 日至10 月7 日,在浙江九龙山国家级自然保护区的大岩前、岩背坑、陈坑、源大坑、内阴坑、外九龙等地共设置35 个固定样地(总面积24 740.04 m2,下文简称“样地”)。每个样地是以香果树为圆心,15 m 为半径的样圆。样地设置时,首先需要选一棵香果树作为样地中心。中心香果树的选择原则,一是保证所有径级都有代表作为中心香果树;二是通过中心树的选择尽量用较少的样圆让同一分布点的香果树都被圈入样圆,纳入长期监测。测量每个样地的海拔和地理坐标。用塑料绳拉出样地大致范围后,给样地内所有胸径2.5 cm 以上的乔、灌木挂上(用不锈钢丝绑缚)带有唯一编号的铝牌,鉴定物种名,用游标卡尺(当树木胸径小于3 cm 时使用,同一高度十字交叉测两次取平均)或胸径尺(当树木胸径大于3 cm 时使用)测量胸径,并用莱卡手持激光测距仪测量每棵符合标准的乔、灌木到中心香果树的水平距离。将所有数据记录在案。树木胸径(DBH)是指离地面高度1.3 m 处树木的直径。树木在1.3 m 高度以下有分叉的,测量并记录各个分叉的胸径。
1.3 径级结构和存活曲线
为分析径级结构成因和种群未来趋势,将所有样地内香果树(含样地中心香果树和非中心香果树)的胸径(DBH)(如存在多个分叉时,取最大胸径)进行径级结构分析,参照文献[11]方法,本研究以5 cm 为1 个径级,一共分为12 个径级。A 级胸径为2.5 ≤ DBH < 7.5 cm,B 级胸径为7.5 ≤ DBH < 12.5 cm,C 级胸径为12.5 ≤ DBH < 17.5 cm,D 级胸径为17.5 ≤ DBH < 22.5 cm,E 级胸径为22.5 ≤ DBH < 27.5 cm,F 级胸径为27.5 ≤ DBH < 32.5 cm,G 级胸径为32.5 ≤ DBH < 37.5 cm,H 级胸径为37.5 ≤ DBH < 42.5 cm,I 级胸径为42.5 ≤ DBH < 47.5 cm,J 级胸径为47.5 ≤ DBH < 52.5 cm,K 级胸径为52.5 ≤ DBH < 57.5 cm,L 级胸径为DBH ≥ 57.5 cm。
存活曲线可以直观地表达同龄(或同径级)群体的存活过程。本文以径级(x)为横坐标,以从x到x+1 径级间隔内标准化存活量(Lx,即径级x和径级x+1 的平均标准化存活量)的自然对数ln(Lx)为纵坐标,绘制九龙山香果树种群的存活曲线。径级x的标准化存活量为径级x 的存活数占单径级最高存活数的千分数。根据Deevey的定义,种群存活曲线一般可分为三种[12]。Ⅰ型曲线为凸形,幼年个体的存活率高,老年个体的存活率低;Ⅱ型曲线呈对角线型,在所有年龄阶段的死亡率稳定;Ⅲ型曲线凹形,幼年个体的死亡率极高。Ⅰ型存活曲线为幂函数,Ⅱ型存活曲线为指数函数,Ⅲ型存活曲线为幂函数,其计算公式如下[13]。
Ⅰ型存活曲线:
Ⅱ型存活曲线:
Ⅲ型存活曲线:
式中,x为径级,Lx为平均标准化存活量,模型参数L0、a和b均由回归分析估计得到。用实际观测数据对各型曲线进行函数拟合,使用Akaike 信息量准则(AIC)进行模型选择,即在上述3 个模型中选择AIC 统计量最小的模型作为最优模型。
1.4 点格局分析方法
本研究选用成对关联函数g(r)来分析香果树的空间分布格局以及中心香果树与(不同径级的)其它种类伴生树的空间关系,其计算公式如下[14]:
式中,K12(r)是基于类型1 的第i点和类型2 的第j点之间的所有距离dij(单位m)的函数,由下式给出[14-15]:
式中,r是计算格局指数的距离(单位m);n1和n2分别是类型1 和2 的点数;A是35 个样地的总面积;Ir是一个判断变量,如果dij≤r,那么Ir(dij) = 1,如果dij>r,那么Ir(dij) = 0;Wij是一个用于校正边缘效应的加权因子,权重Wij是以类型1 第i点为中心、半径为dij的圆面积在研究区域内的比例[15],本研究无边缘效应,取值为1。本研究将35 个样地的中心树作为类型1,而将其它香果树或者其它种的伴生树作为类型2。
为了分析香果树与伴生种不同径级的树木间的空间关系,本研究将伴生种树木按胸径大小分成4 个级别,分别为小树(2.5 cm ≤ DBH < 7.5 cm)、中树(7.5 cm ≤ DBH < 22.5 cm)、大树(22.5 cm ≤ DBH < 37.5 cm)和超大树(DBH ≥ 37.5)。
为了进一步分析中心香果树与样地内不同大小的其它香果树或其它伴生树在空间上的关系,本研究用标记关联函数来分析树木胸径值之间空间相关。标记关联函数估算公式Km(r)如下:
式中,m(u)和m(v)是连接到过程的两个点上的标记(即树木的胸径,单位cm),位置u和v由距离r(单位m)隔开,而M和M'是标记边际分布的独立格局,f(m(u),m(v))可以视为返回非负实值的标记对定义的任何函数。本研究将其定为f[(m(u),m(v))] =m(u)×m(v)。
本研究选择完全空间随机(CSR)作为香果树空间分布和中心香果树与其他种类伴生树空间关系分析的零模型[16],即保持中心香果树的位置不变而每个样圆中的其它个体在研究区域的任何位置发生的概率相等,并且一个点的位置独立于任何其它点的位置(即点之间不相互作用)。在分析树木胸径空间关系时,本研究选用随机标签零模型。将点格局内的点位置固定,在每个点上随机贴上标签(即胸径)。
本研究利用999 次Monte Carlo 模拟构建出零模型条件下的95%置信区间包络线[15]。在此基础上,进行结果偏离零模型状态的显著性检验。由于同时对多个尺度下的空间格局是否符合零假设进行推断可能会提高统计推断的第1 类错误[17]。本研究首先用适合度检验(GOF test)来检测数据是否在全局上符合零假设。如果全局适合度检验的P值大于0.05,不管其在具体尺度上表现如何都视为符合零模型。如果P值小于0.05,当实际曲线位于置信区间上时,种群呈聚集分布,种间关系为相互聚集;曲线位于置信区间内,种群为随机分布,种间的空间分布是相互独立的;当位于置信区间下方,种群则为均匀分布,种间关系为相互排斥。以随机贴标签法建立零模型时,当观测值小于置信区间下限时,表示给定距离内点的胸径差异大;当观测值大于置信区间上限时,表示给定距离内点胸径差异很小。点格局分析借助R 语言软件包spatstat 进行。
2 结果与分析
2.1 径级结构和存活曲线
在35 个样地内一共调查到香果树93 棵,最大胸径为66.3 cm,最小胸径为5.9 cm,平均胸径为23.1 cm。样地中所有DBH ≥ 2.5 cm 的香果树径级结构分布图(图1A)显示,径级C 级(12.5 ≤ DBH < 17.5 cm)的植株个体最多,占总数的27.96%,径级B ~ F 级(7.5 ≤ DBH < 32.5 cm)的植株个体占总株数的78.49%。
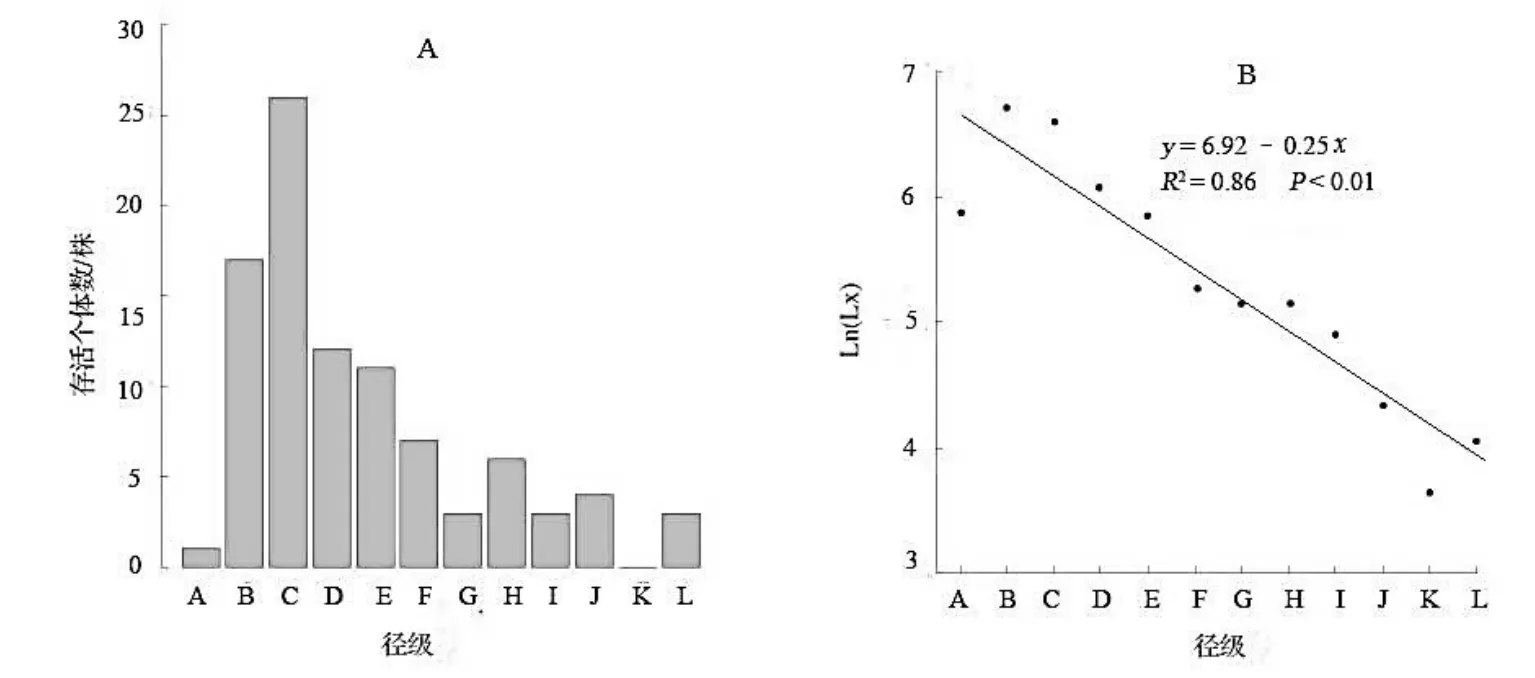
图1 九龙山香果树种群的径级结构(A)和存活曲线(B)Fig. 1 Diameter class distribution(A) and survival curve(B) of E. henryi population in Zhejiang Jiulongshan National Nature Reserve
径级大于I 级(DBH >42.5 cm)的香果树数量很少,且缺少径级K 级(52.5 ≤ DBH < 57.5 cm )的香果树个体。九龙山香果树种群在各径级的存活量虽有所波动,但总体上随着径级的增大,存活个体数稳定下降(图1B),模型比较结果表明Deevey 的Ⅱ型存活曲线的指数函数(取对数后为线性函数)的AIC 统计量最小(14.57),R2最大(0.86)。这说明在本研究调查的胸径范围内,九龙山香果树在各生长阶段的死亡率较稳定。
2.2 香果树的空间分布格局
全局适合度检验表明,香果树的空间分布格局显著偏离完全空间随机的零假设(P< 0.05)。具体来看,在距离小于4 m 时,香果树呈聚集分布;距离在4 ~ 15 m 时,香果树呈随机分布(图2A)。
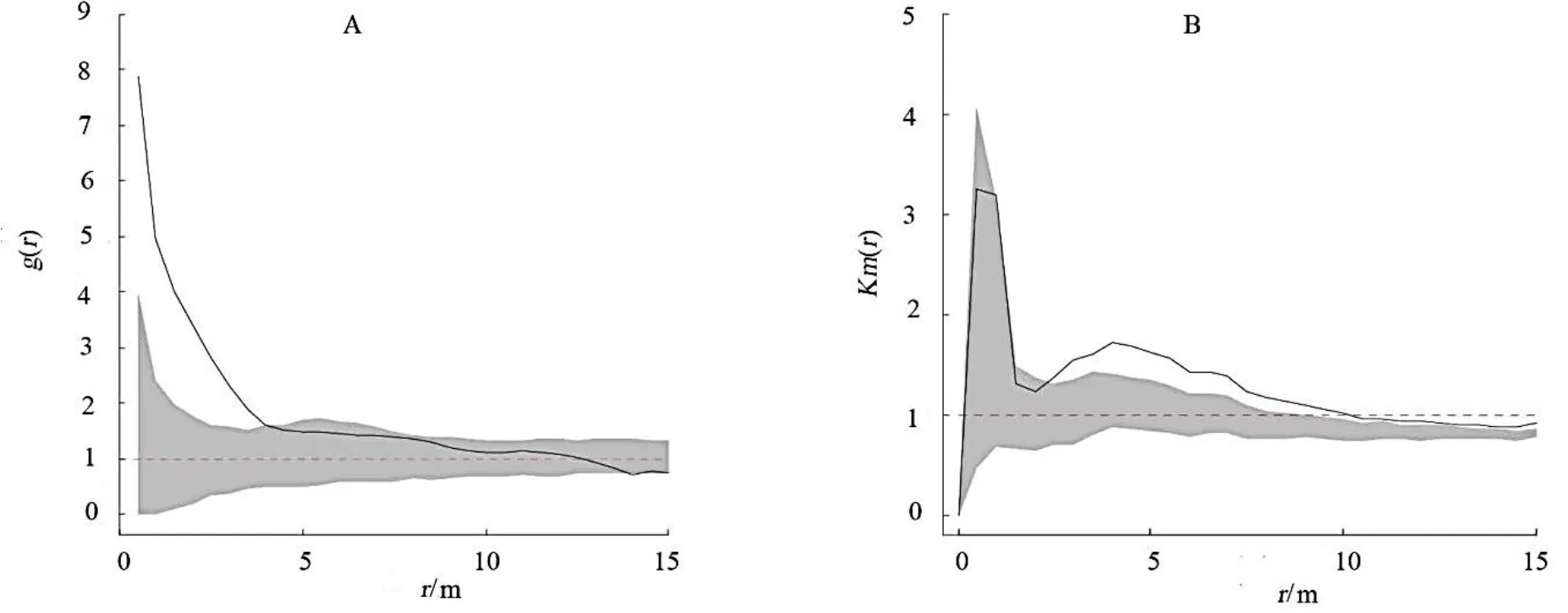
图2 九龙山香果树的空间分布格局和香果树胸径的空间关系Fig. 2 The spatial relation between distribution pattern and DBH of E. henryi in the Reserve
全局适合度检验表明,香果树的胸径在空间上显著偏离胸径随机标签的零假设(P< 0.05)。具体来看,在0 ~ 3 m 距离内周围香果树与中心香果树的胸径无显著关系,而分布在距中心香果树3 m 以上的其它香果树的胸径与中心香果树的胸径近似(图2B)。这说明分布在距离中心香果树3 ~ 15 m 范围内的香果树多来自同生群(即为中心香果树的同龄树);而在3 m 以内各年龄段的香果树均匀混合。
2.3 香果树与其它种类伴生树的空间关系
全局适合度检验表明,中心香果树与其它种类伴生树的空间关系显著偏离完全空间随机的零假设(P< 0.05)。具体来看,在5.5 ~ 15 m 的距离上,中心香果树与其它种类伴生树的种间关系主要为相互聚集;但在距离小于5.5 m 时,中心香果树与其它种类伴生树的空间分布格局基本是相互独立的(图3A)。
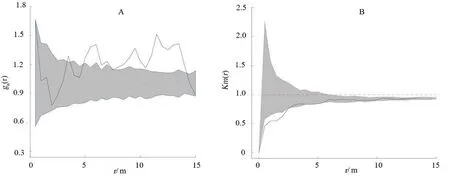
图3 中心香果树与其它种类伴生树个体的空间关系(A)以及中心香果树和其它种类伴生树胸径的空间关系(B)Fig. 3 The spatial relation between the central tree of E. henryi (A)and associated trees and with DBH of associated trees in the Reserve(A)
全局适合度检验表明,中心香果树与其它种类伴生树的胸径空间关系显著偏离胸径随机标签的零假设(P<0.05)。具体来看,在3 m 以上的距离,其它种类伴生树的胸径与中心香果树的胸径大小在空间上是相互独立的,而在3 m 范围内其它种类伴生树的胸径与中心香果树的胸径差异较大(图3B)。这说明在3 m 范围内较大中心香果树附近只存在较小的其它种类伴生树;而较小的香果树附近有较大的其它种类伴生树存在。
进一步对中心香果树与不同径级的其它种伴生树的空间关系进行分析表明,中心香果树与伴生的其它种小树、中树和大树在不同距离上存在不同程度的相互聚集,全局上显著偏离完全空间随机的零假设(P< 0.05)。具体来说,中心香果树与伴生的其它种小树在5.5 ~ 14 m 距离时,空间分布格局为相互聚集,在大于14 ~ 15 m时相互独立,在小于5.5 m 距离时,中心香果树与伴生的其它种小树的空间分布为相互聚集和相互独立的交替出现(图4A);中心香果树与伴生的其它种中树的空间关系在距离小于6 m 时相互独立,在距离大于6 m 时为相互聚集和独立交替出现(图4B);中心香果树与伴生的其它种大树的空间关系在距离小于5 m 时相互独立,在距离大于5 m 时相互聚集和相互独立的交替出现(图4C);中心香果树与伴生的其它种超大树的空间关系是相互独立的(图4D)。
3 讨论
3.1 九龙山香果树的种群动态
植物的径级结构不仅反映出当前种群的特征,更反映了未来的变化趋势[18]。本研究的野外调查发现香果树种群A 径级的个体仅1 棵,其胸径已达5.9 cm。虽然本次调查的起测胸径是2.5 cm,但我们在野外调查时并没有在样地中发现胸径小于2.5 cm 的香果树实生幼苗和幼树,根萌的幼苗和幼树也很少。因此,从整个生活史阶段来看,九龙山的香果树种群为衰退型,林下自然更新极其困难。这与康华靖等在包含九龙山的浙西南主要保护区得到的研究结果一致[19]。由于香果树种群开花不同步、花期中雨季较长、缺少传粉者和雌蕊柱头易受干扰等原因,香果树结实率非常低[20]。郭连金等的研究表明,香果树在达到23 年生后进入成年期,花期长短和生殖构件量随香果树树龄增长而增加[21]。从九龙山香果树种群的径级结构来看,大树很少。根据前人提出的香果树胸径与树龄的关系模型估算[2,21],九龙山香果树种群的平均树龄在23.7 ~ 27.5 年之间,大部分的香果树在10 ~ 30年之间。这说明九龙山香果树种群的繁殖母树并不多,且大多数能繁殖的母树还未达到繁殖最佳年龄(80 ~ 110年)。年轻香果树的花期相对较短,授粉成功的可能性降低。因此,种子繁殖能力较低可能是目前九龙山香果树种群更新困难的重要原因之一。从九龙山香果树种群的Ⅱ型存活曲线看,香果树胸径达到5.9 cm 以后其存活率比较稳定。现存大多数香果树今后应该能够顺利达到性成熟阶段,参与有性繁殖。根据郭连金等的研究结论[21],随着九龙山香果树种群大树年龄的增长,其母树数量和繁殖力都将增加。这是有利于九龙山香果树种群恢复的因素。
九龙山香果树种群的径级结构在一定程度上反映了九龙山自然保护区的历史。九龙山在1983 年成为省级自然保护区,在2003 年升为国家级自然保护区。九龙山胸径大的香果树植株数量较少,可能与在九龙山成为保护区之前,香果树遭到农业生产等人为破坏有关。随着保护区的建立,香果树种群开始逐渐恢复,因此大部分香果树的年龄在10 ~ 30 年之间。然而从2010 年开始,森林完全郁闭以后,香果树就开始更新困难,因此A 径级的香果树极少。有研究表明在受到中度干扰的不稳定生境中,香果树种群会表现出周期性零星更新的模式[3]。然而,香果树幼苗存活受岩石裸露度、郁闭度(或乔灌木盖度)、大气温湿度、群落内优势种群的生长状况和人为破坏程度等多方面因素影响[22-23]。郑伟成等[24]曾在九龙山大岩前、上廖坑和内阴坑等地发现81 株香果树幼苗。本次调查中,这些幼苗均未见到。这说明周期性零星更新的香果树幼苗极易夭折。
3.2 香果树空间分布及其与其他伴生种的空间关系
本研究通过g(r)函数分析表明,随着距离的增加,九龙山香果树种群逐渐由聚集分布变为随机分布。这与康华靖在大盘山的研究结果一致[25]。生境异质性和扩散限制是造成种群聚集分布的主要因素[26]。由于树木具有较大的根系和冠幅,小尺度上的生境异质性对树木生长和存活的影响较小,树木可以通过内部生理整合消除小尺度异质性对自身的影响。在森林中,较大尺度的生境异质性常使树木形成较大的聚集斑块,而扩散限制常可以使树木在小尺度上形成聚集格局。香果树喜分布于岩石裸露度高、土层浅薄的山谷林中,对生境有一定的要求。在总面积为5 525 hm2九龙山国家级自然保护区内,香果树仅在大岩前等9 个地点建立了居群。这显然是受保护区内生境异质性影响造成的。然而,本研究的调查直接针对九龙山现有的香果树分布居群展开,所设样圆的半径为15 m。在这一空间范围内,九龙山香果树种群主要在0 ~ 4 m 的距离上发生聚集,这是受繁殖体扩散限制影响造成的。九龙山香果树的实生苗更新能力有限,现有香果树种群中的小树和中树中的很大部分可能是通过萌生苗繁殖的。萌生繁殖极大地限制了香果树的扩散。另一方面,香果树种子的扩散能力也不强。香果树种子带翅且千粒重较低,是典型的风力传播的种子。一般认为靠风力传播的种子的扩散能力弱于靠动物传播的种子[26]。尤其是在密林中,靠风力传播的种子的扩散会受到树木枝叶的阻挡而难以远距离传播。扩散限制导致香果树子代(或萌生苗)常分布于母树周围。前人的研究普遍都显示香果树种群的空间分布呈现聚集特征,尤其在幼苗、幼树阶段聚集强度较高;随着龄级上升,香果树的分布格局逐渐从聚集分布过渡到随机分布[19,22,25,27-28]。在这一生态学过程的影响下,香果树母树和小树在3 m 以内的小尺度上聚集;同时在4 ~ 15 m 的距离上,香果树常有同龄香果树伴生,但大树缺少子代树伴生。香果树的这一空间分布特性将导致香果树种内竞争随着径级的增大逐渐增大。来自浙江大盘山的香果树种内竞争分析的结果验证了这一预测[29]。激烈的同生群内竞争可能是前人在多地发现随着径级增大香果树分布格局逐渐变得均匀的重要原因。
种间空间关系分析在各个距离上都未检测到中心香果树与其他种类伴生树间的空间排斥现象。在5.5 ~ 14 m的距离上,中心香果树与其它种类伴生树的空间关系为相互聚集。这说明作为干扰生境的开拓者——香果树的定植有利于其他种类的树木在其附近5.5 ~ 14 m 的范围内生长。香果树一般喜欢长在砾石堆中,这些地方往往土层浅薄、土壤贫瘠。香果树的生长促进了土壤层的发育,从而为其它树种幼苗和幼树的生长提供了良好的环境。进一步从其他种类不同径级的伴生树与中心香果树的空间关系来看,中心香果树周围2 m 以内、4 m 和5.5~ 14 m 的距离上聚集了较多的其它种类的小树,在5 ~ 7 m、8 ~ 10 m 和11 ~ 14 m 距离上聚集了较多的其它种类中树,在5 ~ 6 m、9 ~ 10 m 和12 ~ 13.5 m 距离上聚集了较多其它种类大树。野外研究发现在林窗和母树冠缘生境中香果树实生苗的光合生理表现最好[30]。大量的其它种类伴生树在香果树周围5.5 ~ 14 m 范围(即冠缘附近)聚集,阻碍了该区域内香果树的后代小苗获得充足的光照,从而使香果树小苗无法在其母树附近更新。另外,可能是由于存在激烈的种间竞争,本研究的结果显示其它种类的大树并未在中心香果树周围聚集。与康华靖在大盘山的研究结果相似[29],在九龙山随着伴生树胸径增大,其与中心香果树的空间关系趋向于独立。由于较大树木带来的竞争压力较大[29],在3 m 范围内较大中心香果树附近只存在较小的其它种类伴生树;而较小的中心香果树附近才会有较大的其它种类伴生树存在。
4 结论
总的来说,目前九龙山香果树种群幼树和小树很少,存在更新困难。香果树胸径达到5.9 cm 以后存活率比较稳定,存活数量随着径级增长而稳定下降。香果树繁殖体扩散能力有限,种群在小尺度(0 ~ 4 m)上聚集分布明显。分布在距中心香果树4 ~ 15 m 的其它香果树的胸径大小与中心香果树相似,存在同生群内竞争。香果树的生长改善了立地土壤环境,在中心香果树周围5.5 ~ 14 m 的距离上聚集了较多其它种类的小树、中树和大树(DBH < 37.5 cm)。这严重阻碍了扩散能力有限且喜光的香果树幼苗在母树附近生长和存活,加剧了香果树更新困难。今后如果能适当清理香果树周围的伴生树,降低乔、灌层盖度,加强对香果树幼苗和幼树的管理保护,改善香果树种子的萌发和幼苗发育条件,将极大地促进九龙山香果树种群的恢复。
本研究仅从径级结构推断了香果树的种群动态,从香果树空间格局及其与伴生种的空间关系推断了它们的种间和种内互作。今后应进一步从香果树的生长、开花结实、种子萌发和幼苗存活等角度对香果树的种内、种间互作和种群动态开展长期监测研究,要深入理解造成香果树濒危的群落学机制。
致谢:感谢九龙山自然保护区潘成椿和周世旺同志、丽水学院李文和柏正伟同学在野外调查过程中提供的支持和帮助。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
广西林业科学(2021年6期)2022-01-21
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
中华戏曲(2020年1期)2020-12-04
中华戏曲(2020年1期)2020-02-12
甘肃农业(2017年12期)2017-07-25
林业勘察设计(2017年4期)2017-07-06
西北植物学报(2015年9期)2015-07-05
