
日本结缕草(Zoysia japonica)酵母杂交cDNA文库的构建及分析
2023-12-04 06:21李艳茹周丽莹刘亚玲李舒文王梦迪晁跃辉
草地学报 2023年11期
李艳茹, 周丽莹, 董 笛, 刘亚玲, 李舒文, 王梦迪, 晁跃辉*
(1.北京林业大学草业与草原学院, 北京 100083; 2. 北京理工大学生命学院, 北京 100081; 3. 内蒙古草业技术创新中心有限公司, 内蒙古 呼和浩特 010010)
近年来随着功能基因组学研究的深入推进,分子生物学领域蛋白质相互作用的相关研究成为一个备受关注热点问题[1]。研究蛋白质之间的相互作用是探究基因功能的重要方面,通过该研究可以深入分析基因编码产物的作用机制和调控网络。酵母双杂交技术是一种分子生物学技术,主要应用于蛋白质相互作用研究[2]。该技术基于酵母菌细胞中的转录因子的结构和功能,通过激活报告基因的表达来进行目的蛋白的筛选[3]。同时,酵母杂交技术也可以在一定程度上反映细胞内部的真实情况[4]。构建高容量的cDNA酵母文库是利用酵母双杂交技术筛选目的基因以及研究蛋白互作关系的基础,也是揭示蛋白功能及其作用基础的前提,所以构建高质量cDNA酵母文库具有重要意义[5-6]。近年来,酵母杂交技术已广泛应用于蛋白质互作筛选,并在新基因挖掘及基因调控相关分子机制探究方面发挥了重要作用[7]。
日本结缕草(Zoysiajaponica)适应性广,抗逆性强,地下根系发达,密度高,覆盖面广,是一种优良的禾本科草坪草品种[8-9]。日本结缕草具有极强的逆境适应能力,不仅适应潮湿的沿海地区,还能适应内陆的干旱极端气候[10]。这些优良特性使得日本结缕草在城市绿化、运动场建设以及水土保持等方面有重要的应用价值[11-12]。高容量的日本结缕草酵母杂交cDNA文库的构建,为更深层次的探索日本结缕草相关基因提供便利。本研究挑选了来源于不同组织的日本结缕草,并进行不同的胁迫处理或激素诱导,再经过SMART技术进行了高容量的日本结缕草酵母杂交cDNA文库的构建,该文库能够用作筛选试验,可作为基础进一步进行蛋白与DNA互作或蛋白质互作等基础生物学研究。
1 材料与方法
1.1 试验材料
本实验所使用的酵母文库构建试剂盒购自Invitrogen(美国),包括CloneMinerTMII cDNA文库构建试剂盒、FastTrack MAG mRNA分离试剂盒、PureLinktm快速凝胶提取和PCR纯化组合试剂,以及质粒DNA纯化试剂盒。此外,于日本TaKaRa公司购买了MatchmakerTM酵母筛库试剂盒、实验菌株大肠杆菌DH10B菌株和Y187菌株,以及DNA连接酶、DNA聚合酶和T4 DNA连接酶。实验药物乙醇、琼脂、异戊醇、LB培养基购买自美国Sigma公司。
1.2 日本结缕草总RNA的提取及mRNA分离纯化
选取结缕草种子、生长30天的日本结缕草的根茎叶植物组织进行取样。首先,取0.1 g样品,将取出的样品置于液氮中并进行充分研磨直至样品变成粉末状。用Trizol法进行总RNA的提取,之后对提取的mRNA进行分离和纯化。最后,采用1%琼脂糖凝胶电泳检测RNA的完整性。
1.3 cDNA初级文库的构建及检测
将质量合格的日本结缕草总RNA作为模板,进行第一链cDNA的反转录合成。在E.coliDNA Ligase (10 U·μL-1)、E.coliDNA PolymeraseⅠ(10 U·μL-1)、E.coliRNaseH (2 U·μL-1)和T4 DNA Polymerase的催化下,利用合成的第一链cDNA作为模板合成第二链cDNA。将三框attB1重组接头连接至获得的第二链cDNA,随后进行cDNA分级分离并收集。对收集到的cDNA进行BP重组反应,并将反应产物转化到大肠杆菌DH10B细胞中。转化阶段完成后,将细胞转移到SOC培养基中,在37℃、摇床速度为225~250 r·min-1的条件下培养60分钟左右。随后,将菌液原液分为两部分,一部分原液取出进行后续的测定库容、重组率和插入片段长度。剩余原液则加入甘油至终浓度为20%,存储在-80℃的环境中进行长期保存。
1.4 酵母杂交cDNA文库的构建及保存
首先,需要进行初级文库质粒的提取,然后将提取好的质粒稀释到300 ng·μL-1。之后,进行LR重组反应,将质粒与载体pGADT7-DEST相结合。然后进行转化大肠杆菌DH10B感受态细胞的相关实验。最后细胞培育需要在SOC培养基上进行,用以获得酵母双杂交cDNA文库。随后,文库质量鉴定需要取出10 μL转化后的细菌原液进行相关实验。将甘油加入到剩余的菌液中直至浓度达到20%,并且需要将菌液在-80℃长期存放,以备后续使用。
1.5 Y187酵母感受态细胞的制备
为了获得Y187酵母感受态细胞,需要取少量菌株,在YPDA培养基上划线。随后,将其倒放在30℃培养箱中培养3~5天。接着,在YPDA培养基振荡培养多次挑选好的酵母单菌落,直至菌液OD600达到0.4~0.5。用单独的离心管分装最终的菌液,然后将分装好的菌液进行离心并把上清液弃掉,之后需要用无菌去离子水将沉淀进行重悬,再次离心后重悬沉淀需要加入TE/LiAc溶液。最后使用TE/LiAc溶液悬浮细胞,可以得到Y187酵母感受态细胞的悬浮液。
1.6 cDNA文库转化Y187酵母感受态细胞
首先,将cDNA文库质粒取出5 μL,并将其转化到制备好的Y187酵母感受态细胞中。将转化后的细胞原液取出10 μL,分别稀释到原来的10,100,1 000,10 000倍,然后将稀释后的液体涂布于SD/-Leu平板上,之后在30℃培养箱中将SD/-Leu平板倒放并将其培养3~5天。最后进行转化子的收集,并计算细胞密度和进行文库的滴定数的确定。在SD/-Leu平板上随机挑选24个克隆,用来鉴定菌落PCR。挑选单菌落进行PCR检测以及琼脂糖凝胶电泳检测,并进行文库重组率的相关计算。
1.7 转录自激活验证
将转化好的pGBKT7-ZjCCD7和pGBKT7载体的酵母菌液点涂在SD/-Trp,SD/-Trp/X-α-Gal和SD/-Trp/X-α-Gal/AbA培养基上,放入30℃培养箱中进行2~3天的培养,之后对pGBKT7和pGBKT7-ZjCCD7出现的菌落状态进行观察验证,看其是否具有转录自激活活性。
1.8 ZjCCD7互作蛋白的筛选及验证
按照MatchmakerTM酵母筛库试剂盒的要求进行ZjCCD7蛋白的筛库。将pGBKT7-ZjCCD7和pGADT7-atpG一起转化至酵母感受态细胞中作为实验组,并设置pGBKT7-53+pGADT7-T作为阳性对照,pGBKT7-53+pGADT7-atpG,pGADT7-T+pGBKT7-ZjCCD7共同作为阴性对照,将摇好的酵母菌液在SD/-Leu-Trp-His-Ade/X-α-Gal固体培养基上点涂,并放入30℃培养箱下进行2~3 d的培养,之后对菌落状态进行观察记录。
2 结果与分析
2.1 日本结缕草mRNA分离质量良好
用1%琼脂糖凝胶电泳进行检测(图1),检测结果显示,纯化后的mRNA条带呈现为一条模糊的拖带,大于500 bp的部分是该拖带最亮的部分。该结果表明,分离纯化后的mRNA没有发生分解,且质量良好,能够用作建库起始样品。

图1 mRNA分离结果Fig.1 mRNA separation results
2.2 初步构建cDNA初级文库
将纯化后的ds-cDNA取出5 μL,用以1%琼脂糖凝胶电泳检测。检测结果显示,ds-cDNA的条带呈现为一条弥散状,该条带大小在250~2 000 bp之间(图2)。表明纯化后的cDNA满足下一步建库试验的要求,可以用于进行后续实验。
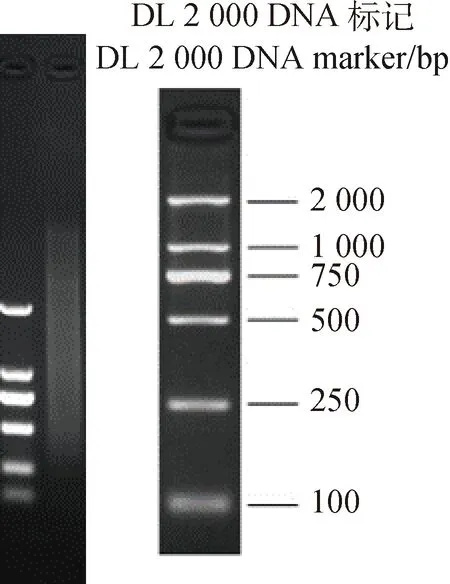
图2 二链合成结果Fig.2 Two-strand synthesis results
2.2.1初级文库重组率鉴定 从该克隆文库中随机挑选24个克隆,并将挑选好的克隆进行菌落PCR的鉴定。经过1%琼脂糖凝胶电泳检测,并对其重组率和插入片段长度进行了分析(图3)。结果表明该初级文库拥有100%的重组率,并且其平均插入片段长度在1 000 bp之上,因此该文库符合高质量初级文库的要求。

图3 重组率鉴定Fig.3 Identification on the reorganization rates of primary library注:初级文库的24 个克隆的菌落PCR鉴定Note:PCR identification on the colonies of 24 clones of primary libraries
2.2.2初级文库库容量鉴定 取10 μL转化后的细菌原液,并对其进行稀释至原来的100倍,之后从稀释好的液体中取50 μL,并将其均匀涂布在含有卡那霉素的LB培养基上。根据公式计算得到,初级文库总克隆数为cfu=2.6×106×4=1.04×107(图4)。
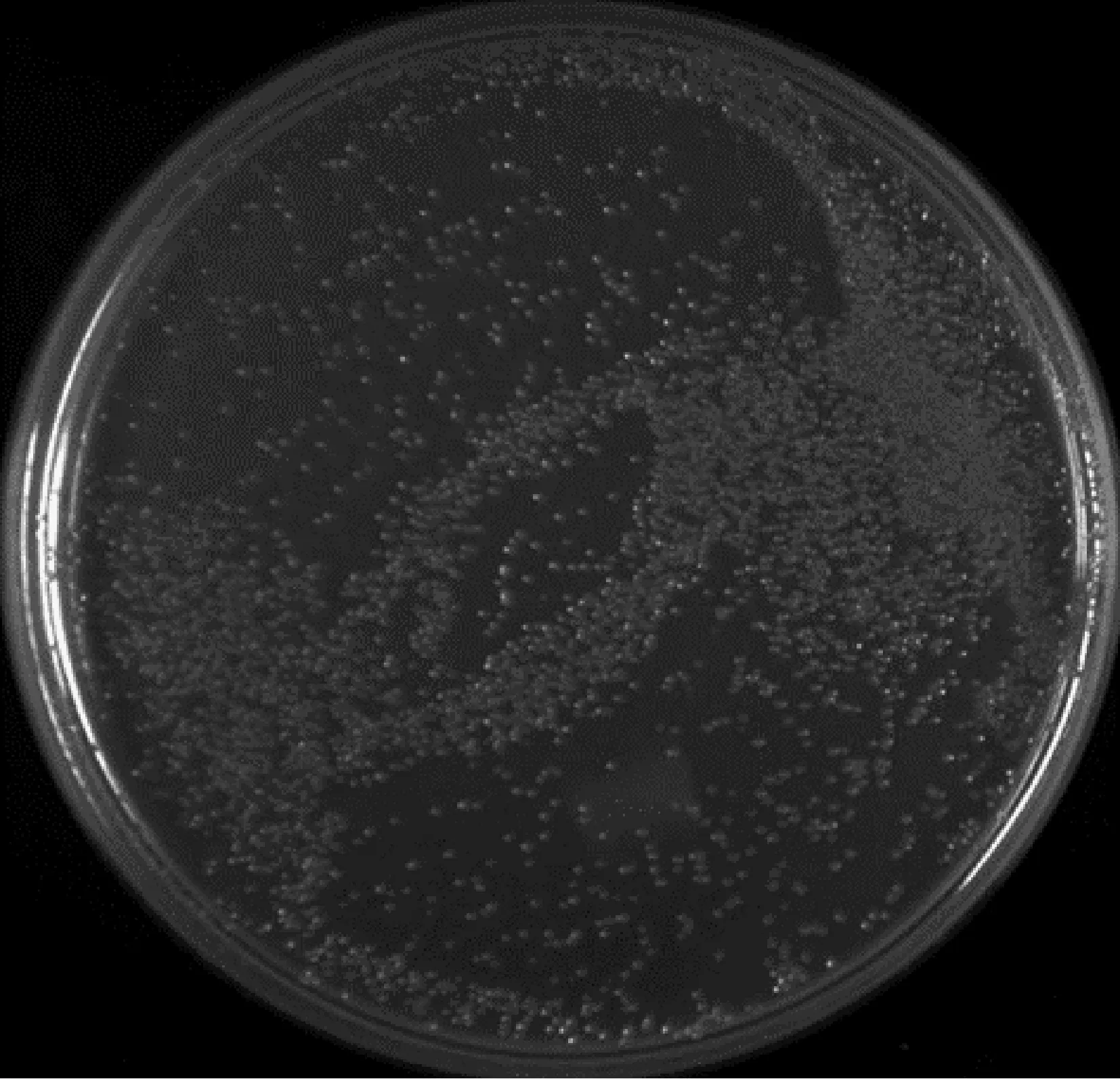
图4 初级文库库容量鉴定Fig.4 Capacity identification of the primary library
2.3 次级文库构建
2.3.1次级文库插入片段重组率鉴定 从文库中随机挑选了24个克隆,并对选取的24个克隆进行菌落PCR鉴定,还进行了重组率和插入片段长度的分析。该凝胶电泳结果显示,随机挑选好的24个克隆拥有100%重组率,且平均插入片段长度在1 000 bp以上(图5)。结果表明满足构建该酵母杂交cDNA文库的高质量要求条件,后续的日本结缕草酵母杂交相关试验可以使用该酵母杂交cDNA文库。

图5 次级文库重组率鉴定Fig.5 Identification on reorganization rates of the secondary library注:次级文库的24 个克隆的菌落PCR鉴定Note:PCR identification on the colonies of 24 clones of the secondary library
2.3.2次级文库库容量鉴定 将转化后的细菌原液取出10 μL,并将其稀释至原来溶液的100倍,之后在LR培养基上均匀涂布取出的50 μL稀释好的溶液,将该培养皿倒放在恒温培养箱中,并在第二天对其进行计数(图6),根据公式进行计算,得到次级文库总克隆数为cfu=4.0×106×4=1.6×107(图6)。
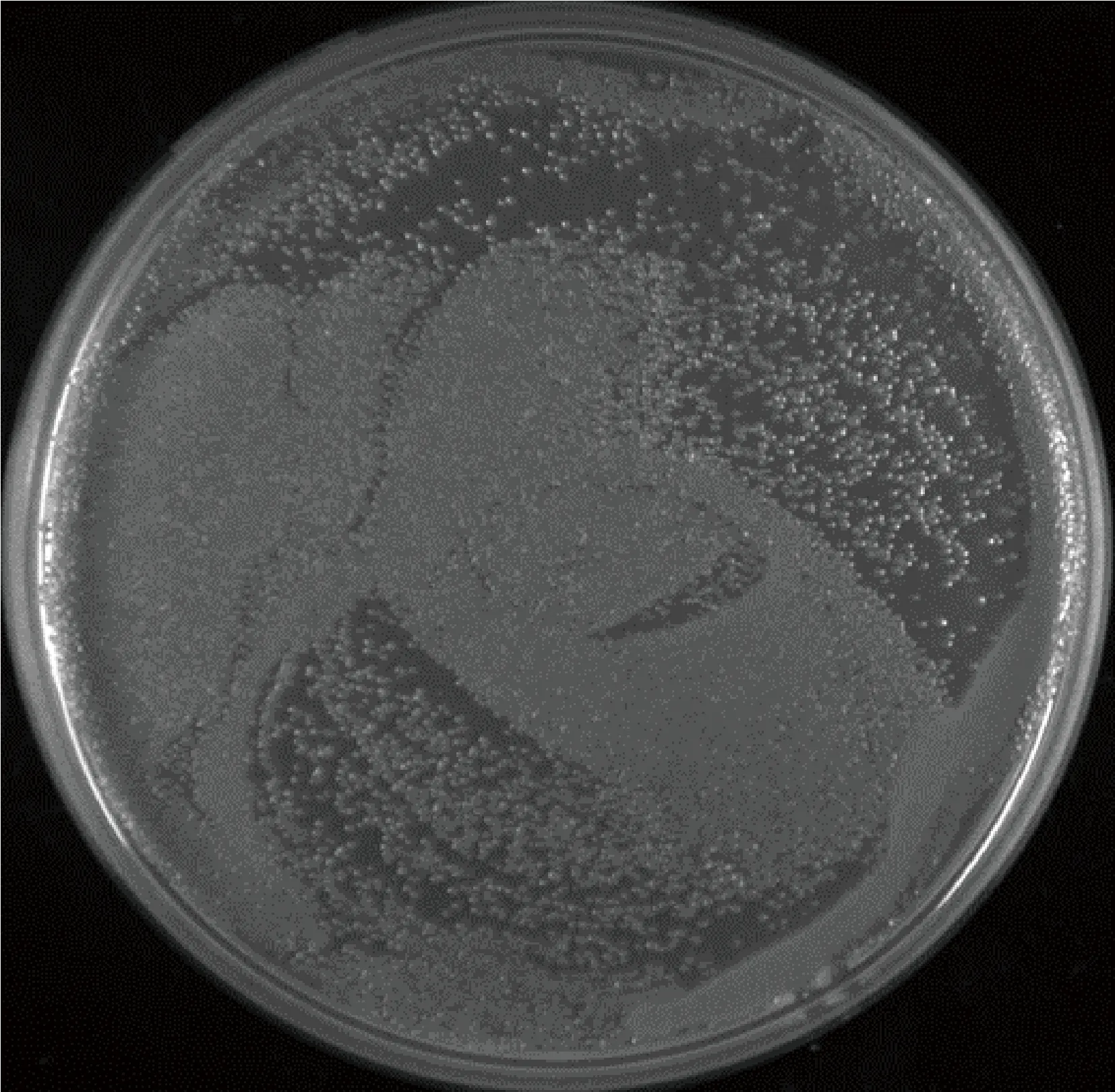
图6 次级文库库容量鉴定Fig.6 Capacity identification of the secondary library
2.4 cDNA文库转化Y187 酵母感受态细胞
取出100 μL液体,将其在稀释度为1∶10 000的SD/-Leu培养基上均匀涂布。该培养基克隆数为300,且细胞密度大于3×107cells·mL-1(图7)。
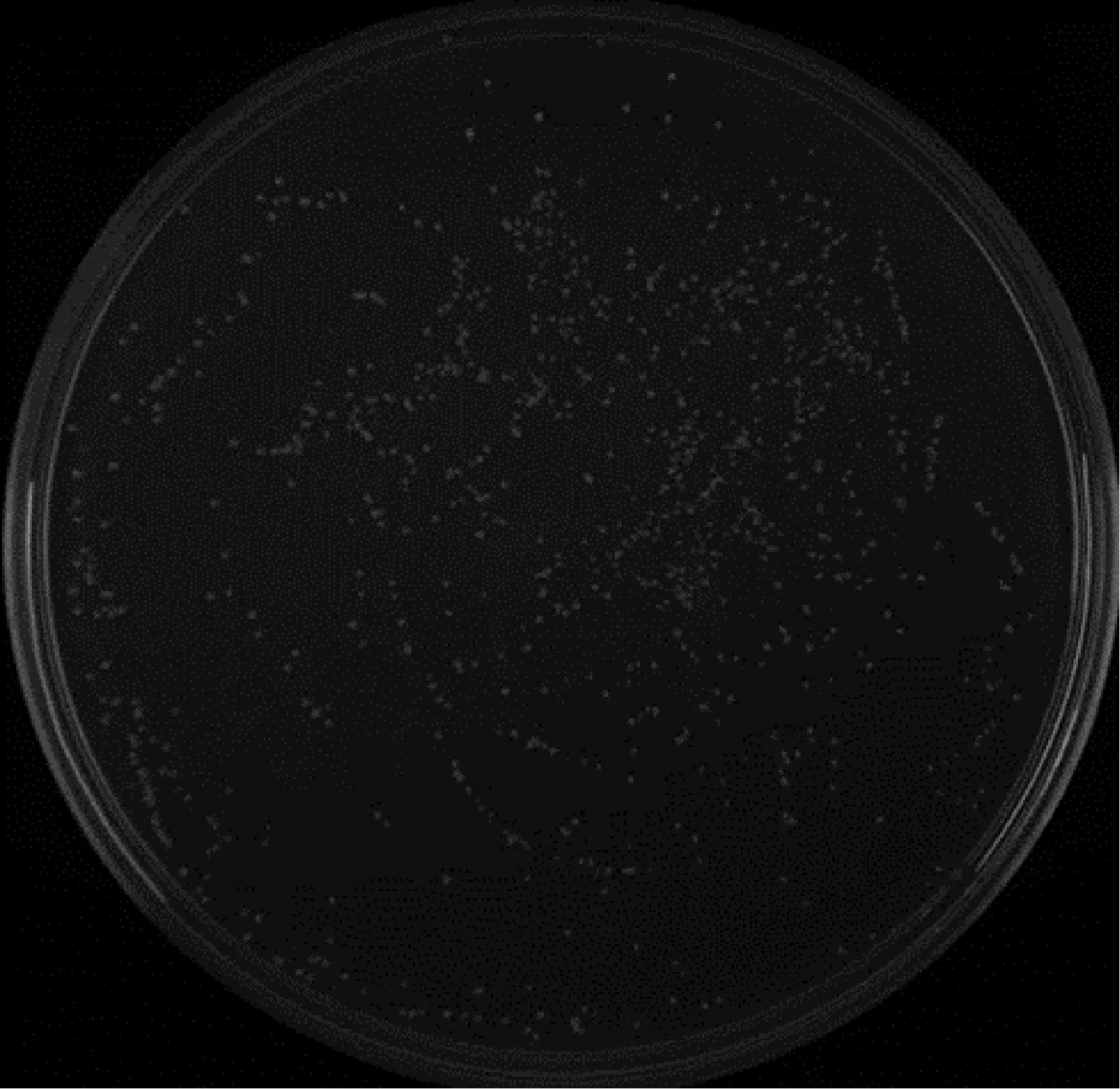
图7 文库滴度鉴定Fig.7 Titer identification on the library
随机挑选该SD/-Leu培养基上的24个单菌落,并将这些单菌落进行扩大培养,之后完成PCR检测(图8)。结果显示其中可扩增出条带的单克隆有23个,平均插入片段长度大于1 000 bp,根据公式可得文库重组率>95%。
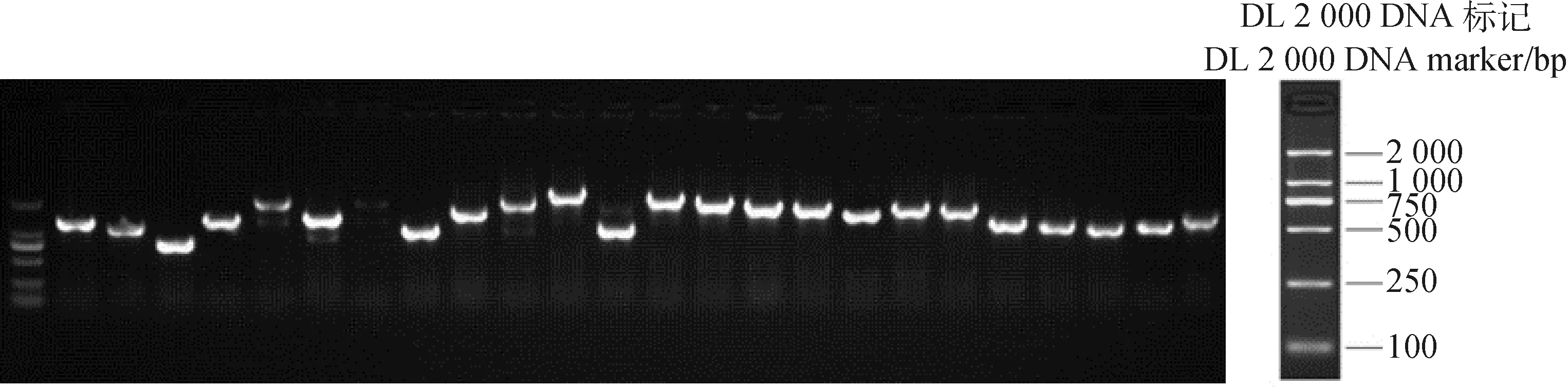
图8 酵母克隆鉴定Fig.8 Identification on the yeast clones注:酵母克隆的24个克隆的菌落PCR鉴定Note:PCR identification on the colonies of 24 clones of yeast clones
2.5 ZjCCD7自激活检测与蛋白互作
2.5.1ZjCCD7自激活检测 为了对构建的日本结缕草酵母文库进行检测,选择类胡萝卜素裂解双加氧酶7进行验证。将转化成功的pGBKT7-ZjCCD7诱饵表达载体的酵母菌株,在SD/-Trp,SD/-Trp/X-α-Gal,SD/-Trp/X-α-Gal/AbA这3种缺陷固体培养上点涂并观察生长情况。实验结果显示,有酵母菌落在SD/-Trp培养基上并且正常生长。而且转入的pGBKT7-ZjCCD7酵母菌株在SD/-Trp/X-α-Gal培养基上酵母菌落没有显示蓝色且正常生长,在SD/-Trp/X-α-Gal/AbA培养基上没有酵母菌落生长且无颜色变化(图9)。因此构建成功的pGBKT7-ZjCCD7诱饵表达载体没有自激活活性,可应用于酵母双杂交系统的蛋白筛选试验。

图9 pGBKT7-ZjCCD7酵母转化转录自激活检测Fig.9 Transcriptional self-activation assay of pGBKT7-ZjCCD7 yeast transformation
2.5.2ZjCCD7互作蛋白筛选及验证 将ZjCCD7蛋白按照MatchmakerTM酵母筛库试剂盒的要求进行筛库。筛选得到了28个变蓝的克隆,通过初步筛选和生物信息学分析后,对其中一个阳性克隆进行了鉴定和酵母互作验证。将pGBKT7-53+pGADT7-T作为阳性对照,pGBKT7-53+pGADT7-atpG,pGADT7-T+pGBKT7-ZjCCD7作为阴性对照,pGBKT7-ZjCCD7+ pGADT7-atpG作为实验组。将它们转化好的酵母菌液均匀点涂在SD/-Leu-Trp-His-Ade/X-α-Gal培养基上并放入培养箱培养2~3天,之后发现阳性对照pGBKT7-53+pGADT7有菌落生长且颜色变蓝,阴性对照pGBKT7-53+pGADT7-atpG和pGADT7-T+pGBKT7-ZjCCD7未见菌落生长且没有颜色变化,作为实验组的pGBKT7- ZjCCD7+ pGADT7-atpG在SD/-Leu-Trp-His-Ade/X-α-Gal培养基上正常生长且变蓝(图10),说明ZjCCD7与ZjatpG互作能激活GAL4酵母双杂交系统的报告基因MEL1的表达。应用所构建的日本结缕草酵母文库成功筛选得到了ZjCCD7的互作蛋白,这表明文库可用于后续的日本结缕草互作蛋白筛选实验以及功能验证实验。

图10 ZjCCD7蛋白互作Fig.10 ZjCCD7 protein interactions
3 讨论与结论
生物技术领域目前的各项研究已经广泛应用了cDNA文库,所以,构建高容量的cDNA文库至关重要,有效研究的进行需要高容量的cDNA文库。文库中所含cDNA种类的完整性可用来体现文库的代表性,文库质量的高低可以用建库来源组织中所表达的遗传信息完整性来反映[13]。同时,高质量的cDNA文库也对文库插入片段的大小有一定要求,文库插入片段的长度应高于0.3 kb[14],高质量的文库插入片段应高于1 kb[15],此外文库插入片段长度应当接近于cDNA长度,过短或过长均会造成文库质量下降[16]。文库质量降低是由于平均插入片段过小,而过度筛除cDNA片段则会造成初始文库滴度过低或丢失部分基因信息[17]。本研究成功构建了高容量的日本结缕草酵母杂交cDNA文库,可用于日本结缕草中进行互作蛋白筛选或调控网络分析。该文库包含丰富的cDNA克隆和完整的遗传信息。日本结缕草为暖季型禾本科草地植物[18],其基因组大小为334 Mb,含有59 271个蛋白编码基因[19]。构建的日本结缕草初级文库总克隆数为1.04×107,为蛋白编码基因的175倍,该文库插入片段平均长度在1 000 bp以上,cDNA片段拥有>95%的重组率,满足cDNA文库的高质量、完整性要求条件,符合进行蛋白互作筛选的文库要求,后续的酵母杂交试验可以使用该cDNA文库。
酵母杂交系统主要包括酵母双杂交技术和酵母单杂交技术。酵母双杂交技术可用于互作蛋白的筛选,酵母双杂交筛选技术是确定两个蛋白质相互作用的关键技术之一,具有高度灵敏性和完善的报告系统[20],酵母双杂交技术可研究抗原与抗体的相互作用,可用作药物作用位点的筛选,在药物的发现和开发过程中起着重要的作用[21],也可用于蛋白和药物相互作用以及基因组蛋白连锁图等方面。而酵母单杂交技术则主要用于探究蛋白质与DNA的相互作用[22-23]。本研究利用酵母杂交技术,进行了日本结缕草ZjCCD7和atpG蛋白互作筛选试验。
构建植物cDNA文库已成为研究基因功能最常用的技术手段之一,cDNA文库是研究某一生物特定器官、组织和发育阶段基因表达的前提和基础,这需要在基因组水平上进行研究[24],构建成功的cDNA文库可用于研究基因的功能、筛选互作蛋白以及探究蛋白质与DNA的相互作用。因此,高质量植物cDNA文库的构建具有重要意义,该文库不仅能够深入研究植物基因和蛋白质功能,还可以用来发掘潜在蛋白功能[25]。本研究成功构建了日本结缕草cDNA文库,为了验证构建的日本结缕草酵母文库,以实验室克隆的日本结缕草类胡萝卜素裂解双加氧酶为诱饵,进行互作蛋白质的筛选。利用酵母双杂交技术筛选ZjCCD7互作蛋白,最终筛选到日本结缕草ZjatpG蛋白。已知类胡萝卜素裂解双加氧酶7(CCD7)是类胡萝卜素降解以及独角金内酯合成途径中的关键酶,在植物生长过程中起着重要作用[26],而atpG编码ATP合酶γ亚基与质子运输ATP合成酶有密切的关系[27]。本研究进行的ZjCCD7和atpG蛋白互作筛选试验,表明日本结缕草酵母文库可应用于蛋白筛选实验,互作结果显示日本结缕草类胡萝卜素裂解双加氧酶行使功能过程可能与ATP合酶相关,相关通路的调控模式仍需更深层次的探究,从而为后续ZjCCD7的调控机制的研究奠定基础。
本研究进行了高容量日本结缕草酵母杂交cDNA文库的构建,日本结缕草互作蛋白的酵母双杂交筛选和蛋白质与DNA互作的酵母单杂交试验都可以用到该文库,具有广泛的应用前景。本研究成功构建了高容量的日本结缕草cDNA酵母文库,满足高质量、完整性的cDNA文库要求,以该文库的高质量和完整性作为试验基础,可继续通过后续酵母杂交深入挖掘日本结缕草生长发育的基因功能。
猜你喜欢
湖南农业大学学报(自然科学版)(2022年2期)2022-05-11
作物学报(2021年11期)2021-08-31
猪业科学(2021年3期)2021-05-21
山东畜牧兽医(2021年6期)2021-01-11
幽默大师(2020年10期)2020-11-10
中华诗词(2019年1期)2019-11-14
猪业科学(2018年4期)2018-05-19
中国调味品(2017年2期)2017-03-20
创新作文(小学版)(2016年16期)2016-11-11
现代检验医学杂志(2016年5期)2016-08-20
