
基于形态和分子的越南蜉属(蜉蝣目:越南蜉科)一新种
——百山祖越南蜉
2023-12-13 08:36张立华黄海明刘雷雷程宏毅章书声
浙江林业科技 2023年6期
张立华,黄海明,刘雷雷,程宏毅,章书声,5
(1. 泰顺县自然资源和规划局, 浙江 温州 325500;2. 浙江省舟山林场,浙江 舟山 316021;3. 浙江乌岩岭国家级自然保护管理中心,浙江 温州 325500;4. 浙江师范大学 生命科学学院,浙江 金华 321004;5. 浙江省野生动物生物技术与保护利用重点实验室,浙江 金华 321004)
1997 年,McCafferty 和Wang 建立晚蜉科Teloganodidae 澳洲小蜉亚科Austremerellinae,包括越南蜉属Vietnamella[1],2000 年,又将该亚科提升为科,命名为越南蜉科Vietnamellidae[2]。目前,全世界报道的越南蜉科物种共有9 种,其中有效物种为6 种,中国分布4 种[3-10]。Hsu[11]将采自江西上饶的中华越南蜉命名为Ephemerellasinensis,而McCafferty 和Wang 将其移入越南蜉属,命名为Vietnamellasinensis[6]。1972 年,Tshernova建立新属越南蜉属并报道采自越南的韩氏越南蜉V.thani[3];同时报道的越南蜉Ephemerellaornata也被McCafferty 和Wang[6]移入越南蜉属并命名为装饰越南蜉V.ornata。1987 年,尤大寿和苏翠荣[4]描述了采自安徽和浙江的新种大别山越南蜉V.dabieshanensis。1995 年,周长发和苏翠荣[5]描述了采自浙江和福建的两新种——庆元越南蜉V.qingyuanensi和挂墩越南蜉V.guadunensis。2017 年,Hu 等[10]将之前的越南蜉属的大别山越南蜉、庆元越南蜉和挂墩越南蜉合并到中华越南蜉Vietnamellasinensis,认为这三个物种均为中华越南蜉的同物异名。近年来,随着形态和线粒体COI基因结合为蜉蝣目Ephemeroptera 新种发现提供佐证[8-9,12-14]。2020 年,Auychinda等发表采自泰国的新种V.nanensis[8];Luo 等发表采自车八岭国家级自然保护区的新种车八岭越南蜉V.chebalingensis[9];2021 年,Tong 等发现庆元采集的越南蜉为中华越南蜉的隐存种[15]。
蜉蝣作为原始的有翅昆虫,其系统发生关系一直是研究的热点[16-18]。小蜉科Ephemerellidae、越南蜉科和晚蜉科Teloganodidae 的系统发生关系存在一定的争议[19-23]。2005 年,Ogden 等[19]利用形态数据认为小蜉科、越南蜉科和晚蜉科三者的亲缘关系较近:越南蜉科与晚蜉科形成姐妹群,越南蜉科和晚蜉科支系再与小蜉科形成姐妹群。2009 年,Ogden 等[20]结合12SRNA、16SrRNA、18SrDNA、28SrDNA和H3基因的分子数据集与形态数据认为小蜉科、越南蜉科和晚蜉科等为并系关系,未有效得出三者的系统发生关系。之后,Cai 等[21]、Gao 等[22]和Ye 等[23]基于蜉蝣目线粒体13 个蛋白编码基因构建的系统发生树均支持小蜉科与越南蜉科形成姐妹群,而两者与晚蜉科关系较远。
线粒体基因(mtDNA)因其组成稳定、基因排列相对保守、普遍为母系遗传、进化速率比核基因快、极少发生重组等特点,故被广泛应用于进化、系统发生、种群遗传结构及系统地理学等方面的研究[15,24-30]。而线粒体COI基因作为DNA 条形码,能有效地进行物种鉴定,是研究物种遗传结构、种群多样性和隐存种的有效分子标记[30-31]。本研究拟对淳安、桐庐、庆元和泰顺四县的越南蜉属物种通过线粒体COI基因确定是否存在隐存种,并通过光学显微镜和扫描电子显微镜对不同地区的越南蜉稚虫口器、鳃、足等形态特征进行比较,在显微结构和超显微结构上研究其形态的差异,并证明为新种。
1 材料与方法
1.1 样本的采集与初步鉴定
越南蜉物种样本的采集地参考周长发等的方法[32],分别在2019 年7 月和2022 年7 月通过踢网法采集越南蜉稚虫,在浙江省淳安县(CA)、桐庐县(TL)、庆元县(QY)和泰顺县(TS)分别采集到8 只、5 只、15只和10 只稚虫。在尼康SMZ-1500 体视解剖镜下对采集到的越南蜉稚虫样本进行形态上的初步鉴定,参照《中国蜉蝣概述》[33]中的蜉蝣物种名录对样本进行分类,并做拍照记录。
1.2 形态特征观察
采用尼康SMZ-1500 体式解剖镜对越南蜉稚虫样本包括足、头、触角、下颚、下唇、舌、上唇、上颚、鳃及尾丝等形态结构进行解剖观察并拍照记录。用无齿小镊子将经过100%乙醇脱水3 次制备好的样品粘在导电胶上,用Phenom Pro 扫描电子显微镜进行扫描观察,选取结构完整、成像效果较好的部位进行拍照保存。使用Photoshop 软件对光镜、电镜下的越南蜉样本各形态结构数据进行测量统计并界定各采集地越南蜉的形态差别。
1.3 分子数据研究
1.3.1COI基因序列的获得 对4 个采集地各取2 个样本的胸部肌肉,使用Ezup 柱式动物基因组DNA 抽提试剂盒(生工生物工程(上海)股份有限公司生产)抽提总DNA。使用相应的COI基因引物[34]在PCR 仪(Bio-Rad,美国)中对提取的DNA 进行聚合酶链式反应(PCR)扩增,送生工生物工程(上海)股份有限公司进行双向测序。将测序成功的COI基因序列在NCBI 上进行BLAST 比对,确定是否为目的基因。对所获得的8 条COI基因序列比对,序列完全一致合并为相同单倍型,最终获得4 个采集地越南蜉的4 个单倍型COI基因序列。
1.3.2 遗传距离的计算和系统发生树的构建 在MEGA 7.0 中使用Clustal W 程序对NCBI 数据库中的越南蜉属物种(6 种14 条序列)与本研究越南蜉样本的COI基因序列(4 条单倍型序列)进行比对,而后在MEGA 7.0中选择K2P 模型分别计算越南蜉属各物种COI基因的遗传距离[35-36],并构建NJ 系统发生树。
2 结果与分析
2.1 形态特征
从庆元县和泰顺县采集的越南蜉物种稚虫(雌,图1A)(酒精中)主要特征:体长11.2 ~ 11.5 mm,尾丝11.0 ~ 12.0 mm,体色棕色至棕褐色,头部具突起和瘤突,边缘有密集的细毛。
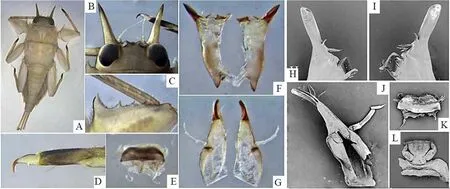
图1 采自庆元县和泰顺县的越南蜉稚虫的形态结构Fig. 1 Morphological structure of naiad of Vietnamella from Qingyuan and Taishun
头部(图1B):头部具额颊背突起,前部具2 对角状突起,内侧1 对较小,外侧1 对发达,略向下弯;触角着生于内外侧角之间,略长于外侧大角,13 ~ 17 节不等,节间具稀疏环细毛;复眼突出、圆形。
口器(图1E-L):上颚(图1F & H-I),细长,切齿发达,外切齿完全融合,呈匙状,尖细,暗红色,内切齿转化为两簇具有共同基部的刚毛,磨齿块状,表面粗糙;左右上颚不对称,右上颚(图1H)磨齿内侧有一较长的指状突起,内缘附加小簇刺状刚毛;下颚(图1G & J)端部尖细,端部内侧具2 ~ 3 个较大的刺状突起,外侧有一小簇刚毛;下颚须三节,有微小的刚毛,从基部到顶端的长度比为1.0∶(1.5 ~ 1.6)∶1.0;舌分三叶,两侧叶发达,端部和边缘具细毛;上唇(图1E & K)棕褐色,中部有一排横向排列的微刺,下缘及侧下缘密生细毛,侧缘细毛更长更密;下唇(图1L)上缘密生细毛,侧唇发达,中唇舌较小,下唇须3 节,每节均有稀疏微小的刚毛,基部比第二节宽和长,顶端非常小。
胸部(图1A):前胸背板的前侧有圆形小突起、前角向前呈刺状突起。每节胸部各具一对足,前中后足的腿节:胫节:跗节分别为3.5∶3.0∶1.0、(3.5 ~ 3.7)∶3.0∶1.0、4.5∶3.0∶1.0;前足腿节宽扁,其内缘有一排5 ~ 10 枚利齿,由近基部到远基部逐渐变小(图1C),胫节和跗节正常、无突起利齿,中后足正常;前足腿节最宽处形成明显的横向脊背,内、外、基部边缘和脊具一排刚毛,中后足腿节内、外、基部边缘有刚毛,各足胫节表面具细毛,内缘有一排密集的刚毛;各足的跗节呈深褐色,具非常稀疏的刚毛;各爪子均相似,有一个爪底小齿和一些细小的顶端刚毛,尖端(图1D)棕红色。
腹部(图1A):腹部背板灰褐色;腹面颜色稍浅,两侧具褐色纵纹;腹节背板的中部具矮的成对纵脊;1 ~9 腹节的侧后角先后延伸呈尖锐的刺状。腹部侧位具鳃7 对,位于第1 ~ 7 腹节;鳃方形,第1 对鳃较小,手指状,不分节,明显不同于其他6 对鳃;第2 ~ 6 对鳃结构相似,分背侧和腹侧片层;背叶膜质,腹叶分成两叉状,每叉又分成多个小叶;第7 对鳃较小,通常被第6 对鳃覆盖,也有2 叶,但腹叶不分叉,仅分成2 ~ 3 个裂叶。尾丝3 根,中尾丝长于另两根尾丝;尾丝棕褐色,其上具密集细毛。
经形态鉴定发现从淳安县采集的中华越南蜉与桐庐县采集的中华越南蜉一致,而从庆元县和泰顺县采集的越南蜉样本在稚虫期可通过以下特征与从淳安县和桐庐县采集的中华越南蜉区分:①口器的上、下颚切齿尖端更为细长;②头部两个大而直的角状突起夹角较中华越南蜉的小;③腿节相对较宽,尤其是中后足的腿节。其中细长尖锐的上颚外切齿和下颚顶端为鉴别庆元县和泰顺县采集的越南蜉样本重要形态特征。从庆元县和泰顺县采集的越南蜉样本跟其他越南蜉物种可以用下颚须从基部到顶端的长度比明显区分开,如车八岭越南蜉V.chebalingensis和V.nanensis下颚须从基部到顶端的长度比为1.0∶1.3∶1.0;V.maculosa下颚须从基部到顶端的长度比为1.3∶1.2∶1;韩氏越南蜉V.thani下颚须基部节与第二节几乎等长。而装饰越南蜉V.ornata因至今未有稚虫报道,无法从形态特征进行区别,但可以从遗传距离进行区别。
2.2 遗传距离
从淳安县采集的中华越南蜉与从桐庐县采集的中华越南蜉的遗传距离为0.002,两者与NCBI 数据库的中华越南蜉(HM067837)的COI基因遗传距离分别为0.042 和0.044,小于昆虫物种种间差异范围[37];而从庆元县和泰顺县采集的越南蜉样本的COI基因与NCBI 数据库的中华越南蜉(HM067837)的COI基因遗传距离分别为0.097 和0.099,达到种上水平。Williams 等利用COI基因发现不同地理种群的Baetisrhodani遗传距离达0.08~ 0.19,推测部分遗传距离大于0.08 的种群存在隐存种[38]。本研究中从庆元县(QY)和泰顺县(TS)采集的越南蜉样本的COI基因与越南蜉属已知物种的遗传距离达0.097 ~ 0.220,大于昆虫物种种间遗传距离,支持其为新种,命名为百山祖越南蜉Vietnamellabaishanzuensissp. nov. Zhang, Huang, and Zhang, 2023。
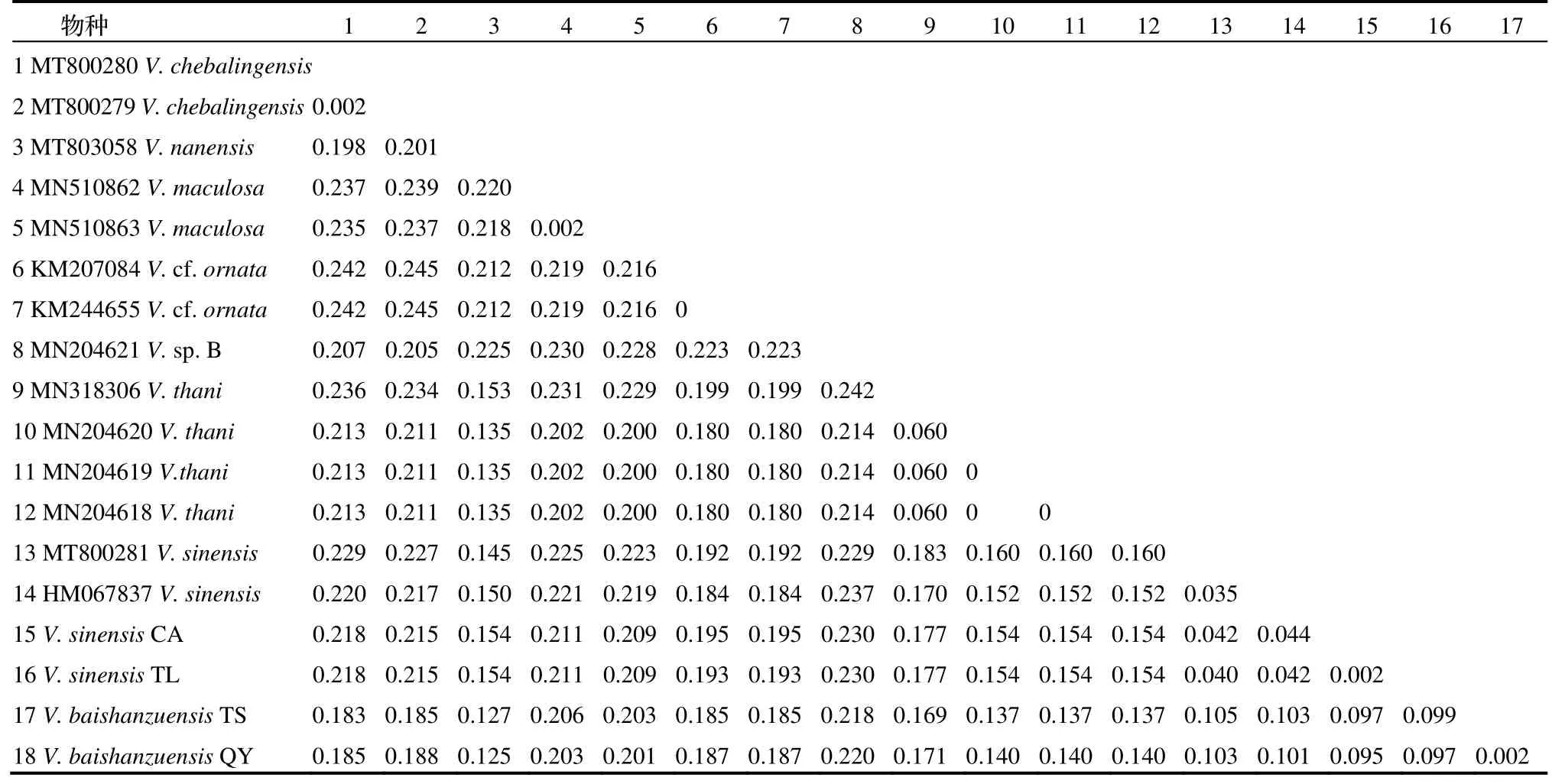
表1 基于K2P 模型的越南蜉属物种间的遗传距离Tab. 1 Genetic distance between Vietnamella species based on K2P model
2.3 系统发生关系
基于越南蜉属8 种18 条线粒体COI基因所构建的NJ 系统树(图2),结果支持新种百山祖越南蜉与中华越南蜉互为姐妹群关系,该(百山祖越南蜉+中华越南蜉)支系再与(韩氏越南蜉蝣V.thani+V.nanensis)支系形成姐妹支系;而装饰越南蜉V.ornata与蓝纹越南蜉V.maculosa形成姐妹群关系。Vietnamellasp. B 为所有已知越南蜉属物种的基部,该结果与Luo 等的结果一致[8-9]。但其他蜉蝣属物种的相互关系存在差异,有待进一步研究。本研究中虽然百山祖越南蜉与中华越南蜉形成姐妹群关系,但根据其遗传距离差异,以及形态特征差异,均支持百山祖越南蜉为新种。
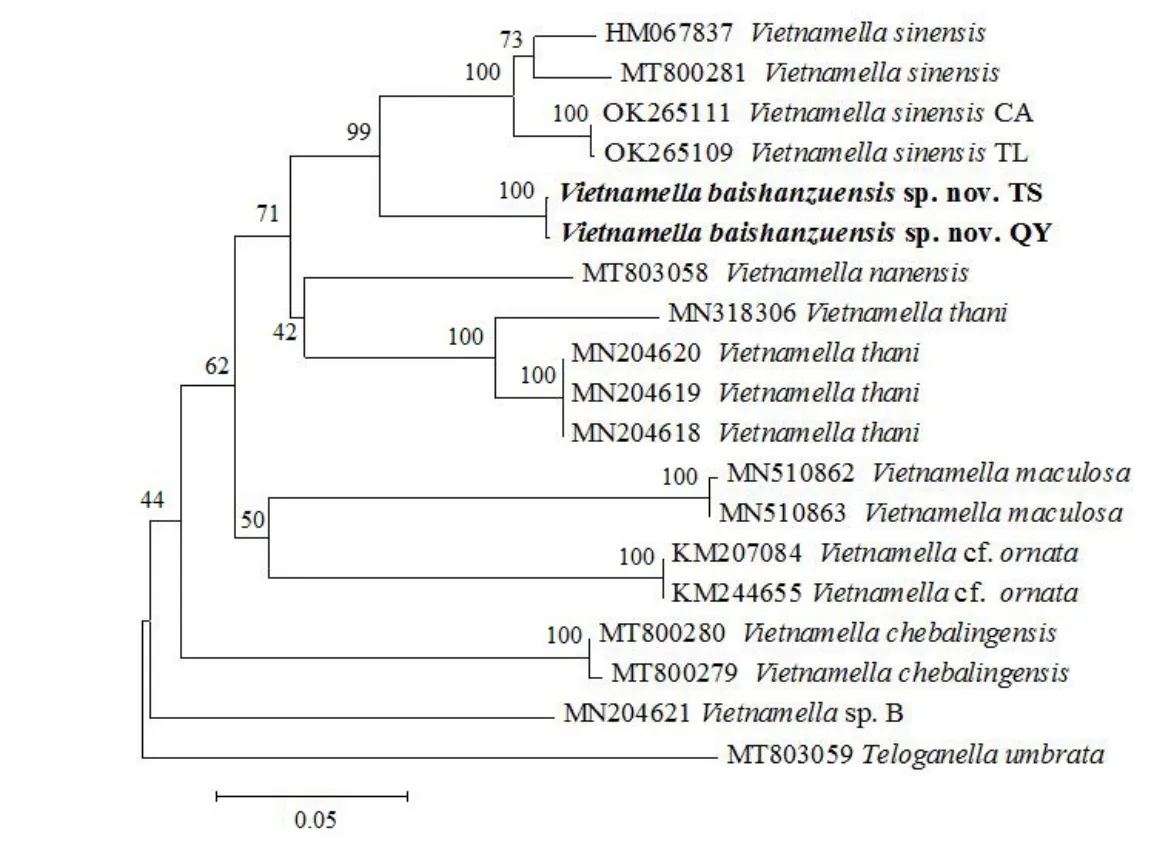
图2 基于线粒体COI 基因构建的NJ 系统发生树(粗体为新种百山祖越南蜉)Fig. 2 NJ phylogenetic tree constructed based on mitochondrial COI gene (bold words mean new species, V. baishanzuensis sp. nov.)
2.4 词源学
结合形态特征和分子数据,我们认为从庆元百山祖和泰顺大溪源采集的越南蜉样本为新种,以正模模式标本采集地百山祖国家级自然保护区为种名词源,定名为百山祖越南蜉Vietnamellabaishanzuensissp. nov. Zhang, Huang and Zhang, 2023。
2.5 模式标本
正模1♀:浙江省庆元县百山祖国家级自然保护区,2019—07—02,章书声。副模:3♂♂,11♀♀,浙江省庆元县百山祖自然保护区,2019—07—02,章书声;2♂♂,5♀♀,浙江省泰顺县大溪源省级保护区,2022—07—22,张立华,章书声;3♀♀,浙江省泰顺县乌岩岭国家级自然保护区,2022—07—25,章书声。所有模式标本保存在浙江师范大学昆虫标本馆。
3 结论
综上所述,在越南蜉隐存种探讨中,通过形态和分子数据研究发现从淳安县和桐庐县采集的越南蜉与模式产地中华越南蜉为同一个种,而从庆元县百山祖和泰顺县大溪源采集的越南蜉为新种,词源学以正模模式标本采集地庆元百山祖为词源命名,定名为百山祖越南蜉。
猜你喜欢
浙江林业(2022年8期)2022-09-13
北京大学学报(自然科学版)(2022年4期)2022-08-18
今日农业(2021年3期)2021-12-05
学苑创造·A版(2021年9期)2021-09-16
食用菌(2021年4期)2021-07-29
食用菌(2021年3期)2021-06-03
科学导报(2020年50期)2020-09-09
中国篆刻·书画教育(2020年5期)2020-06-08
世界家苑(2018年2期)2018-04-28
环球人文地理·评论版(2017年4期)2017-07-16
