
不同秋眠级紫花苜蓿叶片响应渍水胁迫的代谢组学分析
2023-12-04 06:22:02宋佳阳胡龙兴
草地学报 2023年11期
宋佳阳, 胡 瑜, 徐 倩, 卢 蕊, 胡龙兴
(湖南农业大学农学院草业科学系, 湖南 长沙 410128)
紫花苜蓿(MedicagosativaL.)具有产量高、营养价值高、适应能力强等特点,被誉为“牧草皇后”,是世界上种植广泛且十分重要的豆科牧草,也是我国实施农业结构调整、发展可持续农业的首选饲料作物之一[1-2],具有巨大的经济、环境和社会价值[3-4]。我国南方地区因降雨量大、季节分配不均、地下水位高等易形成内涝或渍水胁迫现象,进而影响紫花苜蓿的种植面积以及产量[5]。因此,厘清苜蓿对耐渍性胁迫的生理代谢响应机制对选育耐渍性强的品种具有重要的指导意义[6-7]。
秋季休眠(Fall dormancy)被定义为苜蓿在秋季响应日照减少和温度下降的适应性生长特征[8],被广泛用作抗逆性、生产性能和春季生长的重要指标[9]。张卫红等[10]对4个不同秋眠等级紫花苜蓿苗期生物学特性探究发现,秋眠等级高的紫花苜蓿品种的耐淹性高于秋眠等级低的紫花苜蓿品种;安渊等[11]]对国外引进的28个半秋眠和非秋眠品种的耐涝性分析发现,秋眠级数越高,耐涝性越强,但目前苜蓿秋眠级影响其耐渍性的作用机理尚不明确。
渍水胁迫对植物具有多方面的影响,如叶片的光合速率下降、根系呼吸受抑制、植物分生组织的伸长等,其中渍水胁迫对植物造成损伤的主要原因是根系呼吸受到抑制导致的长期缺氧。在缺氧条件下,植物根系只能进行无氧呼吸。O2是线粒体电子传递链末端的电子受体,渍水胁迫时O2可用性的降低干扰了电子传递链,迅速抑制三磷酸腺苷(ATP)的产生,从而抑制线粒体呼吸[12-13]。渍水胁迫使植物只能通过厌氧代谢途径来提供维持植物在低氧环境下各项生理活动所需的能量,而糖酵解的终产物丙酮酸被转化成乳酸和乙醇,无法通过三羧酸循环转化成植物所需能量,有机物损耗大,能量生成少[14]。为应对渍水胁迫造成的能量短缺[15],植物体内细胞首先通过糖酵解和乙醇发酵途径产生能量,同时也会伴随乳酸、醇、醛类等有毒物质的积累[16]。在高等植物中,抗涝性植物除了通过糖酵解和乙醇发酵途径产生能量外,还可以通过苹果酸代谢、琥珀酸合成、谷氨酸转化、磷酸戊糖等多条代谢途径共同保证稳定的能量及物质供应,避免单一代谢产物及有害物质的过度积累[17]。长时间在渍水胁迫下的植物会激活相关基因(如ENO1,ADH1等)进入丙氨酸发酵途径,为植物生长发育提供能量[18]。因此,紫花苜蓿的代谢途径可能与其抵抗涝渍胁迫密切相关。
因此,本研究以不同秋眠级紫花苜蓿叶片为试验材料,利用非靶向代谢组学方法,采用液相色谱-质谱(Liquid chromatography-mass spectrometry,LC-MS)技术测定渍水胁迫下紫花苜蓿叶中相关代谢物的差异,探究渍水胁迫对不同秋眠级紫花苜蓿的影响,以期为紫花苜蓿耐渍品种的选育和栽培提供理论依据。
1 材料与方法
1.1 植物材料与培养
试验于2023年5—7月在湖南农业大学草业科学系步入式人工气候室内进行。将细沙与营养土按体积比为2∶1的比例混匀,然后将等量的基质装入塑料方盆(上口径10 cm×10 cm,下口径8 cm×8 cm,高度8 cm)中。选用饱满、大小一致的种子均匀播种于基质中,每盆播种30粒,覆盖约1 cm基质,在人工气候室内培养。待种子发芽后幼苗长至2片真叶时进行间苗,每盆保留生长一致且健壮的幼苗6~8株,每隔5 d每盆浇50 mL的1/2Hoagland营养液。待植株长到15 cm时进行渍水处理,每个处理组设置4个生物学重复(4盆)。人工气候室生长条件为:温度25℃/20℃(昼/夜),光照时间14 h,相对湿度70%~75%,光照强度为450 μmol·m-2·s-1。
1.2 试验设计与处理
试验采用完全随机设计,将处理组盆栽材料放置于装满自来水的方形盆中,使水面没过培养基质表面约1 cm左右,每天补充因蒸发散失的水量。每隔2 d换一次水,分别在渍水处理第7 d和14 d取植株的倒数第三、四片复叶冻于液氮,存储于-80℃冰箱用于后续指标的测定。每个处理设置4个重复,以正常浇水对照。
1.3 测定项目与方法
1.3.1丙二醛含量测定 采用TCA-TBA法测定[19]。
1.3.2代谢产物提取与测定 称取25 mg冻干样品,加入500 μL提取液(甲醇∶水=3∶1 (V/V),含同位素标记内标混合物);35 Hz研磨处理4 min,超声5 min (冰水浴);重复步骤2~3次;—40℃静置1 h;将样品4℃,12 000 r·min-1[离心力13 800(×g),半径8.6 cm],离心15 min;取上清于进样瓶中,上机检测;所有样品另取等量上清混合成质量控制(QC)样品上机检测。
1.4 数据处理与统计分析
试验数据采用Microsoft Excel 2016整理;用SPSS 17.0软件进行方差分析和Ducan式多重检验(P<0.05),使用SigmaPlot 12.5软件进行作图。代谢组数据分析中,采用正交偏最小二乘判别分析(Orthogonal projections to latent structures-discriminant analysis,OPLS-DA)模型,以第一主成分的变量投影重要度(Variable importance in the projection,VIP)大于1与P值(P-value)小于0.05为阈值,筛选丰度上调或下调的代谢物。本研究基于京都基因和基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)Pathway数据库,使用SIMCA软件(V16.0.2,Sartorius Stedim Data Analytics AB,Umea,Sweden)对不同样本的所有代谢物进行主成分分析(Principal component analysis,PCA)并注释差异代谢物富集的代谢途径[20]。
2 结果与分析
2.1 不同秋眠级紫花苜蓿渍水表型分析
如图1所示,正常生长条件下,‘SK 3010’和‘WL 525HQ’生长状态均良好无差异;渍水处理7 d时,二者均出现涝害症状,但‘WL 525HQ’的黄叶率比‘SK 3010’低。渍水处理14 d时,‘SK 3010’叶片萎蔫严重,而‘WL 525HQ’植株叶片萎蔫较少,植株状态相对较好,叶片黄叶率低于‘SK 3010’,表明秋眠级高的‘WL 525HQ’品种比秋眠级低的‘SK 3010’品种耐渍性更强。
2.2 渍水胁迫下不同秋眠级紫花苜蓿生理指标分析
为了进一步探究不同秋眠级紫花苜蓿耐渍性,对‘SK 3010’和‘WL 525HQ’渍水处理后的叶片组织进行生理指标测定,发现紫花苜蓿‘SK 3010’和‘WL 525HQ’的MDA含量在渍水处理7 d和14 d后均高于对照组(图2),且‘SK 3010’在渍水处理7 d和14 d积累的MDA含量比‘WL 525HQ’高,表明‘SK 3010’的细胞膜创伤程度更高。
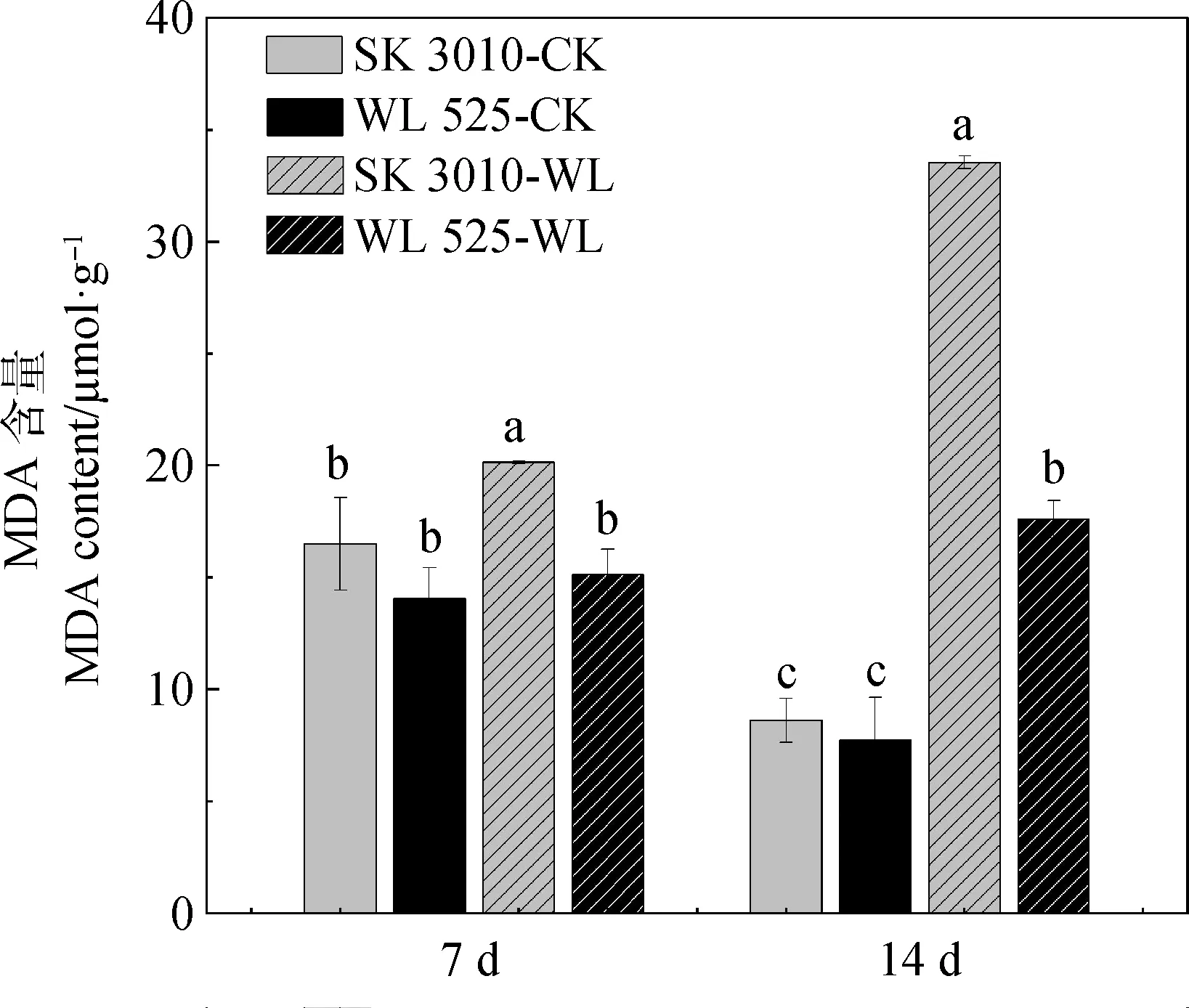
图2 渍水处理对不同秋眠级紫花苜蓿MDA含量的影响Fig.2 Effect of waterlogging on MDA content of alfalfa with different fall dormancy levels
2.3 不同秋眠级紫花苜蓿叶片响应渍水胁迫的代谢组学分析
2.3.1非靶向代谢组学数据分析 为分析渍水条件下不同秋眠级紫花苜蓿叶片代谢产物含量及代谢途径的变化,采用LC-MS技术对渍水处理前后紫花苜蓿‘SK 3010’和‘WL 525HQ’的叶片代谢物进行检测。主成分分析(PCA)结果表明四组样本的重复性良好,PC2可将两个品种的对照与渍水处理分开(图3A)。通过MS进行定性和定量分析,共检测到1 077个代谢产物,分为13组,其中,脂质类化合物345个(32.07%),有机杂环化合物147个(13.68 %),苯丙素类和聚酮类145个(13.45 %),有机酸及其衍生物140个(12.99%),苯环型化合物115种(10.69%),有机氧化合物93个(8.62%),21种核苷酸类和有机氮化合物(1.95%),碳氢化合物11个(1.03%),5个生物碱及其衍生物和木脂素类化合物(0.46%),3个有机硫化合物(0.23%),以及26种其他代谢物(2.41%)(图3B)。对渍水前后紫花苜蓿‘SK 3010’和‘WL 525HQ’叶片中差异代谢产物进行分析发现‘SK 3010’中有139个差异代谢产物,‘WL 525HQ’中有231个差异代谢产物。随后对这些差异代谢产物进行聚类分析,如图3C、3D所示,‘SK 3010’聚类为两簇,第一簇差异代谢产物含量上调,共10个,第二簇差异代谢产物含量下调,共129个;‘WL 525HQ’聚类为两簇,第一簇差异代谢产物含量上调,第二簇差异代谢产物含量下调,其中上调差异代谢产物19个,下调差异代谢产物212个。

图3 非靶向代谢数据分析Fig.3 Analysis of untargeted metabolic data注:图A,主成分分析(PCA);图B,不同秋眠级紫花苜蓿渍水处理前后代谢产物分类;图C,‘SK 3010’差异代谢物热图;图D,‘WL 525HQ’差异代谢物热图Note:Panel A,Principal Component Analysis (PCA);Panel B,Classification of metabolites of alfalfas with different fall dormancy levels before and after waterlogging treatment;Panel C,‘SK 3010’ differential metabolite heat map;Panel D,‘WL 525HQ’ differential metabolite heat map
2.3.2差异代谢物KEGG通路富集分析 本研究利用KEGG数据库对每个组的差异代谢产物进行了通路富集分析,‘SK 3010’-WL vs. ‘SK 3010’-CK、‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK中的差异代谢产物涉及多个途径,主要途径如图4所示,在‘SK 3010’-WL vs. ‘SK 3010’-CK组中,差异代谢产物在丙酮酸代谢,糖酵解和糖异生,苯丙氨酸代谢,氰胺酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢,酪氨酸代谢,D-氨基酸代谢,生物碱合成,2-氧羧酸代谢,缬氨酸、亮氨酸和异亮氨酸的生物合成,抗坏血酸和醛酸盐代谢,氨基酸的生物合成,氨酰生物合成相关等代谢途径显著富集(P<0.05)(图4A)。
2.4 关注突出成就和社会热点,培养学生的社会责任意识 生物学知识与生产、生活和社会息息相关,如健康知识、饮食常识、环保知识、遗传病的预防、吸烟的危害、转基因食品的安全问题等;生命科学的突出成就与人类的健康生活和国家的社会经济利益密切相关,如诺贝尔生理学或医学奖对维护人类健康的意义、袁隆平杂交水稻对人类社会的贡献等。教学中,教师要鼓励学生运用所学的生物学知识去参与个人和社会事务的讨论,尝试解决生活中的生物学问题,激发学生热爱自然、热爱生命、热爱生物科学、关心和维护地球生态环境的责任意识,树立建设祖国和家乡的义务感和责任感。
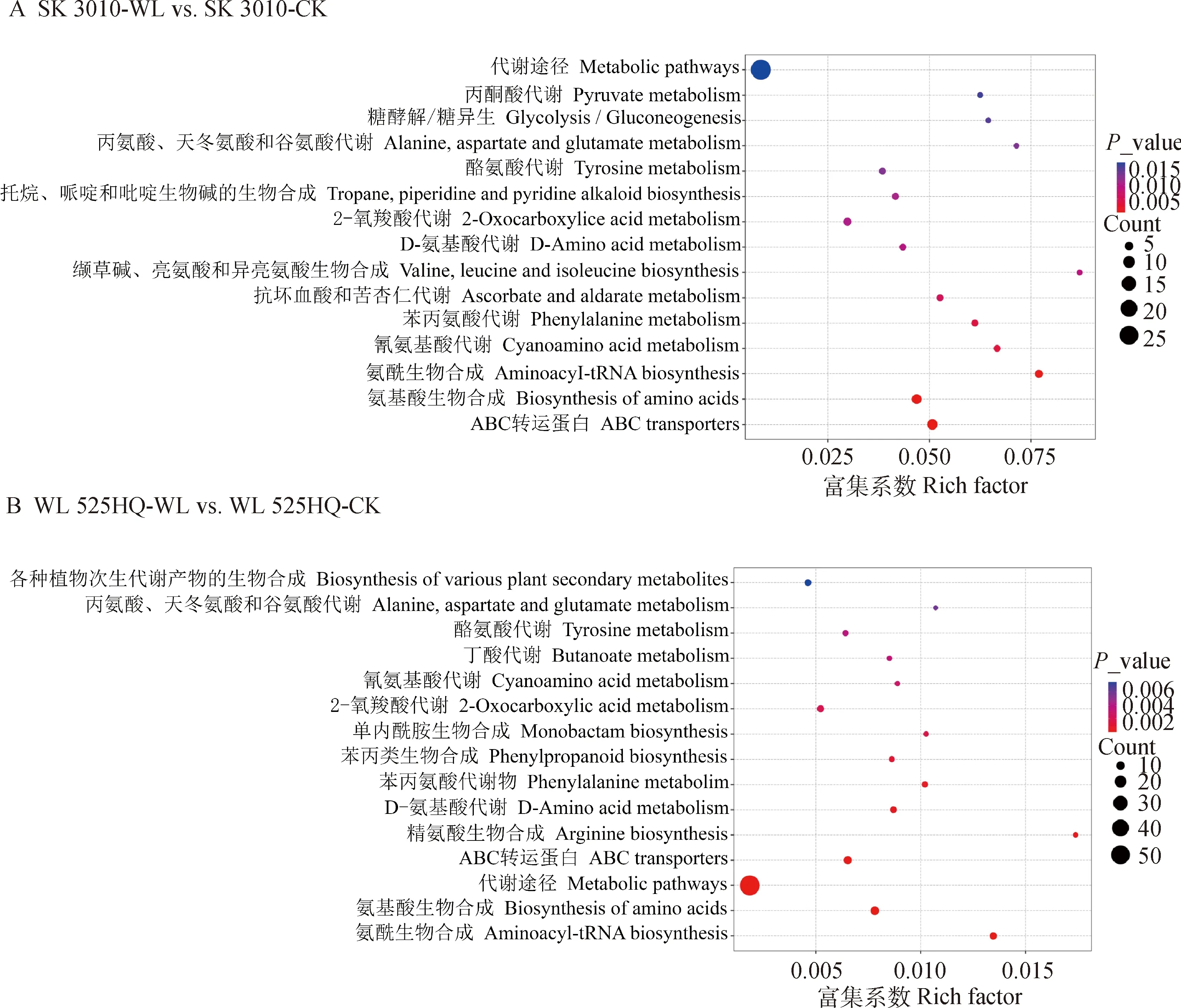
图4 成对比较差异代谢物KEGG注释和富集分析Fig.4 KEGG annotation and enrichment analysis on the pairwise comparison of differential metabolites
在‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK组中,差异代谢产物在丙氨酸、天冬氨酸和谷氨酸代谢,酪氨酸代谢,氰胺酸代谢,2-氧羧酸代谢,苯丙氨酸代谢,D-氨基酸代谢,内酰胺生物合成,苯基丙酸类生物合成,精氨酸生物合成,氨基酸的生物合成,氨酰生物合成等代谢通路显著富集(P<0.05)(图4B)。
2.3.3差异代谢物表达分析 本试验鉴定到的差异代谢物主要包括氨基酸及其衍生物、糖类、有机酸、脂类、黄酮类、核苷及其衍生物等6大类。不同类型的代谢物在‘SK 3010’-WL vs. ‘SK 3010’-CK和‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK两个比较组中的含量变化不同。如图5A所示,共有13种氨基酸在两个比较组中含量变化不同,如甲基脯氨酸、L-谷氨酸、苏氨酸等,其中甲基脯氨酸和L-苏氨酸在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中上调,在‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中下调。L-谷氨酸在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中下调,在‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中上调。共有5种脂肪酸在两个比较组中含量变化不同(图5B),D-苹果酸和柠康酸在两个比较组中受到了渍水胁迫相反的调控,二者均在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中上调,在‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中下调。如图5C所示,有4种有机酸受到了渍水胁迫的影响,在两个比较组中含量不同。其中L-乳酸和丙酮酸在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中下调幅度较大,戊二酸和羟基丙酸‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中下调更多。如图5D所示,共有11种黄酮类化合物在两个比较组中含量变化不同,如6-羟基黄酮在两个比较组中受到了渍水胁迫相反的调控,在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中上调,在‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中下调。共有5种糖类受到了渍水胁迫的影响,其中阿拉伯糖醇在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中上调幅度较大,氨基葡萄糖、D-麦芽糖、葡糖酸内酯、熊果苷在‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中下调更多。
2.3.4差异代谢物代谢通路分析 为解渍水胁迫对不同秋眠级紫花苜蓿叶片代谢途径的影响,根据‘SK 3010’-WL vs. ‘SK 3010’-CK和‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK两个比较组中的差异代谢物,绘制了其所在的代谢途径,并利用方格中不同颜色表征各差异代谢物的变化情况(图6),表明渍水胁迫显著影响TCA循环、氨基酸代谢和黄酮生物合成相关代谢途径[20-21]。
3 讨论
当植株长期处于渍水胁迫时,根系主要进行无氧呼吸,叶面气孔应激性关闭,从而导致体内ROS积累过量,细胞内产生和清除氧自由基的动态失衡,对细胞产生氧化损伤,进而导致植株地上叶片发黄凋落和地下主根腐烂[22-24]。MDA是细胞毒性物质,它与细胞膜上的蛋白质、酶等结合、交联并使之失活,最终对植物细胞产生毒害作用。因此,植物受渍水胁迫的程度可以间接通过MDA含量的高低反映[25-26]。本研究发现渍水处理后‘SK 3010’中MDA含量显著高于‘WL 525HQ’,说明‘SK 3010’受损伤程度更高,这与其地上部分黄叶率高于‘WL 525HQ’的表型相对应,这些结果表明‘WL 525HQ’的耐渍性强于‘SK 3010’,这与张卫红等[10]报道的秋眠级高的紫花苜蓿耐渍性强的研究结果一致。
代谢组学是在转录组和基因组学之后又一门新兴学科。植物感受到胁迫时,会在生理及代谢水平及时进行调整确保生命活动正常运行[27]。紫花苜蓿的耐渍调节是一个复杂的代谢网络,涉及能量代谢、氨基酸代谢、脂质代谢、黄酮类代谢、信号转导等各种通路[28]。本研究结果表明,氨基酸代谢中苏氨酸、脯氨酸、苯丙氨酸、丝氨酸、天冬酰胺、谷氨酸和谷氨酰胺等代谢途径参与了耐渍调节,黄酮和黄酮醇的生物合成,丙酮酸代谢,生物碱合成等代谢途径也参与其中。氨基酸是多种生物功能大分子蛋白的前体,在多种非生物胁迫下具有多种功能,可以通过参与相关基因表达和关键酶的活性调节等途径改变植物生理代谢进而提高植物对逆境的适应性[29]。同时氨基酸和其他含氮化合物的积累是迄今报道的几乎所有植物胁迫反应的一个显著生化特征。氮代谢的这种变化被解释为相容溶质的积累,为未来需求产生碳和氮储备。因此,含氮化合物的积累可能是对胁迫下生长速率降低导致的氮需求减少的一种反应[30]。天冬酰胺在植株体内氮的长距离运输中起着核心作用,直接或间接参与氮同化、脯氨酸和多胺的生物合成以及光呼吸对铵的解毒[31],因此,天冬酰胺代谢可能通过支持核心氮代谢在耐渍性中发挥关键作用[32]。谷氨酸和谷氨酰胺在通过谷氨酰胺合成酶/谷氨酸合成酶途径的NH4+同化过程中起着关键作用[30],谷氨酸在白菜的耐旱性和长叶莴笋的抗水分胁迫方面产生积极影响[33-34],同时谷氨酸也是其他氨基酸的氨基主要供体,谷氨酸合酶可催化谷氨酰胺和α-酮戊二酸形成谷氨酸[35]。本试验结果表明,谷氨酸、谷氨酰胺和天冬酰胺在耐渍性强的‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中含量上调,其中谷氨酰胺和天冬酰胺较耐渍性弱的‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中含量上调差异显著,因此,猜测此三类氨基酸可能与紫花苜蓿耐渍性相关。丝氨酸作为被鉴定出的防御蛋白[36],是保护膜免受活性氧损伤的内源性植物抗氧化剂之一[37]。有研究发现在镉处理的拟南芥植株中,丝氨酸的代谢水平较处理前升高[33],说明丝氨酸代谢与植物逆境胁迫相关,这与Kavi等[38]研究结果一致。本试验中,耐渍性强的‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK比较组中丝氨酸含量上调,耐渍性弱的‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中丝氨酸含量下调,说明丝氨酸对‘WL 525HQ’应对渍水胁迫具有积极重要作用,这与Wang等[36]研究结果一致。黄酮类物质具有显著的抗氧化性,可以有效地抑制或消除自由基对植物的损害,是一类重要的次生代谢产物,可以通过调控相关蛋白和基因的表达参与植物的抗逆过程[39]。在本试验中,‘SK 3010’-WL vs. ‘SK 3010’-CK和‘WL 525HQ’-WL vs. ‘WL 525HQ’-CK两个比较组中差异代谢产物含有多个黄酮类化合物,其中6-羟基黄酮在‘SK 3010’-WL vs. ‘SK 3010’-CK比较组中含量增多,说明渍水胁迫后紫花苜蓿‘SK 3010’体内产生的活性氧自由基更多,引起了具有抗氧化作用的6-羟基黄酮的上调,这与渍水胁迫下‘SK 3010’受到的损伤更严重结果相一致。
4 结论
本研究通过对不同秋眠级紫花苜蓿渍水处理发现,高秋眠级品种‘WL 525HQ’较低秋眠级品种‘SK 3010’的MDA含量更低、耐渍性更强。代谢组学分析结果表明,‘WL 525HQ’受到渍水胁迫影响的代谢途径中谷氨酸、丝氨酸、谷氨酰胺和天冬酰胺等代谢产物含量显著上调,说明这些代谢产物参与的氨基酸代谢过程提高了紫花苜蓿的耐渍性。本研究不仅为不同秋眠级紫花苜蓿耐渍性差异机制研究提供了理论参考,还为开发利用和挖掘不同耐渍性紫花苜蓿有效资源提供了重要依据。
猜你喜欢
现代临床医学(2022年4期)2022-09-29 07:36:10
中国麻业科学(2021年5期)2021-12-02 02:08:42
麦类作物学报(2021年4期)2021-05-25 06:37:30
草业科学(2021年4期)2021-05-12 14:09:06
三门峡职业技术学院学报(2017年1期)2017-06-05 10:17:30
中国洗涤用品工业(2017年2期)2017-04-16 05:07:45
中国比较医学杂志(2017年5期)2017-01-17 06:17:05
中国环境监察(2016年7期)2016-10-23 05:36:30
分析测试学报(2015年7期)2016-01-13 06:19:16
医学研究杂志(2015年12期)2015-06-10 06:57:46
