
引进大白菜种质资源表型多样性分析
2023-11-27 10:31:36魏书洞孙晓雪赵建军
中国瓜菜 2023年11期
魏书洞,孙晓雪,孟 川,赵建军,马 卫,卢 银
(1.华北作物改良与调控国家重点实验室·河北省蔬菜种质创新与利用重点实验室·河北农业大学园艺学院河北保定 071000;2.河北省农林科学院经济作物研究所 石家庄 050057)
大白菜(Brassica rapaL.ssp.)原产于我国,是芸薹属芸薹种大白菜亚种的2 年生草本植物,是我国第二大蔬菜作物[1-2]。我国大白菜种质资源十分丰富,截止到2000 年国家种质库保存的大白菜种质达1680 份,且仍旧有新的种质资源不断被搜集、引进和保存[3]。然而,我国大白菜种子市场上却存在严重的同质化问题,不同公司的许多品种无论在商品性、产量、生育期还是在抗逆性等方面都存在相同或相似现象。面对当前蔬菜市场上激烈的竞争形势和人们日益多样化的消费需求,大白菜种质创新迫在眉睫。有效利用已保存的多样化种质资源既是种质创新的重要途径,也可为大白菜基础生物学研究提供更多有价值的材料。
种质资源又称遗传资源,是某一生物种内遗传多样性的全部,主要包括农家种、野生种以及育成品种等[4]。种质资源在自然进化和人工驯化的长期选择中形成,携带丰富多样的表型变异,是育种和生物学基础研究不可或缺的重要材料。实践证明,育种的许多突破性进展往往取决于特殊种质资源关键基因或性状的开发利用[5]。随着科技的迅猛发展,多学科、多领域的先进知识和技术手段被广泛应用于作物育种和生物学研究中,表型性状始终是作物育种和生物学研究的起点和最终落脚点,开展表型性状调查并利用表型数据进行遗传多样性分析是大白菜种质资源高效利用的有效途径,该方法在玉米、萝卜、青花椒、甜瓜、茄子等多种作物上均有报道[6-10]。大白菜中也有相关研究,孟淑春等[11]、范伟强等[2]、韩睿等[12]分别对65、33 和47 份大白菜种质进行了表型性状调查和多样性分析。赵美华等[13]利用表型性状结合分子标记的方法分析了41份大白菜种质的遗传多样性。李国强等[14]对国家蔬菜种质资源库中收集保存的1651 份大白菜种质进行表型数据分析,并筛选获得了248 份核心种质。这些研究报道为大白菜种质资源的表型鉴定提供了参考,但表型性状的调查方法多为传统的观察和测量,调查效率较低,部分重要性状如面积相关性状无法获得。随着科技的发展,表型数据分析软件、图像提取和处理软件、拍照扫描仪器、无人机和机器人等智能化、自动化的软、硬件设施设备被逐步开发并应用于表型性状调查中,大大提高了调查的效率和准确性[15-16]。其中,通过表型图像汲取性状数据的应用较为广泛,马晓丹等[17]利用高通量表型平台获取大豆冠层发育的3D 图像,Merchuk 等[18]开发的Coverage Tool 软件可用于从已获得的植物图像中提取和分析表型数据,但该技术在大白菜表型性状研究中的应用较少,作为重要的叶用蔬菜,大白菜叶片面积、叶片绿叶占比等重要农艺性状尚无研究报道。
在当前国内大白菜种业市场同质化情况日趋严重的情况下,笔者引进了138 份非主栽大白菜种质资源,以传统的调查测定结合创新的图片分析技术汲取了32 个表型性状数据并进行分析,丰富了大白菜的表型调查方法和可调查性状,为大白菜新品种选育、种质改良与创新提供新的材料和表型数据参考,也为大白菜的基础生物学研究奠定坚实的基础。
1 材料与方法
1.1 材料
138 份材料为引自中国北京、河北和荷兰三地的农家种、原始种、野生种等非主栽种质资源,经多代自交纯化后获得的纯合自交系,其中源自北京的种质资源73 份,材料编号W1-W73;源自荷兰的种质资源11 份,材料编号W74-W84;源自河北的种质资源54 份,材料编号W85-W138。
1.2 方法
试验材料分别于2018 年种植于河北省保定市徐水区崔庄镇农家田,2021 年种植于石家庄辛集市的华北作物改良与调控国家重点实验室马兰试验基地,2022 年种植于河北省保定市的河北农业大学农科教学实习基地,试验数据源自三地的调查结果。
试验材料播种于32 孔穴盘内进行育苗,长至4~5 片真叶时定植于露地,每份材料定植30 株,每10 株一组,分为3 组,每组均采取完全随机排列,株距60 cm、行距50 cm,随机选取3 个长势一致的单株为生物学重复,进行表型性状调查。共调查32个表型性状,包括12 个间断性性状和20 个连续性性状,以赋值的形式将间断性性状的调查结果转化为数据。具体的调查时期、部位、方法及赋值标准参考国家标准和已有研究报道并进行补充修改(表1)[19-21]。

表1 大白菜表型性状调查部位、赋值标准及调查方法Table 1 Location,assignment standards,and investigation methods for phenotypic traits of Chinese cabbage
连续性性状中,13 个性状以先拍照再通过图像处理软件分析照片的方式获得数据,拍照在设有标尺的平整黑色背景下以支架固定微单相机(索尼ICLE-6000L)垂直向下拍摄(图1),以ImageJ(1.46 r)软件分析照片汲取相关性状的表型数据;其余7 个性状采用直接测量和人工计数法测定,测定工具包括直尺、游标卡尺和台秤。

图1 不同部位拍摄照片Fig.1 Photos of different parts
1.3 数据处理
利用Microsoft Office Excel(2019)进行数据统计并计算变幅、平均值、变异系数、多样性指数和均匀度指数;采用系统聚类法以Origin(2022)进行聚类和典型材料的提取分析;利用IBM SPSS Statistics(26.0)进行主成分分析和差异显著性分析。
2 结果与分析
2.1 大白菜引进种质资源的表型多样性分析
由表2 和表3 可以看出,32 个表型性状的变异系数、多样性指数和均匀度指数分别为3.64%~52.82%、0.69~2.09、0.68~1.00,可见供试的138 份大白菜种质资源具有丰富的表型变异。通过比较分析,发现连续性性状较间断性性状变异系数低、多样性指数高。
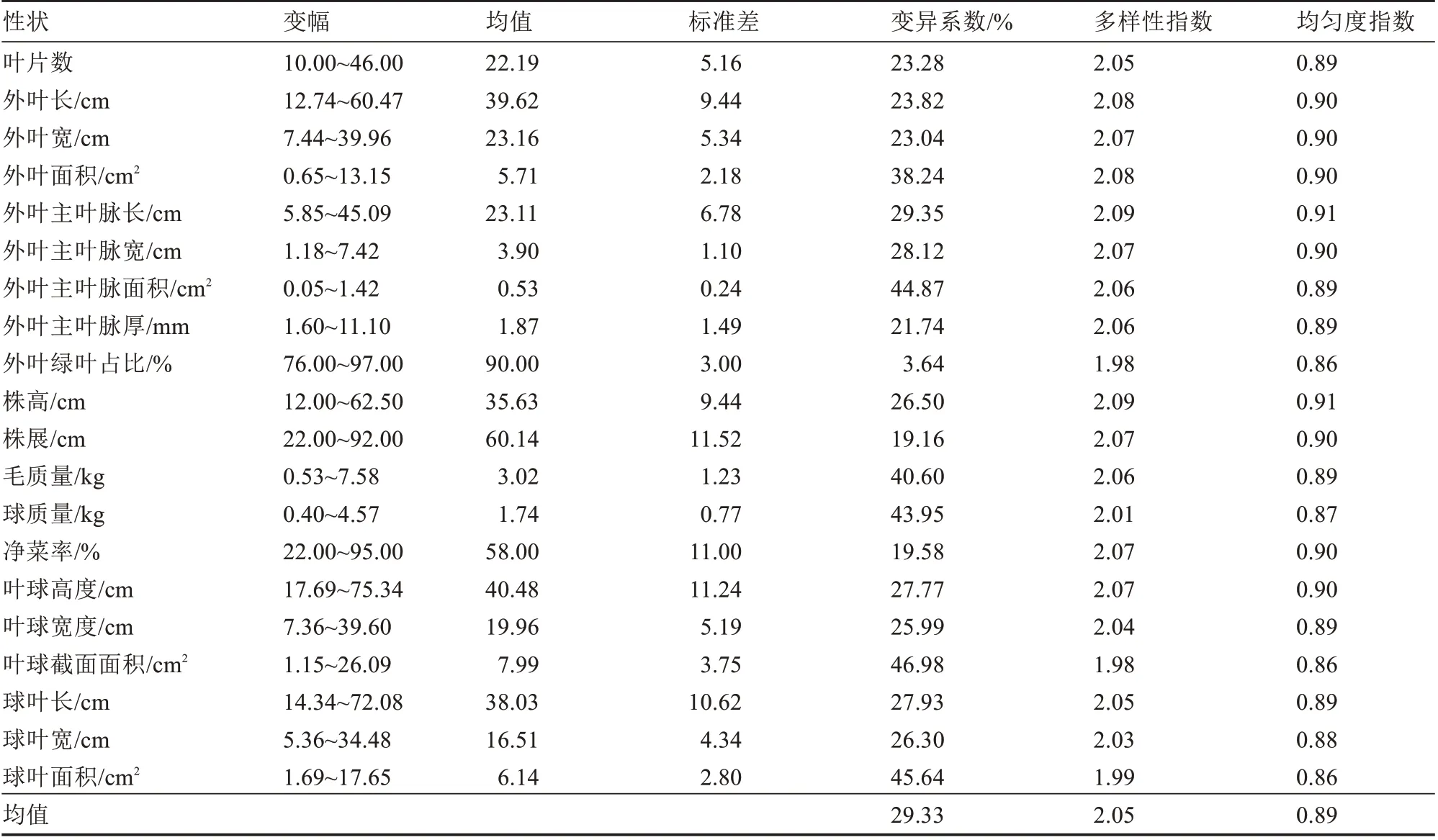
表2 20 个连续性性状的多样性分析Table 2 Diversity analysis of 20 continuous traits
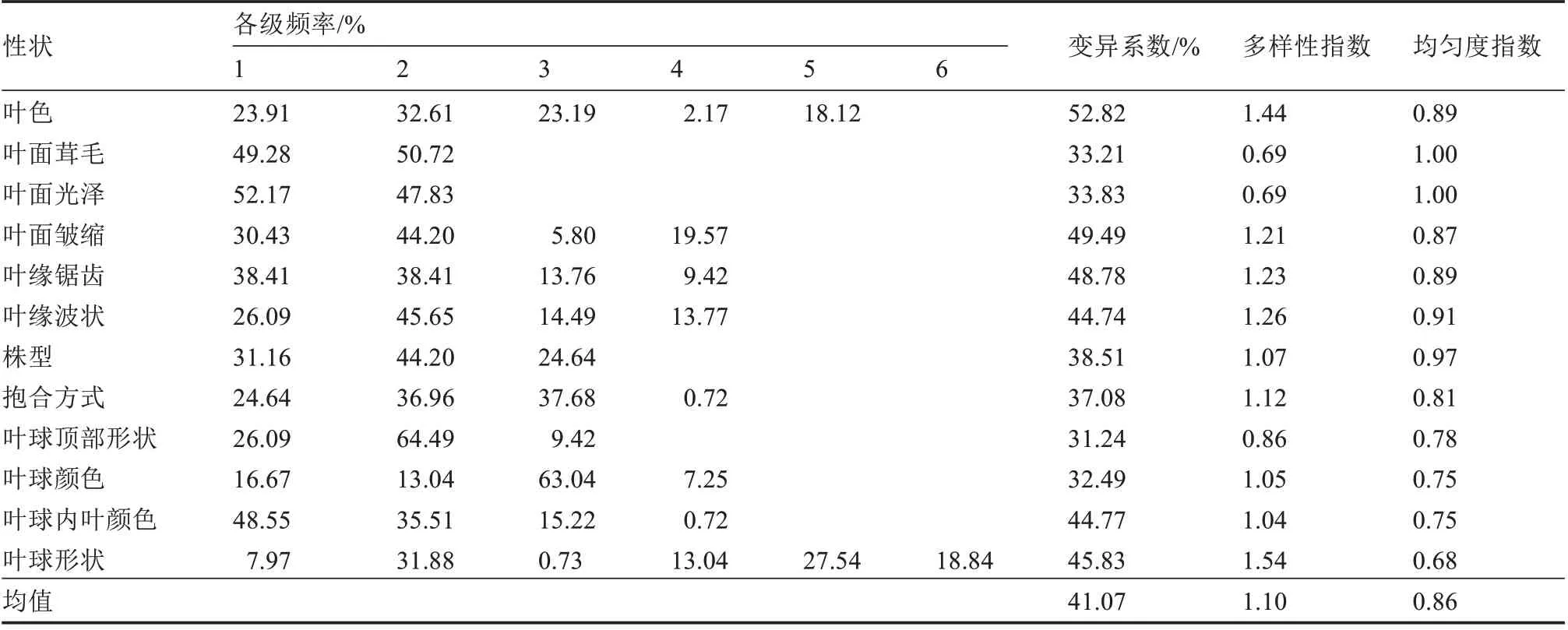
表3 12 个间断性性状的多样性分析Table 3 Diversity analysis of 12 intermittent traits
根据20 个连续性性状多样性分析结果(表2),可知变异系数、多样性指数和均匀度指数分别为3.64%~46.98%、1.98~2.09 和0.86~0.91,均值分别为29.33%、2.05 和0.89,其中叶球截面面积的变异系数最高,外叶绿叶面积占比的变异系数最低;外叶主叶脉长和株高的多样性指数最高,外叶绿叶面积占比和叶球截面面积的多样性指数最低;20 个性状各表型均匀度指数变化幅度较小。
12 个间断性性状多样性结果如表3 所示,变异系数、多样性指数、均匀度指数分别为31.24%~52.82%、0.69~1.54、0.68~1.00,均值分别为41.07%、1.10、0.86,其中叶色变异系数最高,叶球顶部形状的变异系数最低;叶球形状的多样性指数最高,叶面茸毛和叶面光泽的多样性指数最低。138 份材料的7 个性状均匀度指数大于0.85,均匀度较高,5 个性状均匀度指数介于0.68~0.81 之间,具有一定偏好性。偏好性分析发现叶球形状有6 种类型,仅有1 份种质为炮弹形,编号W13;叶球内叶颜色的4 种类型中,仅1 份种质为橘黄色,编号W26;抱合方式的4 种类型,仅1 份种质为拧抱,编号W34,占比仅为0.72%;叶球顶部形状的3 种类型中,89 份种质为圆头形,占比高达64.49%;叶球颜色的4 种类型中87 份种质为浅绿色,占比高达63.04%。
2.2 不同来源大白菜种质资源的表型多样性比较
对中国北京、河北和荷兰三地大白菜种质资源的表型多样性进行比较,由图2 可知,三地种质的32 个性状变异系数各有不同,除外叶绿叶占比外,其他性状的变异系数均高于11.27%,说明三地种质各性状均表现为一定的表型多样性。其中荷兰种质叶缘波状、外叶绿叶占比、叶球颜色和叶球顶部形状4 个性状的变异系数高于其他两地种质;叶片数、叶缘锯齿、外叶主叶脉厚度、株型及净菜率5 个性状介于其他两地种质之间;其余23 个性状均低于其他两地种质。三地种质各性状多样性指数均较高,其中荷兰种质球叶宽、叶缘波状和叶球颜色3个性状多样性指数高于其他两地种质;叶缘锯齿、株型、叶球内叶颜色3 个性状介于其他两地种质之间;外叶主叶脉面积、株高和球叶面积3 个性状与河北种质相同但低于北京种质;叶面茸毛多样性指数与其他两地种质相同;其余22 个性状均表现为多样性指数低于其他两地(图3)。
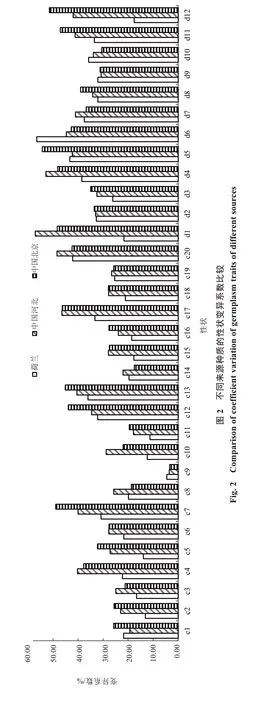
图3 不同来源种质性状的多样性指数比较Fig.3 Comparison of diversity index of germplasm traits of different sources
连续性性状的均值和变幅比较结果如图4 所示,3 地种质仅叶片数和净菜率2 个性状无显著差异,其余18 个性状均存在不同程度的差异。其中荷兰种质的外叶长、外叶主叶脉长、外叶主叶脉面积、株高、株展、毛质量、球质量、球叶长、球叶宽、球叶面积10 个性状显著大于河北和北京两地种质;外叶主叶脉厚度、叶球高度、叶球宽度、球截面面积4 个性状显著大于河北种质;外叶宽、外叶面积和外叶主叶脉宽3 个性状显著大于北京种质;仅外叶绿叶占比1 个性状表现为显著小于北京种质,与河北种质无显著差异。荷兰种质虽多数性状的均值较大但变幅普遍较小,20 个性状变幅均小于北京种质;与河北种质相比,17 个性状的变幅较小,仅外叶绿叶占比、毛质量和球质量3 个性状变幅偏大。间断性性状分级频次比较结果表明(图5),中国北京和河北两地种质不具有明显的偏好性,而荷兰种质以叶色浅绿,叶球筒形合抱为主。各表型中,3 个具有特殊表型的种质(叶球炮弹形、橘黄色球内叶和叶球拧抱)均源自中国北京。

图5 不同来源种质间断性性状分布频次比较Fig.5 Comparison of distribution frequency of discontinaous traits of different sources
2.3 大白菜引进种质资源的主成分分析
主成分分析结果表明(表4),32 个性状可归为10 个主成分,累计方差贡献率为72.70%,可代表这些性状的绝大部分信息。其中,第1 主成分贡献率最高,为20.68%,包括叶球的6 个连续性性状;第2主成分贡献率为12.71%,包括结球期最大外叶的6个连续性性状;第3 主成分包括株高、株型、抱合方式、叶球形状、叶球顶部形状6 个与株型和球形相关的性状;毛质量、球质量等产量相关性状居于第4主成分,其余11 个性状分别居于第5 至第10 主成分。
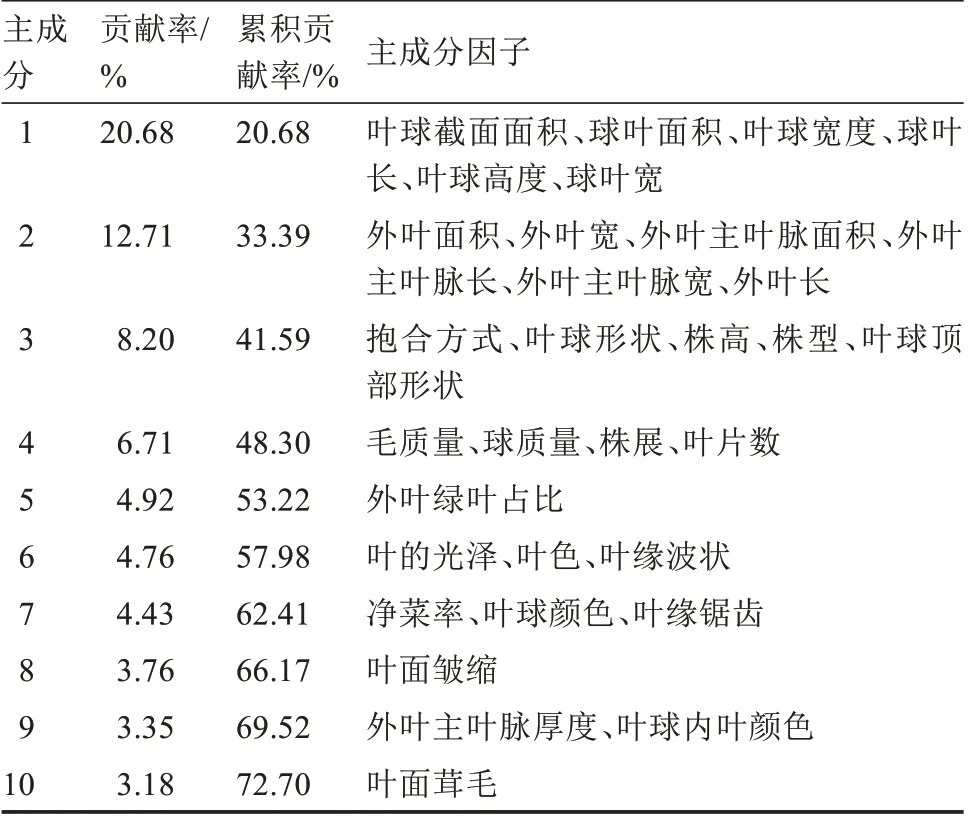
表4 主成分贡献率Table 4 Contribution rate of principal component
2.4 大白菜引进种质资源的聚类分析和典型种质特征分析
聚类分析结果见图6-A,138 份材料分为5 类,分别包括27、37、23、16 和35 份材料,未发现分类与种质来源地相关。经软件分析分别提取了5 个分类的典型种质,I~V类典型种质的编号分别为W67、W49、W113、W105 和W21,表型特征差异明显(图6-B)。
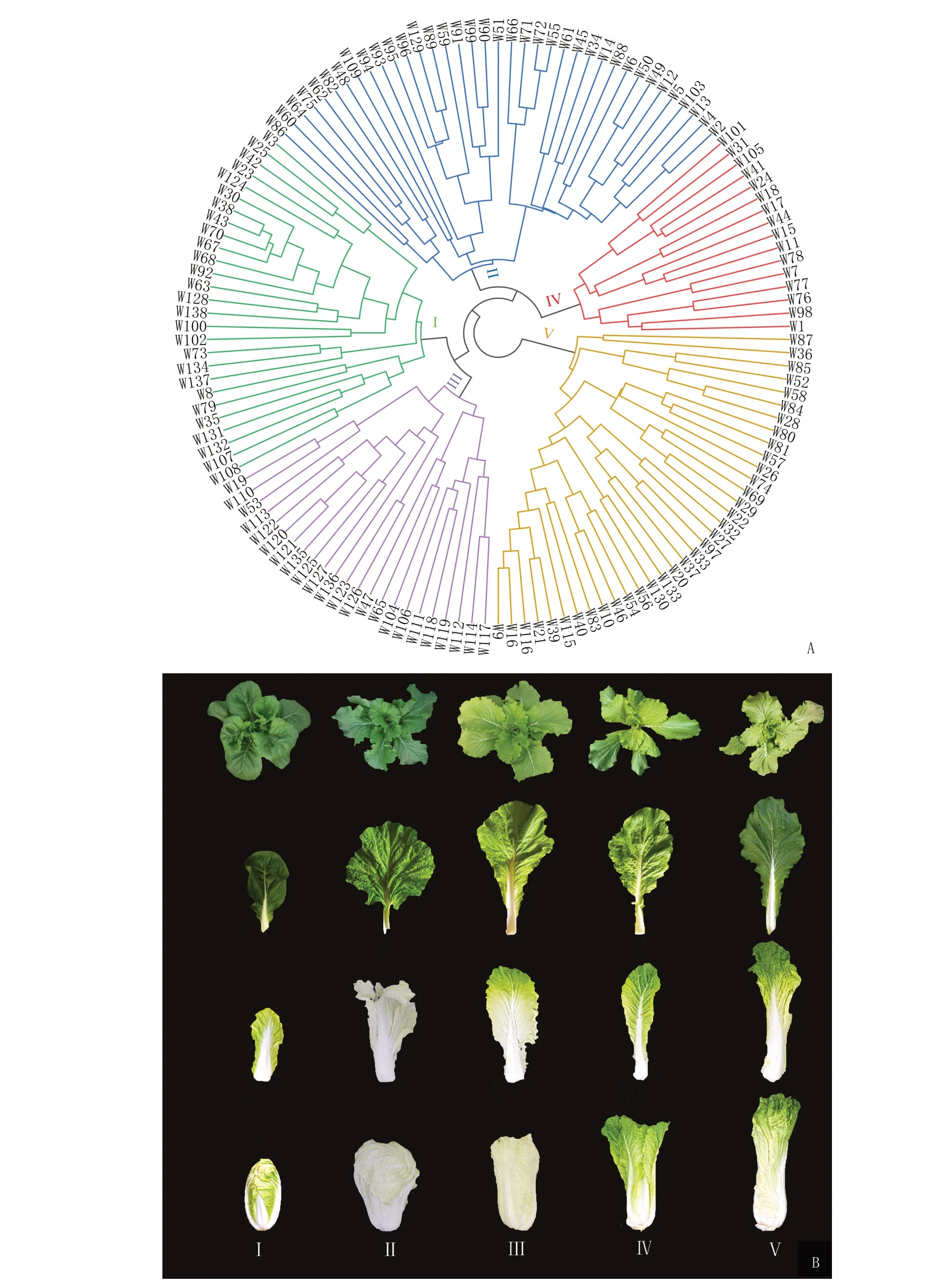
图6 聚类分析(A)与典型特征材料(B)Fig.6 Cluster analysis(A)and typical characteristic materials(B)
各类材料表型数据的差异显著性分析结果见表5,类别I的叶片数、株高、毛质量、球质量、叶球高度、叶球宽度、叶球截面面积、球叶长、球叶宽和球叶面积10 个植株大小相关性状除株高与类别Ⅱ差异不显著外,其他性状均表现为显著变小,可归纳为小株型;类别Ⅱ的外叶长、外叶宽、外叶面积、外叶主叶脉长、外叶主叶脉宽、外叶主叶脉面积、株高和株展8 个外叶相关性状,除外叶宽与类别Ⅳ、株高与类别I差异不显著外,其他性状均表现为显著变小,可归纳为小外叶型;类别Ⅲ外叶宽、外叶面积、外叶主叶脉宽、毛质量、球质量和净菜率6 个性状均表现为最大,可归纳为高产型;类别Ⅳ叶片数、株展、球叶宽、球叶面积较大但外叶宽和净菜率却表现为最小,可归纳为叶球松散型;类别V的外叶长、外叶面积、外叶主叶脉长、外叶主叶脉宽、外叶主叶脉面积、外叶主叶脉厚度、株高、株展、叶球高、叶球宽、叶球截面面积、球叶长和球叶宽13 个性状均表现为较大,可归纳为大株型。数据分析结果与典型特征材料表型一致。
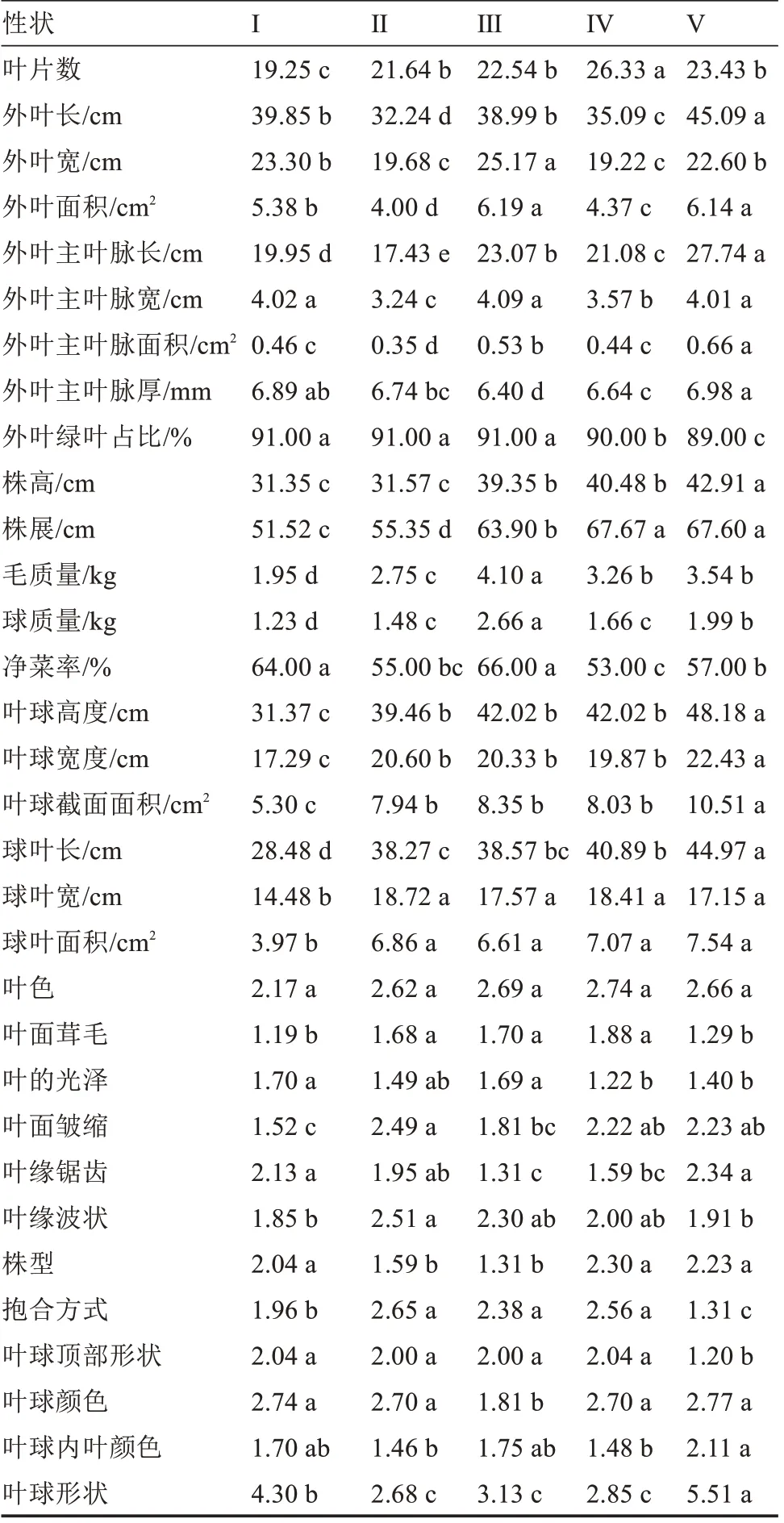
表5 各类材料表型数据的差异显著性分析Table 5 Difference significance analysis of phenotypic data of various materials
3 讨论与结论
变异系数体现性状的变异幅度,多样性指数则体现性状类型的丰富程度,二者均是评价作物表型多样性的重要指标。研究表明变异系数大于10.00%、多样性指数大于1.00,可说明不同样本的多样性丰富[22]。在笔者的研究中变异系数除外叶绿叶面积占比较小外(3.64%),其他性状均大于10.00%;多样性指数除叶面茸毛、叶光泽和叶球顶部形状这3 个间断性性状外,其他性状多样性指数均大于1.00,可见引进的138 份大白菜种质资源具有较丰富的表型多样性。在笔者的研究中,连续性性状和间断性性状相比,连续性性状的变异系数低而多样性指较高,与何伟锋等[23]、赵玉靖等[24]、罗双霞等[25]研究结果一致。分析发现这与性状本身的数值类型有关,连续性性状为实际测定值,不同材料的测定数值各异且数值间差异小,而间断性性状仅有通过赋值获得的几个有限且跨度较大的数值,类型少,不同种质可能具有相同的数值,从而表现为连续性性状多样性指数高,间断性性状变异系数高,因此两类性状表型数据需要分开比较分析,相互之间比较意义不大。均匀度指数可解析性状各类型的分布是否均一,在笔者研究的32 个性状中,27 个性状分布均一,仅5 个间断性性状表现为一定的偏好性,即叶球颜色、叶球内叶颜色、抱合方式、叶球形状和叶球顶部形状,其中叶球炮弹形、球内叶橘黄色和拧抱3 种表型的种质仅各有1 份,叶球浅绿色和圆头2 个性状均占比60%以上,这可能与大白菜的自然进化和人工选育有关。笔者所选试材并非各地主栽品种而是源自野生种、农家种等,推测3 个较少出现的性状类型非自然演化而来,而是经人工选育获得,在原始种质中这些类型较少,如橘黄色球心大白菜是当前市场上的热销类型,多个育种单位进行了有目的的选育;而大白菜球叶因包裹在外叶中见光少多为浅绿色,抱合紧实的叶球多呈现圆头形,较散叶、舒心等类型更耐低温,进而更容易度过冬天进入生殖生长阶段从而繁衍后代,这2 个性状是大白菜自然进化繁衍的结果,因此占比较大。在主成分分析中,第1 主成分为叶球相关的连续性性状,可见叶球在大白菜生产和生长发育中均为最重要的特征性状之一;第2 主成分为最大外叶相关连续性性状,外叶为大白菜营养生长发育阶段进行光合作用的主要功能叶片,与大白菜叶球发育和产量息息相关,聚类分析结果也可证实这一论点,类别Ⅱ的小外叶型材料即表现为株高、株展、毛质量、球质量偏小,而类别Ⅲ的高产型则表现为外叶宽、外叶面积、外叶绿叶占比大。
笔者引入了图片数据提取技术,在叶片及叶球部分的13 个数量性状的测定中摒弃了传统测量方法,大大提高了测定的精准性和高效性,增加了外叶面积、外叶主叶脉面积、球叶面积、叶球截面面积和绿叶占比5 个传统调查方法无法测定的重要性状。此外,笔者拍照所选标尺为具有几个标准色号的色卡,并以此为基础,利用MATLAB 软件分析图片,成功提取了叶色、叶球颜色和球内叶颜色3 个性状的RGB、HSV、LAB 值,已有研究将叶片RGB参数用于预测叶绿素含量,但是如何将这些成组的数据应用于评价大白菜叶色性状中,仍有待深入研究[26]。随着后基因组时代的快速发展,“表型组”成为生物学研究的下一个战略高点,“表型组+基因组”、“表型组+大数据”、“表型组+人工智能”成为未来的发展趋势,更多能解决表型调查中实际问题的硬件设备和软件技术将不断得到研发和应用。
综上所述,笔者引进的138 份国内外的大白菜种质具有丰富的表型多样性,对不同来源地的种质表型进行比较,发现荷兰引进种质特色鲜明,以叶色浅绿、叶球筒形合抱为主,北京和河北两地种质表型呈现多样化,引进种质和本试验表型数据可进一步应用于大白菜种质创新和基础生物学研究中。在大白菜的32 个表型性状中,叶球相关性状是可用于评价、区分描述不同类型大白菜的关键农艺性状,而外叶(非产品叶片)相关性状可影响大白菜的叶球发育和产量,是大白菜育种中的重要性状因子。
猜你喜欢
中南林业科技大学学报(2022年11期)2023-01-12 08:42:04
上海农业科技(2022年3期)2022-06-11 00:50:18
动漫界·幼教365(大班)(2021年11期)2021-11-20 07:49:02
岭南现代临床外科(2021年3期)2021-07-29 13:11:08
长江蔬菜(2021年2期)2021-03-15 08:58:44
医药前沿(2020年12期)2020-08-13 07:00:46
发明与创新(2020年26期)2020-06-27 07:25:06
小学生学习指导(低年级)(2019年9期)2019-09-25 07:43:16
江西农业(2018年13期)2018-02-11 14:32:08
红领巾·萌芽(2017年2期)2017-03-09 15:25:16
