
4 种水产副产品纳豆菌发酵产物的生物活性比较
2023-11-26 09:28:14周更辉耿铭阳马思沅王婷王震宇孙黎明
中国食品学报 2023年9期
周更辉,耿铭阳,马思沅,王婷,王震宇,孙黎明*
(1 大连工业大学食品学院 辽宁大连116034 2 国家海洋食品工程技术研究中心 辽宁大连 116034)
连续20 年,我国稳居水产品产量世界首位,2020 年全国水产品总产量达到6 549.02 万t。随着我国水产品加工技术和水产品加工比例的不断提高,随之产生了数量庞大的下脚料等副产品,对其综合利用已成为水产养殖和加工产业的主要问题之一。这一问题也同样困扰亚洲其它水产养殖大国以及北欧水产捕捞和加工业发达的几个国家[1]。随着对水产副产物中所含各类营养和功能组分研究的深入,采用各种技术从中提取和分离营养和活性组分,并开发营养保健食品,已经成为水产副产物综合利用的主要思路和手段[2-4]。除了直接提取分离法,微生物发酵法也是水产品加工[5-7],水产副产物、废弃物综合利用并开发酶制剂和降胆固醇、抗氧化等功能产品[8-12]的另一大类有效方法。
纳豆枯草芽孢杆菌(简称纳豆菌,Bacillus subtilis natto)因抗拟性强、安全、可产生多种生物活性组分而知名[13]。纳豆菌分泌产生的纳豆激酶经肠道吸收,具有抗血栓、降压、降血脂、抗炎等作用,在心血管疾病的预防和辅助治疗方面功效明确[14],甚至有研究报道其具有潜在预防和治疗老年痴呆和肿瘤的功能[15-16]。纳豆菌能够产生包括蛋白酶、淀粉酶、脂肪酶、凝乳酶等在内的多种高活性胞外酶,其发酵底物也具有较强的宽泛性。此外,纳豆菌还具有肠道菌群和肠道功能的调节作用,为可以广泛而深入使用的优良菌种。在水产加工及其副产物综合利用方面,国内外已有一些纳豆菌发酵的报道。例如:高洁等[17]及陈应运等[18]优化筛选出高产蛋白酶的纳豆菌种,并用于发酵扇贝裙边和蛤蜊,制备高活性的ACE 抑制肽[19-22]。Song等[23]报道纳豆菌发酵蛤蜊所得多肽具有降压作用。此外,纳豆菌发酵还用于氨基酸口服液[24]、海鲜调味料[25]、脂肽[26]的制备等。
截止目前,纳豆菌用于水产副产物综合利用的研究相对较少,更鲜有与纳豆菌的传统底物大豆进行对比的研究报道。本研究选取鲍鱼生殖腺、扇贝裙边、罗非鱼皮和海参卵4 种常见且蛋白含量高的水产副产品为原料进行纳豆菌发酵,并以大豆为对照,对比其发酵产物的纤溶酶活性、抗凝血及ACE 抑制活性,来评价这些水产副产物是否适宜利用纳豆菌发酵开发多肽等功能食品,从而探讨水产副产品综合利用的有效途径。
1 材料与方法
1.1 材料与试剂
生黄豆粉,市售;冷冻新鲜仿刺参卵和冷冻盐渍扇贝裙边,山东省烟台市售;冷冻罗非鱼皮,广西百洋冷冻产品股份有限公司;鲍鱼生殖腺冻干粉,大连獐子岛渔业集团股份有限公司;纳豆菌(Bacillus natto),实验室保存菌种;ACE,实验室常规从猪肺中提取;纤维蛋白原(74 mg/支)、凝血酶(150 BP/支)、蚓激酶(12 600 U/支),中国药品生物制品检定所;98%马尿酰-组氨酰-亮氨酸(Hippuryl-His-Leu-OH,HHL)、98%马尿酸(Hippuric acid,HA)、乙腈(色谱纯),美国Sigma 公司;盐酸、氯化钠等其它常规试剂均为国产分析纯。APTT、TT、PT 试剂盒,南京建成生物工程研究所有限公司。
1.2 仪器与设备
M200 型酶标定量测定仪,瑞士Tecan Infinite公司;UV-5200 紫外分光光度计,上海元析仪器有限公司;KDN-103F 凯氏定氮仪,上海纤检仪器有限公司;E2695 高效液相色谱仪,美国Waters 公司;FE28pH 计,梅特勒-托利多仪器(上海)有限公司;CF16RXII 高速冷冻离心机,日立HITACHI 公司;SCIENTZ-III 数控层析冷柜,宁波新芝生物科技有限公司;YXQ-LS-50SII 立式压力蒸汽灭菌器,上海博讯实业有限公司;SW-CJ-2FD 双人单面净化工作台,苏州净化设备有限公司;HH-4 数显恒温水浴锅,常州智博瑞仪器制造有限公司;YXQ-LS-50SII 立式压力蒸汽灭菌器,上海博讯实业有限公司。
1.3 试验方法
1.3.1 样品前处理 盐渍扇贝裙边于自来水中浸泡5 h,每隔1 h 换水,冻干并研磨成粉;海参卵冻干后打粉;将冷冻罗非鱼皮解冻后去鳞,冻干后打碎成粉。鲍鱼生殖腺冻干粉和黄豆粉直接使用,上述原料于-20 ℃备用。
1.3.2 总蛋白含量的测定 本试验采用凯氏定氮法测定发酵产物中蛋白质的含量。
1.3.3 纳豆菌发酵 将上述底物以30 mg/mL(以蛋白量计)的质量浓度置于三角瓶中,并加入最终浓度为1%的葡萄糖,调节pH 值至7.5,115 ℃高压15 min 后冷却至室温;于瓶中接种体积分数为3%的纳豆菌(终浓度为107CFU/mL),37 ℃培养72 h,间隔12 h 取样。将发酵产物在7 000 r/min、4℃低温离心20 min,收集上清液部分,以测定纤溶酶活性等指标。
1.3.4 纤溶酶活性的测定 参考Lee等[27]的方法测定发酵产物中的纤溶酶活性。将蚓激酶配制成0,1,2,4,6,8 FU/μL 的系列质量浓度溶液,按照同样方法测定纤溶酶活性,绘制标准曲线。
1.3.5 可溶性蛋白含量的测定 采用Folin-酚法测定发酵产物中可溶性蛋白含量。
1.3.6 三氯乙酸可溶性寡肽(TCASOP)含量的测定 采用TCA 沉淀法与Folin-酚法测定发酵液中TCA 可溶性寡肽的含量。将发酵液与20% TCA等比例混匀,静置20 min 后,16 500 g 离心15 min,采用Folin-酚法测蛋白,并计算TCA 可溶性寡肽含量。
1.3.7 抗凝血活性的测定 按照试剂盒说明书,取0.9 mL 正常人混合血浆,加入0.1 mL 发酵产物,用全自动血液检测仪测定发酵产物对血浆活化部分凝血活酶时间(activated partial thromboplastin time,APTT)、凝血酶原时间(prothrombin time,PT)、凝血酶时间(thrombin time,TT)的影响。阳性对照为1 μg/mL 肝素钠,空白对照为生理盐水。
1.3.8 ACE 抑制活性的测定 参考石晓梅等[28]的方法。取25 μL 发酵液,加入同体积的ACE 溶液,漩涡振荡2 min,37 ℃反应5 min。加入50 μL HHL,37 ℃反应1 h 后加入20 μL HCl(0.2 mol/L)。以蒸馏水代替样品作为空白对照组。反应液过0.45 μm 膜后,用高效液相色谱(Waters)检测HA含量。色谱柱为Ghall 12S05-2546 C18(250 mm×4.6 mm,5 μm)。色谱条件为:进样体积10 μL,流动相为乙腈∶水(25∶75,含0.05% TFA),洗脱流速0.5 mL/min,紫外检测波长228 nm。ACE 抑制率(%)=(A0-A1)/A0×100,A0为空白对照组中HA 峰面积,A1为加入抑制剂组中HA 峰面积。
1.3.9 数据统计分析 所有试验均进行3 次重复,测定数据以平均值±标准差表示,使用SPSS 软件采用单因素方差分析(ANOVA)进行数据统计分析,以P<0.05 为显著性检验标准。采用Originpro 2019b 作图。同一原料的不同字母表示差异显著(P<0.05)。
2 结果与分析
2.1 底物中的粗蛋白含量
利用凯氏定氮仪测定样品中的蛋白含量,经测定,黄豆粉、海参卵、鲍鱼生殖腺、扇贝裙边和罗非鱼皮蛋白含量分别为39.17%,60.43%,49.35%,74.72%和95.91%。上述4 种水产副产品中的蛋白含量均较高,是利用纳豆菌分泌的纳豆激酶生产活性多肽的良好原料。
2.2 纤溶酶活性
图1 显示,随着发酵时间的延长,5 种底物的纤溶酶活性呈先升高后降低的趋势,黄豆粉和鲍鱼生殖腺分别在发酵48,60 h 达到纤溶酶活性峰值,而罗非鱼皮、海参卵和扇贝裙边均在36 h 达到最大值。黄豆粉和罗非鱼皮的纤溶酶活性峰值相近且相对最高,约在5 760 FU/mL 左右,扇贝裙边的峰值活性最低。罗飞鱼皮峰值过后,活力下降较慢且相对能够维持较长时间相对稳定。王婷等[29]以海参卵为底物,经过条件优化,纳豆菌发酵产物的纤溶酶活力在36 h 可达到6 723 U/mL 的峰值活性。底物种类的不同很可能是造成纤溶酶峰值时间和活力差异的主要原因,罗非鱼皮和鲍鱼生殖腺都是良好的用于纳豆激酶生产的底物原料。
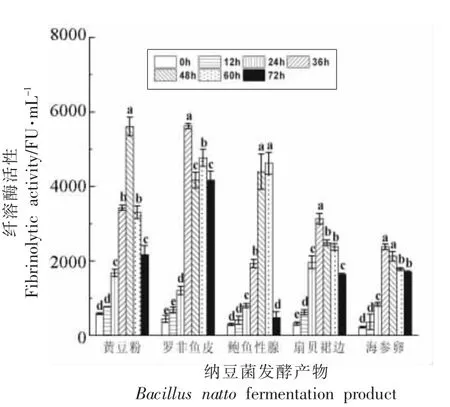
图1 纳豆菌发酵过程中纤溶酶活性变化(n=3)Fig.1 Change of thrombolysis activity during fermented by Bacillus natto(n=3)
2.3 可溶性蛋白含量(SPC)
图2 显示,在发酵0 h,即5 种原料粉经高压灭菌后,上清中SPC 的基数不同,黄豆粉和罗非鱼皮的初始SPC 相对最高,海参卵的SPC 最低。对于蛋白质特性不同的底物,纳豆菌发酵液中SPC呈现不同的变化规律。发酵过程中,黄豆粉、罗非鱼皮和扇贝裙边发酵产物中的SPC 呈先增加后降低的趋势,分别在36,36,48~60 h 达到峰值,而鲍鱼生殖腺和海参卵发酵产物中SPC 则在观察的72 h 内持续上升。与0 h 比,黄豆粉、罗非鱼皮、鲍鱼生殖腺、扇贝裙边、海参卵SPC 峰值分别上升了2.68,1.79,4.31,6.06 和8.63 mg/mL;而发酵72 h后上清中SPC 则分别上升了-0.66,-1.07,4.31,5.73,8.61 mg/mL。由此可见,发酵过程中,纳豆菌通过分泌纳豆激酶(丝氨酸蛋白酶)促进底物中不可溶蛋白向可溶性蛋白转化,同时又有可能将SPC 进一步水解作为菌体自身新陈代谢所用,导致SPC 总量下降。以黄豆和罗非鱼皮为底物时,呈现典型的二段式变化,展示了SPC 生成和被利用的二阶段变化情况;而以海参卵、扇贝裙边、鲍鱼生殖腺为底物时,SPC 的生成量始终大于被菌体利用的量。
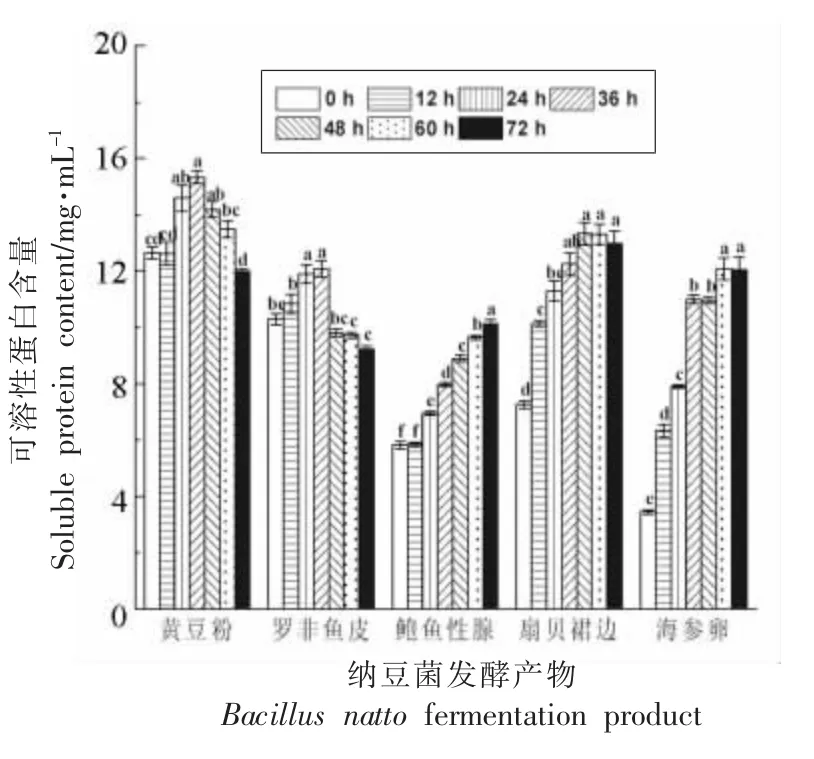
图2 纳豆菌发酵过程中SPC 含量的变化(n=3)Fig.2 Change of SPC during fermented by Bacillus natto(n=3)
2.4 三氯乙酸可溶性寡肽(TCASOP)
图3 显示,5 种底物发酵0 h 的TCASOP 含量均低于1 mg/mL。纳豆菌发酵过程中,罗非鱼皮发酵产物中的TCASOP 含量上升最快,于12 h 就升至5.26 mg/mL,24 h 就升至峰值,是5 种底物中最快到达峰值的且峰值最高。黄豆粉、扇贝裙边、海参卵发酵产物中TCASOP 于24 h 快速上升,均在60 h 到达峰值,之后下降。鲍鱼生殖腺最为特别,发酵过程中TCASOP 始终持续上升,发酵60~72 h 后TCASOP 还呈现快速上升状态。与0 h 比,黄豆粉、罗非鱼皮、鲍鱼生殖腺、扇贝裙边和海参卵发酵产物中TCASOP 峰值分别上升6.62,7.33,5.24,6.42,4.44 mg/mL,这提示罗非鱼皮、扇贝裙边都是适于通过纳豆菌发酵制备活性肽的良好原料。
2.5 凝血酶时间(TT)
图4 显示,与空白对照组比较,鲍鱼生殖腺在0 h 显示出极强的TT 延长作用,远远超过阳性对照肝素钠,但随着发酵时间的延长,对TT 的延长作用减弱,很可能是鲍鱼生殖腺中抗凝活性组分被纳豆菌分泌的纳豆激酶降解所致。60 h 后TT 的延长作用虽然明显下降,但仍与1 μg/mL 肝素钠持平,提示还有其它蛋白酶不能降解的组分发挥抗凝作用。罗非鱼皮发酵产物对TT 的延长作用随发酵时间持续增强,60 h 后接近1 μg/mL 肝素钠的抗凝作用;海参卵的发酵产物也能延长TT,但无明显变化规律。
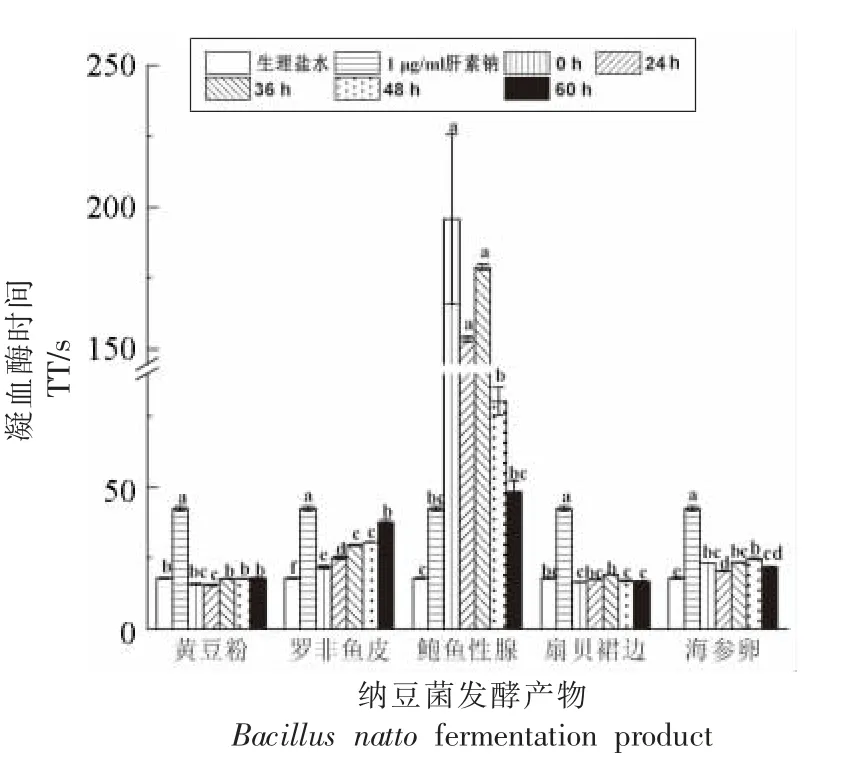
图4 纳豆菌发酵产物对TT 的影响(n=3)Fig.4 Effects of fermentation products by Bacillus natto on TT(n=3)
凝血酶时间(TT)是受检血浆中加入标准化的凝血酶后,血浆纤维蛋白原转化成纤维蛋白所需的时间[30],健康人TT 为14.0~21.0 s。TT 反映的是血浆中凝血酶活性及纤维蛋白原含量的变化。由此推测,鲍鱼生殖腺、罗非鱼皮和海参卵中均有能够直接抑制凝血酶的抗凝组分,而罗非鱼皮中的凝血酶抑制组分能够随着纳豆菌发酵而活性增强或数量增多。
2.6 凝血酶原时间(PT)
图5 显示,鲍鱼生殖腺本身具有凝血酶原时间PT 延长作用,但随着发酵的进行活性逐渐降低。海参卵发酵产物对PT 的延长作用先增强后降低。黄豆粉、罗非鱼皮和陕北裙边的发酵产物均不能延长PT。PT 反映了血浆中外源性凝血途径所涉及的凝血因子Ⅱ、Ⅴ、Ⅶ、Ⅹ的活性以及凝血酶活性是否受到抑制[30],健康人的PT 在9.5~15.5 s。由此推测,鲍鱼生殖腺自身和海参卵发酵产物很可能含有能够抑制上述凝血因子或凝血酶的活性组分。
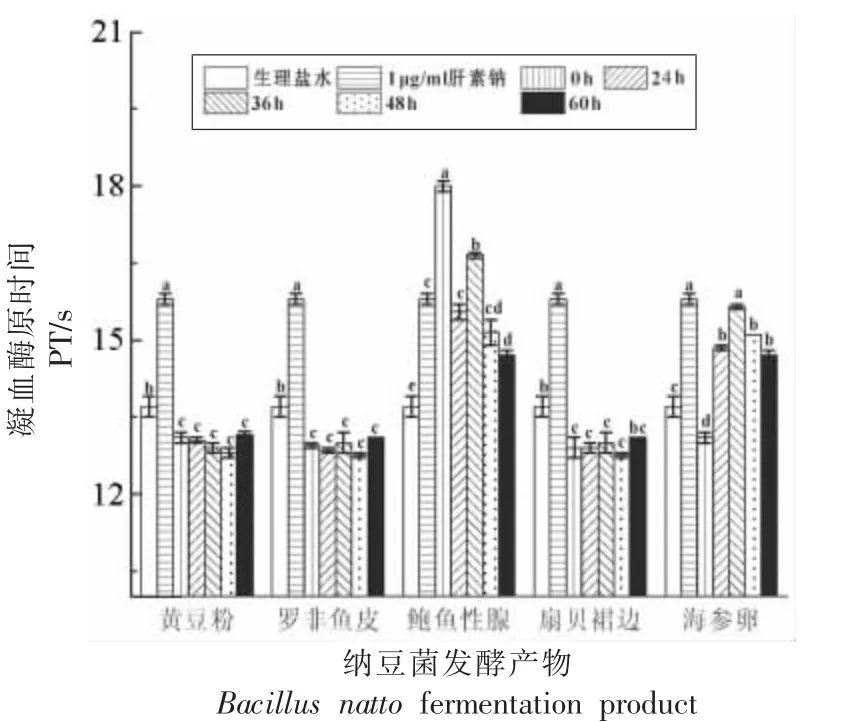
图5 纳豆菌发酵产物对PT 的影响(n=3)Fig.5 Effects of fermentation products by Bacillus natto on PT(n=3)
2.7 活化部分凝血活酶时间(APTT)
图6 显示,鲍鱼生殖腺对APTT 的延长作用很强,且随着在发酵36 h 以内保持稳定,但之后活性显著下降。海参卵对APTT 的延长作用先增强后降低,但总体较为稳定;罗非鱼皮和黄豆粉在发酵0 h 显示出微弱的APTT 延长作用,但随着发酵的进行该活性很快消失。扇贝裙边及其发酵产物对APTT 无影响。APTT 一般用于评价人体内源性凝血途径所涉及的凝血因子Ⅱ、Ⅴ、Ⅶ、Ⅹ以及凝血酶是否出现障碍[30],健康人APTT 为23.0~43.0 s。
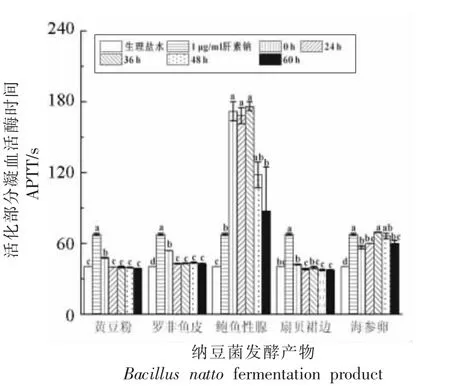
图6 纳豆菌发酵产物对APTT 的影响(n=3)Fig.6 Effects of fermentation products by Bacillus natto on APTT(n=3)
4 种底物中,鲍鱼性腺、海参卵、罗非鱼皮热水提取液自身显示了一定的抗凝活性。其中,鲍鱼性腺的抗凝活性最强。早在1990 年,国内就有鲍鱼提取物抗凝活性的报道[31]。此后,徐美玲等[32]、Li等[33]、Suleria等[34-36]、Zhao等[37]报道了 鲍鱼生殖腺或内脏提取物具有抗凝血活性,Li等[38]在鲍鱼腹足中发现了抗凝血活性组分,上述研究都把抗凝血活性组分指向(硫酸)多糖类物质。但本研究显示纳豆菌发酵会显著降低鲍鱼性腺水浸液的抗凝活性,提示发挥抗凝作用的很可能不只是多糖,本文作者利用中性蛋白酶处理鲍鱼性腺水提取液也发现抗凝活性显著下降,这提示蛋白也很可能发挥抗凝作用。
对纤溶酶(即纳豆激酶)以及纳豆菌其他代谢产品的抗凝活性报道不一。Kurosawa等[39]给志愿者单次口服纳豆激酶2 000 FU 后,TT、PT、APTT数值仍在正常范围内,但血液中D-二聚体含量上升、凝血因子VIII 活性下降、抗凝血酶活性上升。Jang等[40]报道纳豆激酶能够通过抑制血小板聚集而延长凝血时间。Sumi等[41]报道纳豆菌中的吡啶二甲酸与纳豆菌的血小板抑制作用有关。图4~图6 显示,黄豆的纳豆菌发酵产物没有抗凝活性,说明本研究所用菌种产生的纤溶酶以及纳豆菌自身代谢产物并不能延长TT、PT、APTT。由此推测罗非鱼皮发酵产物对TT 的延长作用,也很可能是其自身蛋白的水解所致。对于罗非鱼皮胶原蛋白、明胶以及明胶肽,并未见有抗凝血活性报道。但聂毅磊等[42]发现牛皮明胶的胰蛋白酶酶解产物具有凝血酶抑制活性,作者推测明胶在被胰蛋白酶作用前,其抗凝因子的活性中心被包埋,因而不表现抗凝活性,而被胰蛋白酶作用后,其活性中心得以暴露,形成与抗凝血酶Ⅲ的活性中心序列片段(AGRSLNP)相似的多肽片段(AGRDGNP),抗凝活性得以表现。另外,Suojaranta等[43]报道了明胶对心血管术后病人抗凝血的抑制作用。由此,可以推测,罗非鱼皮经加热灭菌后,胶原蛋白降解成明胶以及少量小分子明胶肽,有微弱延长TT 作用,但在纳豆激酶水解作用下,进一步产生了具有抗凝作用的明胶肽,因此随着纳豆菌发酵的进行,罗非鱼皮发酵产物对TT 的延长作用持续增强(图4)。
2.8 ACE 抑制活性
如图7 所示,5 种底物在纳豆菌发酵过程中均显示出一定的ACE 抑制活性。发酵36~60 h 期间,黄豆粉的ACE 抑制活性略微升高,且一直高于其它样品。其次是扇贝裙边,但其60 h 发酵产物的活性(94.66%)已经超过黄豆粉的活性(91.7%)。鲍鱼生殖腺和罗非鱼皮发酵产物的ACE 抑制活性相对最低。有研究认为水产品中的降压肽相比于其它食品蛋白源的降压肽效果更好[44]。目前,ACE 抑制肽的研究大多数都是基于蛋白酶酶解的制备方法,而采用纳豆菌发酵不但能够生成具有降压作用的ACE 抑制肽,还伴随有其它溶栓等作用的纤溶酶等活性分子。陈应运等[19-20]利用纳豆菌发酵蛤蜊(菲律宾蛤仔)制备ACE 抑制肽,所得多肽的ACE 抑制率为80.49%。王婷等[29]利用纳豆菌发酵海参卵,获得了ACE 抑制率高达90%的发酵产物。高洁等[17]利用纳豆菌发酵扇贝裙边制备ACE 抑制肽,抑制率达到83.7%。Gao等[21]对扇贝裙边纳豆菌发酵产物中的多肽进行了分离纯化并利用大鼠高血压模型验证了多肽的降压作用。由此可见,采用纳豆菌发酵的方法能够用于降压肽的开发。对于罗非鱼皮和鲍鱼生殖腺,纳豆菌发酵产物的ACE 抑制活性较低,尚需进一步优化和确认是否适于进行纳豆菌发酵制备ACE抑制肽。
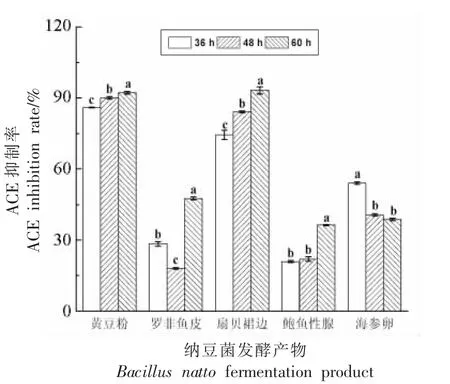
图7 纳豆菌发酵产物对ACE 抑制率的影响(n=3)Fig.7 Effect of fermentation products on inhibition rate of ACE at different time(n=3)
3 结论
对罗非鱼皮、鲍鱼生殖腺、扇贝裙边和海参卵4 种水产品加工副产品进行纳豆菌液体发酵,并与纳豆菌发酵的传统底物黄豆做对照,对比发酵产物中SPC 和TCASOP 的含量、纤溶酶活力、抗凝血活性以及ACE 抑制活性的变化。纳豆菌均能利用4 种副产品合成纤溶酶,其中以罗非鱼皮和鲍鱼性腺为底物合成的酶活力最高;纳豆菌均可将4 种副产品中的未/不溶解蛋白转化为可溶性蛋白及多肽。鲍鱼性腺提取液自身就具有很强抗凝血活性,但发酵过程中活性明显下降。罗非鱼皮和海参卵发酵产物也具有一定抗凝活性,但活性较弱。4 种副产品的发酵产物均有ACE 抑制活性,扇贝裙边活性最强。总之,4 种副产品自身及其纳豆菌发酵产物均在溶血栓、抗凝血、降压这三方面各具特点,对其提取物和纳豆菌发酵产物进行组合应用,有望开发出用于血栓性疾病、凝血倾向和高血压辅助预防和治疗的功能食品,下一步需要将上述功能利用动物实验进行验证和进一步研究。
猜你喜欢
意林彩版(2022年2期)2022-05-03 10:25:08
意林彩版(2022年4期)2022-05-03 00:07:57
家教世界·创新阅读(2019年11期)2019-12-10 09:22:04
文萃报·周二版(2019年2期)2019-09-10 07:22:44
当代水产(2019年3期)2019-05-14 05:43:34
人生与伴侣·共同关注(2018年8期)2018-02-16 06:58:48
汽车实用技术(2017年11期)2017-07-18 12:01:35
小学生作文选刊·低年级版(2015年7期)2015-09-07 09:55:45
作文周刊·小学一年级版(2014年31期)2014-04-29 00:44:03
流行色(2005年3期)2005-04-29 00:44:03
