
蚁丘形成对高寒草甸植物群落多样性的影响
2023-12-04 06:48郭童童陈梦娇李兰平梁德飞
草地学报 2023年11期
郭童童, 刘 艳, 陈梦娇, 崔 靖, 李兰平, 梁德飞, 任 飞*
(1.青海大学省部共建三江源生态与高原农牧业国家重点实验室, 青海 西宁 810016; 2.青海大学生态环境工程学院,青海 西宁 810016; 3.青海三江源草地生态系统国家野外科学观测研究站, 青海 西宁 810016)
青藏高原(Qinghai-Tibetan Plateau,QTP)被称为“世界屋脊”,是全球重要的生态安全屏障[1],聚集了大量植物区系,而植物本身就对高山恶劣环境有高度特化的响应,因此青藏高原植物种类丰富多样,同时它亦是全球生物多样性的敏感地区,保护植物多样性是维护当地生态系统稳定的重点[2]。高寒草地是青藏高原极为关键的生态价值系统,面积可达1.60×106km2,占整个青藏高原总面积的60%,主要包括高寒草甸(Alpine meadow)和高寒草原(Alpine steppe)两种植被类型[3]。而作为青藏高原较为典型的植被类型,高寒草甸生态系统具有典型的地带性分布特征,其生态系统功能较脆弱,抗冲击能力差,对干扰因子的响应较为敏感[4]。高寒草甸生态系统蕴含十分丰富的水土资源以及巨大的昆虫资源库[5],这当中包括诸如蚂蚁,跳虫等土壤节肢动物(Soil arthropods),土壤节肢动物往往在生态系统中担任着关键的角色[6],在维持物质循环和能量流动过程中起着重要作用[7],并且进入新时代以来,越来越多的学者将生态系统功能和土壤节肢动物联系在一起研究[5-8]。这也说明土壤动物的研究进入一个新的阶段。其中蚂蚁就是一类典型的土壤节肢动物[9],尽管体型微小,却被称为“生态系统工程师”[10-11],其对生态系统有不可估量的影响[11]。有研究表明,蚂蚁是草地生态系统中不可忽视的生物因子[12],通过对蚂蚁活动的监测,可以反映出生物多样性的变化,科学家们还以蚂蚁作为生物指示物来评价生态恢复的效果[13]。
蚂蚁类属小型土壤节肢动物,隶属于昆虫纲(Insecta)膜翅目(Hymenoptera)蚁科(Formicidae),生活方式是社会性群居,在微环境尺度上进化较为成功[14],其在大多数生态系统中数量繁多,种类极其丰富,它们至少占所有昆虫生物量的三分之一,可能相当于人类的生物量[15],蚂蚁被认为是陆地生态系统中极具影响力的生物[16],并且蚂蚁和植物是作为草地生物多样性、生态系统服务和保护价值的指示物,蚂蚁类群和植物的多样性指数被证明为整个草原群落的良好生态指标[17]。同时草地是支持高水平生物多样性的重要的生态系统[18],而蚂蚁则作为草地上重要的消费者[9]、互惠保护者和生态系统工程师[19],它影响着植物的多样性[20]和潜在的草地生产力,研究蚂蚁和植物之间的相互关系,就成了当下新颖的研究着力点及热点[20]。这两个群落紧密联系,随着生境异质性的增加,资源的可利用性、筑巢的可能性和小气候条件也会增加,这反过来又可能支持更高的物种丰富性和功能多样性[21]。本研究则着重探究高寒草甸上蚂蚁和植物这两个生态重要指标之间的互作关系。
目前,蚂蚁对生态系统影响的研究在北温带草原[16]、热带雨林[22]、地中海栎林地[10,23]、松嫩草地[24-25]、欧洲半天然草原[26]、非洲稀树草原[27]、科尔沁流动沙地[28]和腾格里沙漠[29]等地域均有报道。如刘任涛等[28]详细研究阐述沙质生态系统中的掘穴蚁(Formiciacunicularia)蚁丘形成活动对植被结构的影响,还探讨了蚂蚁和沙地,土壤之间的紧密联系,这也为土壤动物在沙地生态系统的生态恢复作用提供理论支撑;相似的研究也曾在腾格里沙漠中的一个人工植被区进行,指出掘穴蚁筑丘在流沙固定过程中可改善周边土壤环境[29]。代军[24]则对于东北松嫩草原生态系统蚁丘筑巢的研究中阐明在不同程度的退化草地上,玉米毛蚁(Lasiusalienus)和黄毛蚁(Lasiusflavus)可以通过改变土壤理化性质间接影响植物群落结构,改变其生物多样性。而关于高寒草甸上蚂蚁对植物群落的影响未见系统性的报道。
基于此,本研究以中国科学院西北高原生物研究所青海海北高寒草地生态系统国家野外科学观测研究站(简称海北站)为依托,选择高寒草甸典型植被类群,旨在通过蚂蚁筑巢活动即蚁丘形成来探究对小尺度高寒草甸植物群落多样性的影响,探讨在蚁丘干扰下高寒草甸植物群落内部结构的变化规律,为综合评价和明晰蚁丘对高寒草甸生态系统的稳定、健康发展和功能影响提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于中国科学院西北高原生物研究所青海海北高寒草地生态系统国家野外科学观测研究站(简称海北站)附近的高寒草甸,地理坐标37°39′15.407″ N,101°19′54.200″ E,海拔约为3 231 m,属明显的高原大陆性气候,气温变幅大,寒流和冷空气侵入频繁,强度大,霜冻严重,全年无绝对无霜期,相对无霜期约20 d,在最热月的7月仍可出现霜冻、结冰、降雪等冬季天气现象[30],降水在7月和8月最多,1月和12月最少。降水量主要集中在5—9月暖季,约占年降水量的80%[31],蒸发量1 160.3 mm。夏季受东南季风气候而冬季受西北利亚寒流的影响,无明显的四季之分,暖季短暂而凉爽,冷季寒冷而漫长。每年的5月至9月为植物的主要生长季,植被类型是典型的以矮生嵩草(Kobresiahumilis)、高山嵩草(Kobresiapygmaea)为建群种的群落,群落优势种有垂穗披碱草(Elymusnutans)、羊茅(Festucaovina)、冷地早熟禾(Poacrymophila)、麻花艽(Gentianastraminea)、钝苞雪莲(Saussureanigrescens)、细叶亚菊(Ajaniatenuifolia)、甘肃棘豆(Oxytropiskansuensis)、披针叶野决明(Thermopsislanceolata)等植物。
1.2 研究方法
1.2.1样地设置 选择在2021年8月和2022年8月展开本研究的实验调查,样地在海北站西北方向约2 km位置处的高寒草甸,实验设置3个50 m×50 m的实验区(大样方),单个实验区划分成10个5 m×50 m的样带,重复且平行,年际间的实验区的选择是固定的。总计实验区内蚁丘数量,继而采用彩色旗帜标示蚁丘位置,使用无人机俯看蚁丘大致分布情况。设置标准,随机选择适合的蚁丘,分为小、中、大和废弃类型蚁丘。同时对蚁丘样地植物物种进行调查。采用样方框(100交叉点),样方框(0.5 m×0.5 m)中心点对准目标蚁丘的正中位置,全覆盖在蚁丘上,并用针刺法调查样方框内的植物物种组成,以样方框包含的所有植物物种组成为单个蚁丘界定的全部范围。同时将选择蚁丘进行底面积计算。并且选用一对一样方法进行对照比较。以蚁丘中心点为圆心向外扩展1~2 m,在目标蚁丘左上或者右下设置平行对照样方。两年间野外调查重复且平行。最终每一年选取蚁丘共32个即“大蚁丘”,“中蚁丘”,“小蚁丘”,“废弃蚁丘”各8个,同时对应平行对照样方32个,总计64个调查植物群落样方。
1.2.2测定指标及测定方法
(1)蚁丘的大小与数量
将蚁丘类型划分为小(S)、中(M)、大(B)以及废弃(A)蚁丘4个大类,具体分类的标准如下:
将蚁丘长轴作为计算标准,若测量长轴≤ 15 cm归为小蚁丘,15 cm <长轴≤30 cm归为中蚁丘,长轴> 30 cm归为大蚁丘。废弃类型的蚁丘多为不完整蚁丘,没有具体的大小。不纳入计量范围。除废弃蚁丘外其余类型蚁丘底面积投影可近似视为椭圆,使用钢卷尺分别测量蚁丘的长轴(a)和(b),最后使用椭圆面积公式确定蚁丘的面积,蚁丘面积S的计算公式为:
S=π×ab/4
(2)蚁丘植物群落调查
单个蚁丘,利用0.5 m×0.5 m的样方框(100交叉点)附在上面,保持样方框与蚁丘的长轴和短轴平行。样方框中心点与蚁丘中心相对应。样方框内含有100个小方块格,用针刺法记录在内的植物种类和数量。以目标蚁丘为中心,(1~2)m为半径范围边缘外做蚁丘的对照样方,与蚁丘一样进行群落调查。根据方精云等[32]对植物群落清查方式的指导,本文用相对盖度替代相对多度,采用植物群落的Alpha多样性[33]来解释植物群落结构,具体的指标如下:
①物种丰富度(Species richness,SR):SR=S
②Alpha多样性[33]:
Shannon-Winner群落多样性指数:H=∑Pi×lnS
Simpson优势度指数:D=1-∑Pi2
Pielou均匀度指数:J=(-∑Pi×lnS)/lnS
式中:Pi是样方中各物种i在群落中所占的重要值,重要值用群落中各物种的相对盖度指代;S为物种i所在样方的物种总数。
(3)植物功能群落划分
调查样地所有植被类型,得出植物共有17科42属60种。本实验样地根据植被调查分4大功能群(Plant functional groups,PFGs)即:①禾草类(Grasses),包含所有的禾本科植物(Gramineae plants)。②莎草类(Sedges),包含所有的莎草科植物(Cyporaceae plants)。③豆科类(Legumes),包含所有的豆科植物(Leguminosae)。④杂类草(Forbs),包含所有的杂类草植物(Weedy plants)。
1.3 数据分析
本论文的数据前期整理和初步计算在Excel 2019中完成。采用单因素方差分析和最小显著差异(LSD)多重比较分析,分析不同类型蚁丘对整体植物和各植物功能群落的影响。利用配对样本t检验,分析不同类型蚁丘面积的差异关系。统计分析利用SPSS 25完成,图像可视化采用Origin Pro 2021,Alpha多样性指数分析使用R 4.1.1。
2 结果与分析
2.1 蚁丘的分布格局及其蚁丘面积
大型蚁丘的数量在所有调查蚁丘中占据比例是最大的(表1),平均占比可达31%,这可能与做调查的季节有关,也说明在8月份时蚁丘的活动是比较旺盛的。计算得出蚁丘的平均密度为161个·hm-2。

表1 两年内不同类型蚁丘数量统计表Table 1 Statistics of different types of ant-mound in two years
本研究对两年间共64个不同类型的蚁丘面积进行计算。蚁丘的底面积范围在0.01~0.15 m2之间(图1),通过t检验得出,小蚁丘与中蚁丘存在显著的差异(P<0.05),大蚁丘与小蚁丘存在显著差异(P<0.05),中蚁丘和大蚁丘之间存在显著的差异(P<0.05)。
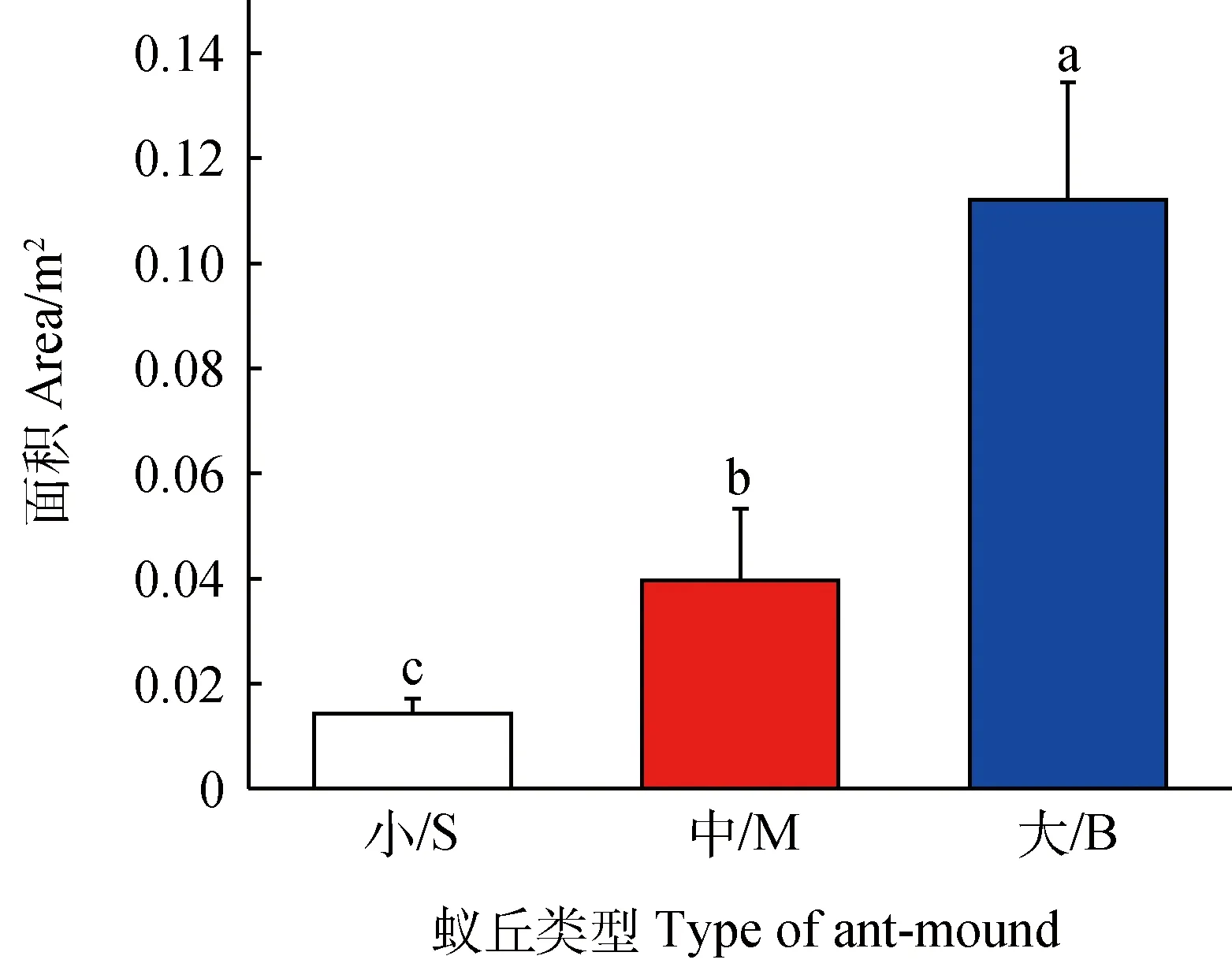
图1 不同类型的蚁丘面积Fig.1 Area of different types of ant-mounds.注:图中小写字母表示不同蚁丘类型面积的差异显著性(P <0.05)Note:Different lowercase letters in the figure indicate a significant difference in the area of different size of ant mounds at the 0.05 level
2.2 蚁丘形成对整体植物群落多样性的影响
蚁丘形成对整体植物群落多样性指数的单因素方差分析(表2)表明,蚁丘形成对植物物种丰富度有极显著的影响(P<0.001),蚁丘形成对植物香农-威纳指数有非常显著的影响(P<0.01),而蚁丘形成对植物辛普森优势度指数和均匀度指数均不存在显著影响。草地群落多样性由高到底的顺序依次为废弃蚁丘>中蚁丘>小蚁丘>大蚁丘(图2),而大蚁丘的物种丰富度指数和香农-威纳指数显著小于各样地和对照样方(P<0.05)。

图2 不同类型蚁丘的群落多样性指数Fig.2 The diversity indexes of different types of ant-mounds注:小写字母表示不同类型蚁丘之间的显著性差异(P<0.05),*符号表示蚁丘与对照样方的显著性差异,***表示P<0.001Note:Different lowercase letters indicate a significant difference between different types of ant-mounds at the 0.05 level,*** a significant difference between ant-mounds and controls at P<0.001

表2 蚁丘形成对整体植物群落多样性影响的单因素方差分析(ANOVA)表Table 2 One-way analysis of variance (ANOVA) table of effects of ant-mound formation on the overall plant community diversity
2.3 蚁丘形成对各植物功能群落多样性的影响
高寒草甸禾草科、莎草科、豆科和杂类草四个功能群随蚁丘面积的增加而变化。单因素方差分析结果(表3)表明,蚁丘形成对各植物功能群均有明显的影响,其中杂类草变化明显,各多样性指数均有极显著的水平(P<0.001)。

表3 蚁丘形成对各功能群的生物多样性指数的单因素方差(ANOVA)分析表Table 3 One-way analysis of variance (ANOVA) table of effects of ant-mound formation on plant functional groups’ community diversity
禾草类功能群多样性,随蚁丘面积变化而改变(图3)。禾草功能群物种丰富度指数由高到低,依次为中蚁丘>废弃蚁丘>大蚁丘>小蚁丘,中蚁丘显著大于小蚁丘;香农-威纳指数和辛普森指数由高到低,依次为中蚁丘>大蚁丘>废弃蚁丘>小蚁丘,而中蚁丘和大蚁丘显著大于小蚁丘;中蚁丘、大蚁丘物种丰富度指数、香农-威纳指数和辛普森指数显著大于对照样方;均匀度指数由高到低,依次为大蚁丘>中蚁丘>废弃蚁丘>小蚁丘,且中蚁丘和大蚁丘显著大于小蚁丘(P<0.05)。综上所述,中蚁丘、大蚁丘和废弃蚁丘增加了高寒草甸禾草类功能群多样性。

图3 不同类型蚁丘的禾草类功能群多样性指数Fig.3 Grass functional group diversity indexes of different types of ant-mounds注:小写字母表示不同类型蚁丘之间的显著性差异(P<0.05),*符号表示蚁丘与对照样方的显著性差异,*表示P<0.05,**表示P<0.01Note:Different lowercase letters indicate a significant difference between different types of ant-mounds at the 0.05 level,* a significant difference between ant-mounds and controls at P<0.05,** at P<0.01
随蚁丘面积的增大,莎草类功能群多样性呈现不同变化(图4)。莎草类功能群物种丰富度指数沿废弃蚁丘、大蚁丘、小蚁丘、中蚁丘的顺序依次降低,废弃蚁丘显著大于中蚁丘,大蚁丘与对照样方存在极显著的水平(P<0.001),且高于对照群落。香农-威纳指数和辛普森指数,沿大蚁丘、废弃蚁丘、小蚁丘、中蚁丘顺序依次降低,不同面积蚁丘和废弃蚁丘之间无显著差异,但小蚁丘显著大于对照样方(P<0.05),大蚁丘极显著大于对照样方(P<0.01)。均匀度指数沿小蚁丘、中蚁丘、大蚁丘和废弃蚁丘的顺序依次降低,小蚁丘显著大于对照样方(P<0.05),说明莎草类功能群多样性在不同面积蚁丘和废弃蚁丘上增大,呈增加的趋势。

图4 不同类型蚁丘的莎草科功能群多样性指数Fig.4 Sedge functional group diversity indexes of different types of ant-mounds注:小写字母表示不同类型蚁丘之间的显著性差异(P<0.05),*符号表示蚁丘与对照样方的显著性差异,*表示P<0.05,**表示P<0.01,***表示P<0.001Note:Different lowercase letters indicate a significant difference between different types of ant-mounds at the 0.05 level,* a significant difference between ant-mounds and controls at P<0.05,** at P<0.01,*** at P<0.001
豆科类功能群多样性低于对照样方(图5)。豆科类功能群物种丰富度指数中蚁丘>小蚁丘>废弃蚁丘>大蚁丘,香农-威纳指数、辛普森指数、均匀度指数小蚁丘>中蚁丘>废弃蚁丘>大蚁丘;大蚁丘极显著小于对照样方(P<0.001);大蚁丘除物种丰富度指数外,其他3种多样性指数显著小于各样地(P<0.05),表明豆科类功能群多样性随蚁丘面积增大,呈降低趋势,大蚁丘明显降低了豆科类群落多样性。

图5 不同类型蚁丘的豆科类功能群多样性指数Fig.5 Legume functional group diversity indexes of different types of ant-mounds注:小写字母表示不同类型蚁丘之间的显著性差异(P<0.05),*符号表示蚁丘与对照样方的显著性差异,**表示P<0.01,***表示P<0.001Note:Different lowercase letters indicate a significant difference between different types of ant-mounds at the 0.05 level,** a significant difference between ant-mounds and controls at P<0.01,***at P<0.001
杂类草功能群多样性低于对照样方(图6)。杂类草功能群物种丰富度、香农-威纳指数、辛普森优势度指数、均匀度指数表现为小蚁丘>废弃蚁丘>中蚁丘>大蚁丘,大蚁丘显著小于对照样方(P<0.001),且显著小于各样地(P<0.05)。说明杂类草功能群多样性在不同面积蚁丘和废弃蚁丘上均不同程度的降低,而大蚁丘降低最为明显。

图6 不同类型蚁丘的杂类草功能群多样性指数Fig.6 Forb functional group diversity indexes of different types of ant-mounds注:小写字母表示不同类型蚁丘之间的显著性差异(P<0.05),*符号表示蚁丘与对照样方的显著性差异,***表示P<0.001Note:Different lowercase letters indicate a significant difference between different types of ant-mounds at the 0.05 level,*** a significant difference between ant-mounds and controls at P<0.001
3 讨论
限制蚁丘密度的因素有很多[35],研究表明,蚂蚁的种类与蚁丘分布格局具有很大相关性[24],如在退化草地玉米毛蚁蚁丘在数量上总多于黄毛蚁蚁丘。本研究中的蚂蚁种类初步鉴定可能为北京凹头蚁(Formicabeijingensis)、草地铺道蚁(Tetramoriumcaespitum)以及光亮黑蚁(Formicacandida),而不同蚁群筑丘活动存在分异性,导致形成了面积不同的蚁丘,进而影响了草地植物群落的演替。而本研究将蚁群视作集合群体,以蚁丘面积对高寒草地植物群落影响为重点,探讨蚁丘面积对草地多样性的影响。熊忠平等[36]在祁连山国家公园青海片区蚂蚁物种多样性研究中表示蚁丘个体密度主要受气温和海拔制约,海拔越高,密度越低[36]。其次,地形的因素,不同的生态系统、不同的土地利用方式和不同的土壤组成等,这些因素都会在一定程度上影响蚁丘的分布格局[35]。而本研究各处理的海拔、地形、生态系统自然条件基本一致,不存在因环境因素而导致的差异性。蒙凤群等[37]对川西北的高寒草甸(无退化)蚁丘调查后,得出的蚁丘密度约为60个·hm-2。代军[24]在东北地区松嫩草原进行蚁丘密度的调查得出,轻度退化生态系统约为600个·hm-2,重度退化生态系统约为400个·hm-2。而对欧洲地区的草原进行蚁丘密度的统计后,得出约0.7个·m-2的结果[38],即7 000个·hm-2蚁丘。因此,国内外蚁丘密度是衡量草地退化的重要指标之一,本研究的蚁丘密度为161个·hm-2,略高于川西北无退化的高寒草甸蚁丘数量,而远低于东北地区松嫩草原和欧洲地区的草原退化草地蚁丘数量。相关文献表明,蚁丘密度直接影响蚁丘的大小和空间格局分布[28],而蚁丘的分布存在过分散分布[38-39]、聚集分布和随机分布等不同的模式,因此,目前对蚁丘分布格局的研究难以找寻规律。由于蚁丘大小和蚁丘的建立时间相关[40],所以蚁丘的面积变化可在一定程度上反映出蚁丘建立的时间演替序列。同时,不同类型的蚁丘往往会对蚁丘自身的理化性质产生不同的影响,如在一定的季节废弃蚁丘中央的温度可能会比大蚁丘更大[41]。
本研究中,大蚁丘的干扰程度在整体群落和各个功能群中尤为凸显,说明随蚁丘面积增大,蚁丘提供异质性微环境特征越来越明显。这为本来在群落中不具备生态优势的植物提供了条件,并使得某些植物“适宜”生存,这与川西北高寒草甸的研究一致。本研究发现,废弃蚁丘在整体和各植物功能群落上和平地(对照)在各项多样性指标上均没有显著差异,甚至极为接近,这说明蚁丘废弃后干扰行为结束,草地具备自我修复的效果,有能力经过一系列自然演替阶段返回到原生植被状态,这和周华坤等[42]对海北高寒草甸自然恢复能力的研究一致。同时,蚁丘形成会对整体植物群落多样性造成降低的影响,蚁丘干扰下的禾草类和莎草类植物功能群在各个指数水平上均呈现增加的趋势,在蚁丘演替中,大类型时均高于平地对照;豆科类和杂类草植物功能群在蚁丘干扰下呈现的是下降趋势,且在不同类型蚁丘均低于对照。这表明豆科类和杂类草的减退趋势大于禾草科与莎草科的增升趋势。其中杂类草功能群响应非常显著,大量杂类草减退,可能的原因是蚁丘建立的初步阶段会搬运土壤,腐殖质和土壤养分增加,特殊的理化性质将大部分矮小,茎软的不耐土埋杂类草植物埋掩窒息致死亡[43],禾草类,莎草科等质硬叶尖直立型或较高的耐埋植物存活下来,并逐渐占据生存优势,也有研究表明蚂蚁和其喂养的蚜虫分泌物会损害植物根系致使植物死亡[44],蚁丘上嵩草、禾草植物在受到干扰时蚂蚁能迅速爬上茎秆,保护蚁丘上的类禾草免受食草动物的采食[37]。值得一提的是,莎草科属高寒草甸建群种[45],在蚁丘干扰下表现为更“适宜”的生长模式。蚁丘上的植物群落逐渐转向禾草类和莎草类为优势物种的方向演替,这与向泽宇等[46]在四川省高寒草甸蚁巢群落和赵敏[47]在青藏高原东北缘高寒草甸的做出的相关研究结论一致。
本研究结果最终体现为蚁丘干扰降低植物群落的多样性。赵敏[47]的研究结果表明,蚂蚁筑巢行为会降低群落物种丰富度和辛普森指数,但是会提高均匀度指数和香农-威纳指数。代军[24]对松嫩草地中的黄毛蚁和玉米蚁的蚁丘研究结果表明,蚁丘的形成并不会改变植物群落的丰富度指数、香农-威纳指数以及均匀度指数,一样的研究结果也出现在国外的温带草原生态系统中[48],侯继华[12]在松嫩草地中做出纯羊草群落多样性高于对照的研究,刘任涛等[28]在科尔沁沙地的研究中表明掘穴蚁筑巢会间接提高植物群落多样性,以上与本研究结果不一致,可能的原因是生态系统、蚂蚁种类和气候的差异性所致,气候和地形因素是影响物种多样性格局的两类重要因素[49]。赵一军等[50]研究中表明蚂蚁筑巢活动降低植物多样性,改变植物群落结构,提升微环境异质性;胡思雨[35]在高寒草甸的研究表明蚁丘筑巢会降低植被群落多样性指数,这与本研究结果一致。由此看来需要对蚁丘、植被类型、生态系统特征之间的相互关系进行更多的研究[51]。
4 结论
研究表明,蚁丘的初步分布特征为随机分布,也伴有均匀分布或分散分布格局。但本研究高寒草甸区蚁丘以随机分布为主,将蚁丘面积大小作为试验变量,经统计分析,各样地蚁丘蚁丘面积在0.01~0.15 m2之间,并未有逾越范围的超大面积蚁丘出现。本研究结果表明,不同面积蚁丘上高寒草甸植物群落多样性存在分异性,说明不同面积蚁丘形成干扰了草地群落演替。蚁丘面积的增大对禾草类和莎草类植物类群多样性有明显增强的影响,而对豆科类和杂类草植物多样性有明显减弱的影响,杂类草多样性变化较为敏感;伴随蚁丘面积的增大,大量的杂类草多样性持续减少,在大蚁丘上体现尤为明显;但当蚁丘被废弃后干扰结束,高寒草甸能自我恢复到天然草地水平。因此,随着蚁丘面积的增大,减弱豆科和杂类草植物的生存优势,蚁丘上植物群落向着禾草类和莎草类为优势种群的方向演替。
猜你喜欢
疯狂英语·新读写(2023年4期)2023-05-10
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
活力(2019年21期)2019-04-01
少儿科学周刊·儿童版(2017年5期)2017-06-29
学苑创造·A版(2017年3期)2017-04-27
水生生物学报(2015年1期)2015-02-28
学苑创造·A版(2014年6期)2014-08-04
