
不同土壤水分对植物光合作用的影响研究进展
2023-11-29 02:20杨佳鹤何进宇刘飞杨崔烜玮
节水灌溉 2023年11期
杨佳鹤,何进宇,2,3,刘飞杨,崔烜玮
(1.宁夏大学土木与水利工程学院,银川 750021;2.宁夏节水灌溉与水资源调控工程技术研究中心,银川 750021;3.旱区现代农业水资源高效利用教育部工程研究中心,银川 750021)
0 引 言
干旱是限制农业发展的主要因素,使得农业生产的发展受到限制和影响[1],但随着农业节水不断增强,流域农田灌溉水利用效率得到提高,农业用水公平性已由“相对公平”转为“高度公平”[2]。而土壤水分是作物正常生长发育和作物生产力的基本保证,土壤水分可以通过调节作物的光合作用来影响作物的生长和生产,继而合理的灌溉可以有效提高作物水分利用效率。1966 年澳大利亚著名水文和土壤物理学家菲利普(J.R.Philip)构建了“土壤-作物-大气连续体”系统(Soil-Plant-Atmosphere Continuum,简称SPAC[3]),它是指土壤中的水分由作物根系吸收进入体内,又由于蒸腾进入大气,随后大气水分又经降水进入到土壤中,如此形成一个动态的连续系统。其中,光合作用是SPAC 系统中蒸腾环节的重要推动因子,土壤含水量是作物生长过程中最为关键的因素[4]。近年来,有大量的学者针对作物光合作用的影响因素进行了大量研究[5-8],包括光照强度、CO₂浓度、光合速率、温度、酸碱度等内部因素,而对作物体内水分对光合作用的影响研究都是个别因素的讨论,没有较完整的水分对所有光合作用因素影响的论述。且作物的光合作用速率、胞间CO2浓度以及叶绿素总量等在不同水分胁迫以及光照条件下都会受到不同程度的影响,作物会由于土壤水分的亏缺使自身作出反应来适应干旱的环境[9],而光照除了保证作物光合作用正常进行外,还为作物的生长提供能量[10],使其对水分与光照的需求达到一定的平衡状态。因此探究土壤水分与光的平衡关系很有价值。本文以节约利用水资源和促进作物高效生长为目标,以提高作物产量和提升作物品质为目的,使农田灌水都能到达充分利用的状态。因此在前人研究的基础上提出土壤水分与光效应对作物生长的影响有潜在关联,并综述了不同土壤水分对于作物光响应的影响,更进一步探究土壤水分与光照对作物光合作用的影响,以期为作物生长提供科学的理论价值。
1 光合作用的基本原理
作物吸收光能,将水和二氧化碳生成富能有机物并释放氧气的过程叫光合作用[11],一般分为2 个过程,第1 个过程为光反应,是作物吸收阳光并在类囊体薄膜上分解水形成能量和氧气的过程;第2个过程为暗反应,是消耗光反应生产的能量及吸收的二氧化碳并生成糖的过程。光合作用对地球上几乎所有生命的存在起到支撑作用,进行光合作用的生物的生长发育与光合利用效率是有关的[12]。光饱和点、表观量子效率、暗呼吸速率、最大光合速率以及光补偿点[13]都为光合特征的有效参数。表观量子效率是反映作物在光照较弱的条件下,对光能利用、吸收以及转换的一个指标;光饱和时的最大光合速率是叶片光合作用合成能力的一个反应标量;暗呼吸速率是指在无光条件下作物的呼吸速率;光饱和点是指作物在光照充足的条件下利用光能的能力指标;光补偿点则相反[14]。
光合作用的影响因素分为环境因素和生理因素,环境因素主要包括水分胁迫、光照强度、大气CO2浓度,生理因素主要包括气孔导度、胞间CO2浓度、气孔限制值。据大量研究表明,水分胁迫是植物进行光合作用的最大限制因子[15-17],光合作用是水分影响植物生长和代谢首先表现出的重要环节[18]。因此,研究土壤水分对植物光合作用的影响,对于植物合理的浇灌水量及有效的水分吸收具有深远影响。
2 土壤水分对光合作用的影响
2.1 土壤水分对净光合速率、表观量子效率和暗呼吸效率的影响
土壤水分对于作物生长的重要性不言而喻,土壤含水量的多少会不同程度地影响作物的光合以及蒸腾作用,作物生育期内的水分轻度胁迫会提高光合速率和蒸腾速率,在适当的水分胁迫下有些作物此时的光合以及蒸腾速率会达到峰值,从而有效地促进光合作用的进行。且相关研究表明,作物经历轻旱再复水后,水分利用效率会明显提高,且能保持作物产量基本不变[19]。
陈起阳[20]选择雷公藤采用盆栽试验的方法,开展了作物在不同土壤水分条件下的光响应研究,结果表明,不同水分胁迫下随着光合有效辐射的不断增加,净光合速率也随之增加直至最大值然后开始下降,且土壤水分的减少,使光响应单峰曲线呈降低走向,表现出水分胁迫影响光能利用率。王永红等[21]选用天杨叶开展了不同土壤含水量对天杨叶生理参数的光响应试验,结果表明,当土壤质量含水量大于13.25%小于17.96%时,光合作用受到土壤含水量的影响不大,且当有效光合辐射达到1 600 µmol/(m2·s)时,光合速率变化平缓,达到最大值;当土壤质量含水量低于和高于上面范围时,会造成光合速率降低,光饱和点下降。朱永波等[22]对玉米叶片光合作用的研究中表明,在正常水分条件下,最大净光合速率较高,在轻度胁迫下,仍维持较高速率,而在重度胁迫下,光合速率则小于1 µmol/(m2·s);暗呼吸速率在不同胁迫下的表现与净光合速率相似;表观量子效率在轻度胁迫下呈下降趋势,严重胁迫下则降至较小值。云文丽等[23]探究了光合作用下向日葵的水分胁迫规律,结果显示,向日葵不同生育时期随着水分胁迫的增加,最大光合速率下降,暗呼吸速率表现出在营养生长阶段达到最大值,随着向日葵的生长逐渐下降,最低值出现在成熟期;开花期、二对真叶—花序形成期表观量子效率在土壤缺少一定水分时出现最大值,土壤过于湿润反而最低,而在开花至成熟期间却表现出相反的现象。陈志成等[24]通过珍珠油杏在不同土壤水分条件下光合作用的响应研究发现,水分胁迫的下降使得珍珠油杏叶片的净光合速率增加,而当土壤含水量一定时,净光合速率会随着光合有效辐射的增加出现先上升后趋于稳定的现象;表观量子效率在不同土壤水分条件下的值有显著的差异,表现为随着土壤含水量的降低,出现了先上升后下降的现象;珍珠油杏的暗呼吸效率则随着土壤水分的降低,呈现出持续下降的变化。李建查等[25]通过对甜玉米的研究发现,净光合速率在土壤水分为11.50%~14.50%时,光合有效辐射的增加会使其增大,直到光饱和点,而土壤水分低于11.50%时,净光合速率在所有组中是最低的,表现为强光加水分胁迫破坏了作物的光合生理特性,且由于水分胁迫的增加光饱和点以及补偿点开始下降,表观量子效率先增加后减小。
土壤含水量以及光合有效辐射都会对作物净光合速率产生影响,但土壤水分是主要的影响因素。当土壤水分过高或者过低时都会使得净光合速率下降,不同的作物,以及同一作物的不同时期,其土壤含水量的不同都会使得其表观量子效率、暗呼吸速率、光饱和点和光补偿点出现不同的变化;在一定水分下表观量子效率增大是由于对弱光的利用能力得到加强导致的,而随着水分胁迫的增加以及光照的增强,作物会因为双重胁迫而导致光系统的破坏更加严重,从而导致光合作用严重下降。综上所述,在前人研究的基础上总结出土壤水分对净光合速率、表观量子效率和暗呼吸效率的影响和不同水分下光合有效辐射对净光合速率、表观量子效率和暗呼吸效率的影响规律,如表1 和表2 所示。因此针对不同作物在不同时期进行有效的灌水,使其达到该时期最合适的生长指标是未来研究的方向之一。

表1 土壤水分对净光合速率、表观量子效率和暗呼吸效率的影响Tab.1 Effects of soil moisture on net photosynthetic rate, apparent quantum efficiency and dark respiration efficiency

表2 不同水分下光合有效辐射对净光合速率、表观量子效率和暗呼吸效率的影响Tab.2 Effects of photosynthetic effective radiation on net photosynthetic rate, apparent quantum efficiency and dark respiratory efficiency under different moisture levels
2.2 土壤水分对作物叶绿素的影响
光合色素在作物光合作用中起到能量吸收以及传递的作用,而土壤含水量的改变也会导致作物色素的变化。有研究表明,叶绿素a 和叶绿素b 在缓慢干燥的环境中会被分解或者损失,当水化时却能够被激活再次合成[26],这也说明了土壤含水程度对作物光合色素的巨大影响。王海珍等[27]对灰胡杨的光响应研究发现,叶绿素a、b 和类胡萝卜素含量以及比值都随着土壤含水量的下降而减少,且土壤水分出现中、重度胁迫时,叶绿体会被造成一定的伤害,使得光合能力降低;陈小龙等[28]通过研究不同土壤水分条件对冬烟旺的光合作用影响表明,土壤含水量的减少会造成光合色素的降低,且随着土壤水分的持续减少,色素含量越少,叶绿素a、b 的比值更小,而类胡萝卜素则受到的影响较小;刘晓静等[29]通过对银杏幼苗的光合特性研究表明,水分胁迫的增加使得光合色素先增高之后下降,叶绿素a与叶绿素b在土壤含水量为59.61%时达到最高值,2 者在土壤含水量分别为25.94%和29.67%时达到最低,而类胡萝卜素则随着土壤含水量的下降出现先下降后上升的现象,在土壤含水量为21.66%时达到最大值;周珺等[30]通过对湿地松幼苗的研究表明,总叶绿体含量在淹灌条件下小于轻度干旱条件下,且与正常灌水条件下差异显著,这表明过多的土壤含水量会导致作物体内的叶绿素含量下降;王振夏等[31]通过研究不同土壤水分交替变化条件下湿地松幼苗的光合特性表明,在水淹条件下,作物总叶绿素的含量出现了下降的现象,从前期正常供水到中期轻度干旱的土壤含水量变化过程中,作物总叶绿素含量相对于对照出现了降低,而在试验末期表现出3个阶段都处于水淹条件下的湿地松总叶绿素含量最高,而类胡萝卜素含量在前期水淹条件下表现出显著低于正常供水组,但在中后期水淹情况下与对照组无显著差异,而中期受到轻度水分胁迫组的类胡萝卜素含量则是最高,在整个周期水淹处理叶绿素a 和b 的比值与对照组没有明显差异,叶绿素与类胡萝卜素的比值则是在中期显著上升,后期与对照组无明显差异。
部分前人们的研究表明随着土壤含水量的下降,作物总叶绿素的含量也出现下降的现象,且在淹水条件下作物总叶绿素含量也表现出同样的结论,但也有研究得出淹水条件下相反的结论,造成不同结论的原因是因为淹水高度不同所导致,并且在淹水高度相同的情况下,由于光照、环境温度的不同从而叶绿素总含量的测定也会有所不同,因此在未来作物叶绿素的研究中,可进一步开展湿地作物和旱地作物分别在淹水条件或者水分胁迫条件下不同时期总叶绿素含量以及类胡萝卜素含量的变化,研究是否能在不同时期通过增加或者减少土壤含水量从而使得作物能够更有效地进行光合作用,最终达到改善品质以及增加产量的效果。
3 土壤水分对叶片蒸发速率、气孔导度和胞间CO2浓度的影响
作物所处环境的土壤水分条件对于作物的蒸腾速率、胞间CO2浓度和气孔导度都会产生不同程度的影响,气孔限制的主要原因通常为水分胁迫严重,导致气孔收缩,使得CO2进入作物受阻,破坏了植物内部的碳水循环[32],无法达到理想的光合作用效果;非气孔限制则通常指由于水分胁迫导致作物光合器官受损,进而影响光合作用。
夏江宝等[33]对紫藤光响应的研究结果表明,蒸腾作用主要受土壤水分的影响,而受光照强度变化的影响较小;当土壤湿度为18.4%~26.5%时,随着光合有效辐射强度的上升,气孔导度降低,紫藤叶片的胞间CO2浓度下降,此时气孔限制主要影响光合作用;当土壤湿度不在该范围时,光合作用由于气孔导度的持续下降,导致CO2浓度降至临界点,从而引起光合机构受损,光合能力下降。董梅等[34]对银水牛果在不同水分胁迫下的光响应试验结果表明,光合有效辐射的增大会使得银水牛果的蒸腾速率增加,土壤严重缺水时,光合有效辐射对蒸腾速率的影响不显著,而当土壤水分增多时,叶片的蒸腾速率也逐渐提升。夏宣宣等[35]对黄刺玫叶片光响应的试验表明,叶片蒸腾速率达到最大值时对应的土壤水分含量为81.2%,而当土壤中的水分含量小于36.2%时,胞间CO2浓度与气孔限制值分别上升和下降。李红等[36]设置不同的水分胁迫,研究了黄荆在此条件下的光响应影响,结果发现,当光强小于临界值时,光强度的增加会导致黄荆蒸腾速率上升,当土壤水分较充足时,蒸腾速率也较高。张仁和等[37]通过对玉米在干旱胁迫下的光合特性研究发现,在同一土壤水分胁迫下,作物的气孔导度随着光强的增加出现先上升后下降的趋势;在不同水分胁迫下,气孔导度随着水分胁迫的增加而下降;在同一光照下时,随着水分胁迫的增加,胞间CO2值出现先下降后上升的现象,而气孔限制值则出现相反的现象,此时说明光合作用受到非气孔限制。陈志成等[38]研究了不同土壤水分条件对生柿树光响应的影响,结果表明,在低光照强度下,蒸腾速率随着光合有效速率的增加出现先上升后趋于平缓的现象,随着水分降低,蒸腾速率出现先升高后下降的现象,气孔导度也表现出相同的趋势;在弱光条件下,光合有效辐射的增强会使得CO2浓度出现减少现象,之后趋于平缓;当土壤含水量在48.8%时气孔限制主要影响净光合速率,土壤含水量在25.5%时则相反。柯世省[39]通过对夏蜡梅的光合作用研究表明,土壤含水量的下降会使得蒸腾速率下降,并且气孔阻力上升,CO2浓度在水分没到达重度胁迫时,表现为下降,当处于重度胁迫时,胞间CO2浓度上升,此时表明气孔限制转化为非气孔限制。SONG[40]对玉米的光合作用特性研究发现,土壤中的水分对于光合作用由气孔限制转化为非气孔限制十分关键,且在不同时期导致气孔限制转化为非气孔限制的水分胁迫不同。
通过前人们的研究可知,作物蒸腾速率主要受到土壤水分的影响,不同的作物表现出气孔限制值转化为非气孔限制值的土壤含水量条件不同,水分胁迫的增加会导致作物气孔的收缩,气孔导度降低,从而使得CO2进入受阻,光合作用效率下降,并且严重的水分胁迫会导致光合机构受到一定的破坏,使得作物生理活性下降;光合有效辐射的增加以及水分胁迫的增加都会使得作物气孔导度升高,胞间CO2浓度增加,进而影响作物光合作用的限制发生转换,也代表着作物光合作用体系受到了损伤。土壤水分对叶片蒸发速率、气孔导度和胞间CO2浓度影响的思维逻辑关系如图1所示。
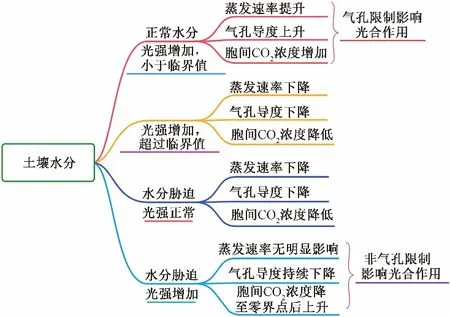
图1 土壤水分对叶片蒸发速率、气孔导度和胞间CO2浓度的影响Fig.1 Effect of soil moisture on leaf evaporation rate, stomatal conductance, and intercellular CO2 concentration
4 土壤水分对作物叶片水分利用效率的影响
叶片是植物进行光合作用的主要器官。叶绿素含量下降是导致后期作物产量下降的主要原因之一。作物对水分的利用效率在节约水资源方面有着不言而喻的重要性,作物净光合速率与蒸腾速率的比值为叶片水分利用效率大小,它表示在每消耗单位重量水分的情况下,固定CO2的多少。魏琼等[41]对蚕豆的光合作用研究结果表明,光合有效辐射增加能够使得水分利用效率在不同土壤含水量条件下增高,在光照条件弱的情况下,水分利用效率随着光合有效辐射的增加而上升直至饱和后,光合有效辐射的增加会导致其下降。范海兰等[42]对短葶山麦冬的光合响应研究表明,在不同水分胁迫下,光合有效辐射的增加会使得叶片水分利用效率上升,当土壤中的水分足够时,叶片水分利用效率基本不受到有效辐射的影响,而当土壤水分缺少时,叶片水分利用效率出现与土壤水分充足时相反的现象。段玉玺等[43]研究沙柳在盐池沙地的光响应表明,不同土壤含水量下,光强的不断增加使得沙柳的叶片水分利用效率增加至最大值,最后出现下降的现象,下降的原因是净光合速率大于蒸腾速率的下降幅度,且在一定水分胁迫下沙柳叶片水分利用效率达到最高值。董梅等[44]对沙棘和银水牛果的研究表明,2种作物在不同水分胁迫条件下都出现随着光合有效辐射的持续增加、叶片水分利用效率上升的现象;光合有效辐射继续增大,则叶片水分利用效率逐渐下降;且2种作物都表现出当存在一定的水分胁迫时,作物叶片水分利用效率达到最高值,叶片的水分利用效率在土壤严重缺水时急剧下降。刘刚等[45]对黄栌的研究表明,随着光合有效辐射的增加,充分供水条件下的叶片水分利用效率低于轻度胁迫下的叶片水分利用效率,且光合速率和叶片水分利用效率都表现为随着光合有效辐射的增加而迅速增加然后趋于平稳,即达到最大值,最后随着光合有效辐射的持续增加开始出现下降趋势。ZHAO[46]和李彦彬等[47]通过研究冬小麦在水分胁迫下的光合响应可知,一定的水分胁迫更有利于冬小麦对水分的利用。
在光合有效辐射的持续增加下,一定的水分胁迫条件会使得作物水分利用效率上升,其原因是,作物的净光合速率增加值高于蒸腾速率;随着水分胁迫的加重水分利用效率开始下降,其原因与前面相反。因而可以得出,作物在适当的水分胁迫下,其水分利用效率达到最大值。
5 展 望
土壤中含水量的不同会对作物光合有效参数带来不同程度的影响。在我国西北干旱地区,日照时间充足,为了更高效地节约水资源,在今后的研究中可以着重探究作物在强光环境下的水分利用效率,尝试通过调整土壤水分状况,使得作物在不同时期净光合速率处于最大值,以至于能够有效积累生长所需能力。郭华[48]等人就开展了土壤水分调控对苹果幼树生理指标的影响研究,试验采用滴灌的方式设置了4个土壤水分下限,分别为田间持水量的50%、60%、70%以及80%,结果显示,叶片光合速率和叶片水分利用效率达到最大值的土壤水分含量是田间持水量的60%。同时也发现灌水量越大气孔导度和蒸腾速率也升高,越不利于节省水分,反而适当的水分胁迫会提高植物的光合速率[49-51]。现代化农业进程中,灌溉农田的方式多种多样,有滴灌、微灌、喷灌,也有精细化灌溉,也就是上升到智能灌溉阶段,有了自动化控制灌溉、专家决策智能系统灌溉,实质上都是需要土壤水分传感器来进行实时监测,给予农田适合于作物生长的水量,以达到植物光合速率的最大程度。基于此,未来通过怎样的设备和系统去调整控制土壤水分来实现对植物光响应的正面作用是一个值得研究的方向。
通过改变土壤含水量使得作物在无光条件下的暗呼吸速率得到加强,从而使得作物吸收CO2生成糖的能力得到加强,进而提高作物的品质。作物气孔导度在一天中处于气孔限制与非气孔限制变化的过程,而气孔开合的大小也会直接影响作物对CO2的吸收,在干旱地区由于日照时间长且温度较高,作物往往会使自己的气孔关闭,使得作物光合作用受到气孔限制的作用,而长时间的水分胁迫以及强烈的光照很可能导致作物光合作用体系受到破环,从而从气孔限制转化为非气孔限制,因此研究作物气孔限制转换到非气孔限制的阈值是未来研究的方向之一,这对作物增产以及作物品质提升有着至关重要的作用。作物水分利用效率的值由蒸腾速率和净光合速率来表示,适当的水分胁迫会使得作物的水分利用效率得到提升,未来可以尝试通过控制土壤水分,进而改善西北地区盐碱荒地无法利用的现状。作物叶绿素作为光合作用的主要色素,大部分结论显示在土壤含水量下降时其总量也随之下降,但也有结论表示其含量在淹灌的情况下会得到提升,探究作物在不同时期土壤含水量的不同对其叶绿素总量的影响状况,当叶绿素总含量改变时,其产量以及质量会否受到牵连是未来研究方向之一。通过在不同时期进行不同量的灌水达到提升作物品质以及产量是研究的最终目的。
从目前研究来看,对于不同土壤含水量条件下作物光合作用的研究还有较大的提升空间,除研究作物光合作用响应的变化外,更可以在提升农作物产量以及品质、节约水资源、改善土壤干旱程度、盐碱地改良方面,开展更为细致、深入、系统的研究,以达到充分节约资源的目标要求。
6 结 论
通过综述近几年来不同土壤含水量对于作物光合响应的研究得出以下结论。
(1)不同水分胁迫条件下,光合有效辐射的增加使得作物净光合速率增大,且当光合有效辐射大于某一阈值时,净光合速率下降。净光合速率、表观量子速率、暗呼吸速率、光补偿点以及饱和点都表现出随着土壤含水量的减少而下降,这也导致作物暗反应过程生成糖的反应受到影响,且重度水分胁迫与强光胁迫共同作用会使得作物光合作用体系受到破环。
(2)作物叶绿素总含量会因为土壤含水量的减少而降低,且叶绿素a、b 的比值减小,类胡萝卜素在不同水分胁迫下含量差异不显著且大于叶绿素含量。
(3)作物的蒸腾速率随着水分胁迫的减少而增加,当土壤含水量高于某一阈值时,作物的蒸腾速率则出现相反现象,水分胁迫以及光合有效辐射在一定范围内增加时,会使得胞间CO2浓度下降,气孔限制值上升,但当水分胁迫以及光合有效辐射超过这个范围时,胞间CO2浓度以及气孔限制值就会出现相反的现象,这也表示光合作用由气孔限制转化为非气孔限制。
(4)土壤含水量是影响叶片水分利用效率的主要因素,在一定水分胁迫条件下,水分利用效率出现上升趋势,其原因是净光合速率大于蒸腾速率;而当土壤严重缺水时,水分利用效率下降,原因为作物净光合速率和蒸腾速率出现了相反的情况。
(5)土壤水分的正确调控对作物的光合特征有效参数有积极影响,对植物的生长发育、对水资源的节约以及对土壤水分与植物光和作用之间的响应关系都有重要的作用。
猜你喜欢
今日农业(2019年14期)2019-01-04
新课程·下旬(2017年7期)2017-08-14
水利规划与设计(2016年9期)2017-01-15
高原山地气象研究(2016年2期)2016-11-10
西北林学院学报(2015年5期)2015-01-03
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
自然资源遥感(2014年2期)2014-02-27
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2012年1期)2012-10-26
