
异常棉NF-YA家族基因鉴定及其在干旱胁迫下的表达分析
2023-11-25 10:38徐珍珍陈祥龙沈新莲
江苏农业学报 2023年6期
徐珍珍, 季 为, 陈祥龙, 徐 鹏, 郭 琪, 沈新莲
(江苏省农业科学院经济作物研究所/农业农村部长江下游棉花与油菜重点实验室,江苏 南京 210014)
核因子Y(Nuclear factor Y,NF-Y)又称为血红素激活蛋白(Heme activator protein,HAP)或者CCAAT结合因子(CCAAT-binding factor,CBF),是一类能够结合基因启动子特异序列CCAAT的转录因子,普遍存在于酵母、植物和动物中[1]。NF-Y由NF-YA(CBF-A或HAP2)、NF-YB(CBF-B或HAP3)和NF-YC(CBF-C或HAP5)3个亚基蛋白组成[2]。各亚基都具有独特的高度保守核心区,其中NF-YA蛋白具有53个氨基酸组成的保守核心区,包括保守结构域A1和A2。其中A1是NF-YA与NF-YB/NF-YC异源二聚体相互作用的N端,而结构域A2是NF-YA参与DNA结合的C端,在特异识别靶基因启动子CCAAT序列中发挥重要作用[3]。一般情况下,NF-Y调控下游基因表达的机制有3种。第一,NF-YA、NF-YB和NF-YC构成活性异源三聚体,与下游目标基因的启动子CCAAT-box区域结合[4];第二,NF-YB和NF-YC形成异源二聚体NF-YB/NF-YC,然后与特定转录因子结合形成复合物来调节下游基因表达,而NF-YA能够阻止NF-YB/NF-YC二聚体与特定转录因子结合,从而抑制其功能[5-6];第三,NF-Y亚基通过与其他蛋白复合物相互作用来调控下游基因的转录水平[4]。目前NF-YA基因家族在多个物种中已有报道。水稻(Oryzasativa)[7]和拟南芥(Arabidopsisthaliana)[8]基因组中各包含10个NF-YA成员。谷子(Setariaitalica)[9]和玉米(Zeamays)[10]中分别鉴定到10个和14个成员;油菜(Brassicanapus)[11]、大豆(Glycinemax)[12]、鹰嘴豆(Cicerarietinum)[13]、茶树(Camelliasinensis)[14]和桃(Prunuspersica)[15]基因组中分别鉴定出14个、21个、8个、10个和6个成员。越来越多的研究结果表明,NF-YA基因广泛参与植物胚胎发育、种子形成和萌发[16-17]、根的形态建成[18]、开花[19]和光合作用[20]等生长发育过程,响应干旱[9]、高温[21]和高盐[9,21]等逆境胁迫。
棉花(Gossypiumspp.)是世界上重要的纺织工业原料,是饲料蛋白和食用油的重要来源之一,在中国国民经济中具有重要地位。近年来,粮棉争地矛盾的加剧加速了中国棉花生产的区域化,年降水量少、干旱问题十分突出的新疆地区已成为中国棉花主产区。目前广泛种植的四倍体栽培种陆地棉(G.hirsutum)是在正常环境条件下选育出的,抗旱性弱。二倍体野生种异常棉(G.anomalum)生长于撒哈拉沙漠边缘和非洲西南部[22],植株矮小且密被茸毛,叶色浅、叶片小,具有较强的抗旱性[23]。因此,利用异常棉抗旱基因改良栽培种陆地棉资源是中国棉花产业发展的重要方向。目前,异常棉基因组数据已在公共数据库公布[24],但对异常棉抗旱性状相关的NF-YA家族基因分析未见报道。本研究基于异常棉参考基因组信息,利用生物信息学手段对NF-YA家族基因进行全基因组鉴定,分析该家族基因的系统进化、保守基序、基因结构、顺式作用元件、组织特异性表达、干旱胁迫下的表达模式以及该家族基因编码蛋白质的亚细胞定位、理化性质等,旨在为后续深入研究异常棉NF-YA基因家族在干旱胁迫下的功能提供理论依据。
1 材料与方法
1.1 试验材料
拟南芥NF-YA家族基因编码的氨基酸序列来自TAIR(https://www.arabidopsis.org/)网站;异常棉基因组注释的氨基酸和编码序列(CDS)来自Cottongen网站(https://www.cottongen.org/)[21];异常棉根、茎、叶和花的转录组数据以及干旱胁迫下异常棉叶片的转录组数据来自NCBI网站(https://www.ncbi.nlm.nih.gov/)[24-25]。
1.2 异常棉NF-YA家族基因的全基因组鉴定及编码的蛋白质理化性质分析和亚细胞定位
首先在SMART网站(http://smart.embl-heidelberg.de/)预测拟南芥NF-YA蛋白家族的保守结构域,均含有CBFB_NFYA结构域(PF02045),然后在Pafm网站(https://pfam-legacy.xfam.org/)下载种子文件PF02045.18,利用搜索工具HMMER3.1b1检索异常棉氨基酸序列,并将获得的NF-YA蛋白成员在NCBI CDD和SMART蛋白数据库进行验证,最终将通过验证的基因作为异常棉NF-YA家族基因(GoanoNF-YA)并进行命名; GoanoNF-YA蛋白家族成员的等电点和相对分子质量信息利用在线网站(https://web.expasy.org/compute_pi/)进行预测,利用在线软件Cell-PLoc 2.0(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/)进行异常棉NF-YA蛋白家族成员的亚细胞定位预测;利用TBtools软件提取GoanoNF-YA家族基因在染色体的位置信息。
1.3 异常棉NF-YA家族基因的进化树构建、基因结构和顺式作用元件分析
利用MEGA11软件的邻接法(Neighbor-Joining,NJ)构建拟南芥和异常棉NF-YA家族基因的系统进化树,并进行1 000次Boot strap抽样自检;利用GSDS在线软件(http://gsds.cbi.pku.edu.cn)分析GoanoNF-YA基因的外显子和内含子结构;利用MEME软件(http://meme-suite.org/tools/meme)对GoanoNF-YA基因进行保守基序(motif)分析;利用PlantCARE软件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html)分析异常棉NF-YA家族基因启动子顺式作用元件。
1.4 异常棉NF-YA家族基因的表达分析
利用Tophat[26]和Cufflink[27]软件包分析异常棉根、茎、叶和花及异常棉叶片在干旱胁迫下的转录组测序数据Clean data并进行表达量计算,用FPKM(Fragments per kilobase of transcript permillion fragments mapped)表示。提取NF-YA家族基因的表达数据,进行标准化后,利用TBtools绘制GoanoNF-YA基因表达量热图。
2 结果与分析
2.1 异常棉NF-YA家族基因的全基因组鉴定及编码蛋白质的理化性质分析和亚细胞定位
根据异常棉氨基酸数据库共鉴定出17个异常棉NF-YA家族基因,根据基因所在的染色体及位置分别命名为GoanoNF-YA1~GoanoNF-YA17。NF-YA家族基因编码的氨基酸序列长度为190~603 aa,平均315 aa。蛋白质的相对分子质量为21 890.3~66 522.6,平均34 774.3。理论等电点为7.87~10.07,平均8.94,均为碱性。除GoanoNF-YA4定位在叶绿体,其余16个成员均定位于细胞核(表1)。
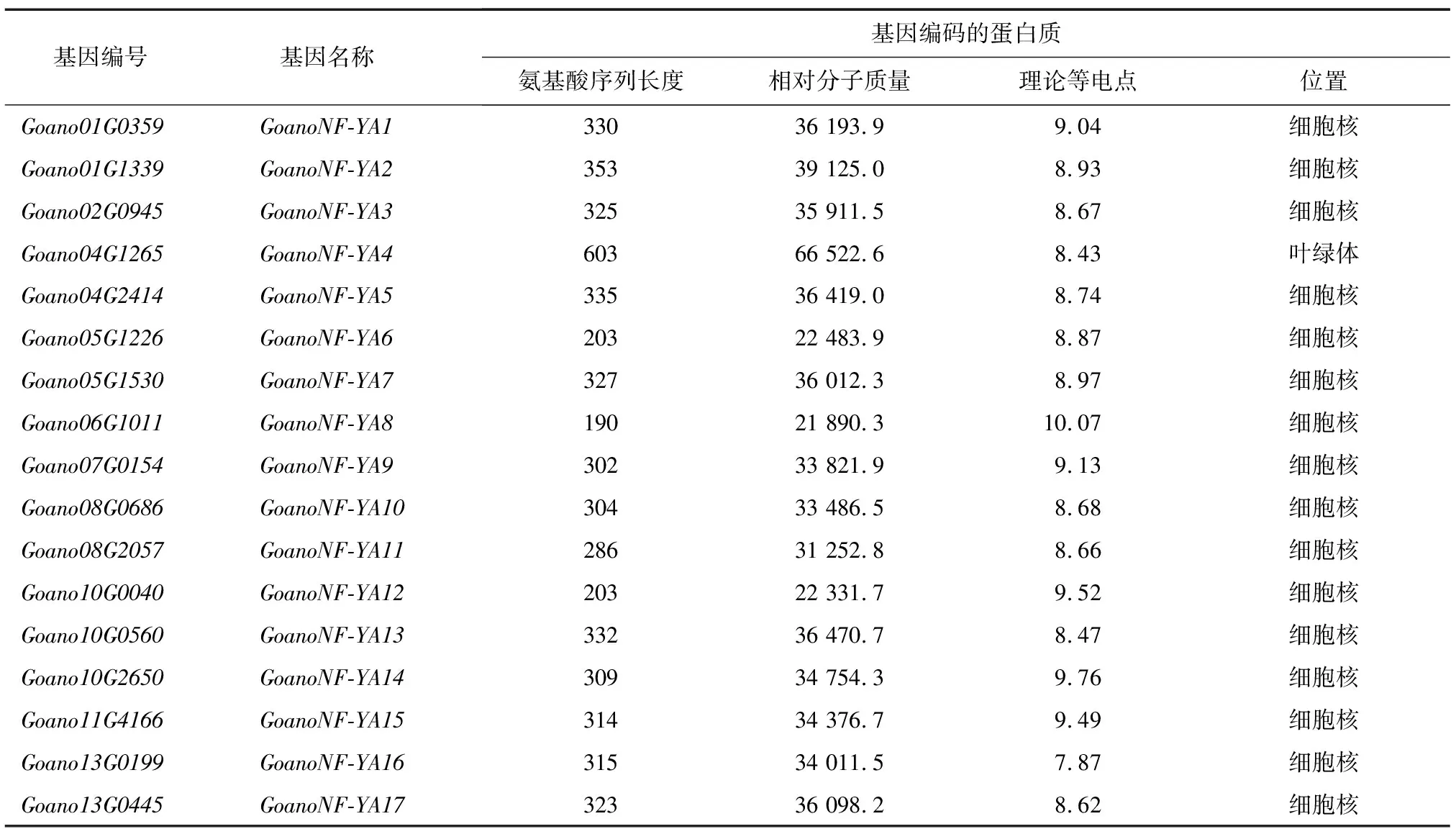
表1 异常棉NF-YA家族基因的基本信息
异常棉NF-YA家族基因分布在10条染色体上。10号染色体上有3个NF-YA基因;1号、4号、5号、8号和 13号染色体上各有2个NF-YA基因;2号、6号、7号和11号染色体上各有1个NF-YA基因; 3号、9号和12号染色体上没有NF-YA基因(图1)。
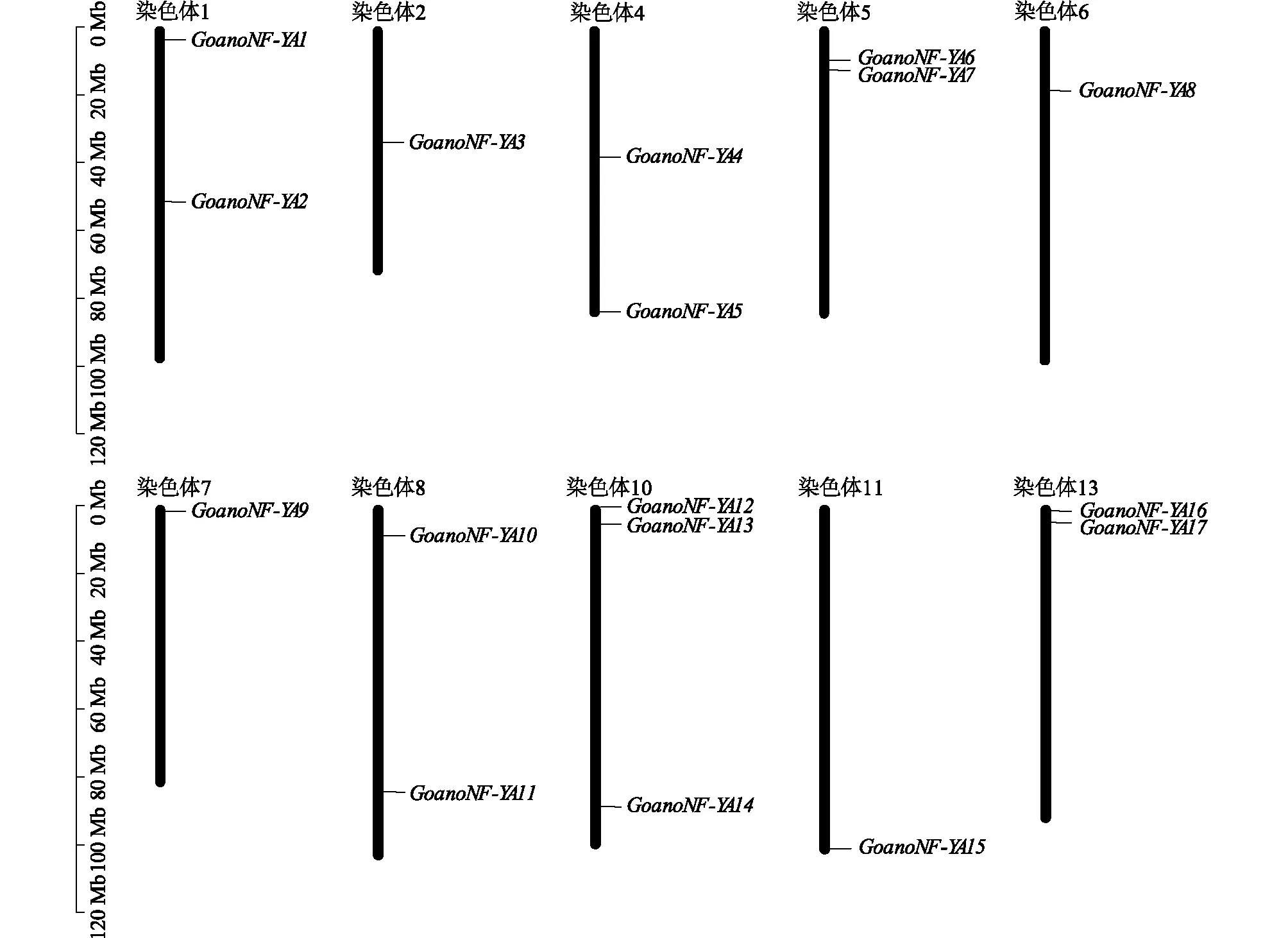
图1 异常棉NF-YA家族基因在染色体上的分布
2.2 异常棉NF-YA家族基因的系统进化树和保守基序
异常棉和拟南芥NF-YA家族基因编码的氨基酸序列比对后构建的系统进化树如图2所示。从系统进化树可以看出,异常棉和拟南芥NF-YA家族基因可分为6类(图2)。Ⅰ类中有5个异常棉NF-YA基因(GoanoNF-YA3、GoanoNF-YA2、GoanoNF-YA10、GoanoNF-YA11和GoanoNF-YA16)和2个拟南芥NF-YA基因(AtNF-YA1和AtNF-YA9)。Ⅱ类中有3个异常棉NF-YA基因(GoanoNF-YA15、GoanoNF-YA4和GoanoNF-YA1)和2个拟南芥NF-YA基因(AtNF-YA2和AtNF-YA10)。Ⅲ类中有2个异常棉NF-YA基因(GoanoNF-YA6和GoanoNF-YA12)和2个拟南芥NF-YA基因(AtNF-YA7和AtNF-YA4)。Ⅳ类中仅有3个异常棉NF-YA基因(GoanoNF-YA8、GoanoNF-YA17和GoanoNF-YA9)。Ⅴ类中有4个异常棉NF-YA基因(GoanoNF-YA5、GoanoNF-YA13、GoanoNF-YA7和GoanoNF-YA14)和2个拟南芥NF-YA基因(AtNF-YA5和AtNF-YA6)。Ⅵ类中仅有2个拟南芥NF-YA基因(AtNF-YA8和AtNF-YA3)。

Motif 1~Motif 10分别为保守基序1~保守基序10。
系统进化树上分支近的NF-YA家族成员具有相同或类似的基序类型、数目和排列顺序(图2)。Ⅰ类中,5个GoanoNF-YA成员(GoanoNF-YA3、GoanoNF-YA2、GoanoNF-YA10、GoanoNF-YA11和GoanoNF-YA16)均具有7个相同类型的保守基序(motif1、motif2、motif3、motif4、motif7、motif9和motif10)且排列顺利相同;Ⅱ类中,GoanoNF-YA15和GoanoNF-YA4基因均具有4个相同类型的保守基序(motif1、motif2、motif3和motif7)且排列顺序相同,而GoanoNF-YA1基因中多了一个motif9,但其余4个保守基序的类型和排列顺序与GoanoNF-YA15和GoanoNF-YA4基因一致;Ⅲ类中,GoanoNF-YA6和GoanoNF-YA12基因均包含5个相同类型的保守基序(motif1、motif2、motif3、motif4和motif10)且排序顺序完全一致;Ⅳ类中,GoanoNF-YA17和GoanoNF-YA9基因含有7个相同类型的保守基序(motif1、motif2、motif3、motif4、motif8、motif9和motif10)且排列顺序相同,而GoanoNF-YA8基因仅包含与GoanoNF-YA17和GoanoNF-YA9相同类型的motif1、motif3、motif4和motif10 4个保守基序;Ⅴ类中,GoanoNF-YA5、GoanoNF-YA13和GoanoNF-YA7基因含有9个相同类型的保守基序(motif1、motif2、motif3、motif4、motif5、motif6、motif8、motif9和motif10)且排列顺序相同,而GoanoNF-YA14仅含有与GoanoNF-YA5、GoanoNF-YA13和GoanoNF-YA7相同类型的4个保守基序(motif1、motif2、motif3和motif5)。
2.3 异常棉NF-YA家族基因的结构
异常棉NF-YA家族基因的结构分析结果表明,除了GoanoNF-YA4含有9个外显子外,其余16个成员含有3~5个外显子,系统进化树上分支近的NF-YA家族成员具有相同或类似的基因结构(图3)。Ⅰ类中,5个基因GoanoNF-YA3、GoanoNF-YA2、GoanoNF-YA10、GoanoNF-YA11和GoanoNF-YA16均含有5个外显子,其中GoanoNF-YA2基因的内含子较长。Ⅱ类中,GoanoNF-YA15和GoanoNF-YA1基因均含有5个外显子,GoanoNF-YA4基因含有9个大小不等的外显子;GoanoNF-YA15基因具有较长的内含子。Ⅲ类中,GoanoNF-YA6和GoanoNF-YA12基因均含有5个外显子,第1个外显子均较短;GoanoNF-YA6内含子较长。Ⅳ类中,GoanoNF-YA17和GoanoNF-YA9基因含有5个外显子,GoanoNF-YA8仅含有4个外显子;3个基因的内含子均较短。Ⅴ类中,GoanoNF-YA5、GoanoNF-YA13和GoanoNF-YA7等基因均含有5个外显子,而GoanoNF-YA14基因仅含有3个外显子;4个基因成员的内含子均较短(图3)。

图3 异常棉NF-YA基因结构特征
2.4 异常棉NF-YA家族基因启动子顺式作用元件
异常棉NF-YA家族基因的相关顺式作用元件分为5类,包括胁迫、激素响应、生长发育、光反应和其他相关元件(图4)。胁迫响应顺式作用元件包括ARE、DRE-core、GC-motif、LTR、MBS、TC-rich repeats和WUN-motif。GoanoNF-YA2、GoanoNF-YA11、GoanoNF-YA16、GoanoNF-YA15、GoanoNF-YA12、GoanoNF-YA17、GoanoNF-YA9、GoanoNF-YA8和GoanoNF-YA14等9个基因含有1~2个干旱响应顺式作用元件MBS。GoanoNF-YA15、GoanoNF-YA1、GoanoNF-YA8和GoanoNF-YA7等4个基因含有1~2个干旱、低温和盐胁迫响应顺式作用元件DRE-core。17个GoanoNF-YA基因都含有抗氧化顺式作用元件ARE,1~7个不等;仅GoanoNF-YA17基因含有GC-motif无氧诱导响应元件。激素响应顺式作用元件包括ABRE、AuxRR-core、CGTCA-motif、GARE-motif、P-box、plant_AP-2-like、TATC-box、TCA-element、TGACG-motif和TGA-element。GoanoNF-YA3、GoanoNF-YA12、GoanoNF-YA8和GoanoNF-YA13等4个基因不含有脱落酸响应顺式作用元件ABRE,其余13个GoanoNF-YA基因都含有ABRE,1~5个不等。生长发育相关的顺式作用元件包括CAT-box、GCN4_motif、HD-Zip、MBSI、Myb-related、O2-site和RY-element。异常棉NF-YA基因启动子分别包含7个CAT-box、1个GCN4_motif、4个HD-Zip、1个MBSI、127个Myb-related、13个O2-site和4个RY-element,涉及分生组织发育、胚乳发育、栅栏组织分化和种子发育的特异调控。参与光反应的顺式作用元件包括AAAC-motif、AE-box、AT1-motif、ATCT-motif、AT-rich sequence、Box 4、CCAAT-box、chs-CMA、circadian、G-Box、GA-motif、GATA-motif、GT1-motif、I-box、MRE、TCCC-motif、TCT-motif和W box。其他顺式作用元件包括TATA-box、CAAT-box、Myc_related、AT~TATA-box、AAGAA-motif和ERE等。

表中数字为启动子区域的结合位点作用元件数目。A1:ARE;A2:DREcore;A3:GC-motif;A4:LTR;A5:MBS;A6:TC-richrepeats;A7:WUN-motif;B1:ABRE;B2:AuxRR-core;B3:CGTCA-motif;B4:GARE-motif;B5:P-box;B6:plant_AP-2-like;B7:TATC-box;B8:TCA-element;B9:TGACG-motif;B10:TGA-element;C1:CAT-box;C2:GCN4_motif;C3:HD-Zip;C4:MBSI;C5:Myb_related;C6:O2-site;C7:RY-element;D1:AAAC-motif;D2:AE-box;D3:AT1-motif;D4:ATCT-motif;D5:AT-rich sequence;D6:Box 4;D7:CCAAT-box;D8:chs-CMA;D9:circadian;D10:G-Box;D11:GA-motif;D12:GATA-motif;D13:GT1-motif;D14:I-box;D15:MRE;D16:TCCC-motif;D17:TCT-motif; D18:W box;E1:3-AF1 binding site;E2:AAGAA-motif;E3:A-box;E4:ACE;E5:as-1;E6:AT~TATA-box;E7:Box III;E8:CAAT-box;E9:CARE;E10:CCGTCC motif;E11:CTAG-motif;E12:dOCT;E13:ERE;E14:GTGGC-motif;E15:Myc_related;E16:Sp1;E17:STRE;E18:TA+G36:G95TA-box;E19:TATA-box;E20:WRE3。
2.5 异常棉NF-YA家族基因的表达模式
异常棉NF-YA家族基因的组织表达模式如图5A所示。从图中可以看出,GoanoNF-YA6、GoanoNF-YA12、GoanoNF-YA11、GoanoNF-YA4和GoanoNF-YA16等基因在根、茎、叶和花中均有较高表达;GoanoNF-YA17、GoanoNF-YA8、GoanoNF-YA13、GoanoNF-YA5、GoanoNF-YA1和GoanoNF-YA7等基因在根、茎和叶中表达量较高,而在花中表达量较低;GoanoNF-YA9、GoanoNF-YA15、GoanoNF-YA10、GoanoNF-YA2和GoanoNF-YA14等基因在叶中表达量最高,根、茎中表达量次之,花中表达量最低;GoanoNF-YA3基因在根、茎、叶和花中几乎不表达。

A:异常棉不同器官NF-YA家族基因的表达模式;B:异常棉NF-YA家族基因在干旱胁迫过程中的表达变化特征。
不同干旱持续时间(0 d、3 d、5 d、7 d和10 d)下,异常棉叶片GoanoNF-YA家族基因的表达模式如图5B所示。从图中可以看出,GoanoNF-YA15、GoanoNF-YA9、GoanoNF-YA16、GoanoNF-YA11、GoanoNF-YA13、GoanoNF-YA12和GoanoNF-YA6共7个基因在干旱胁迫处理下表达量较高,且随着干旱胁迫时间增加表达量呈现上升的趋势;GoanoNF-YA17和GoanoNF-YA8等基因在干旱胁迫处理后,表达量先升高后降低;GoanoNF-YA4基因在干旱胁迫处理后表达量没有明显的变化;GoanoNF-YA1、GoanoNF-YA2、GoanoNF-YA5和GoanoNF-YA10等基因在干旱胁迫处理下表达量较低,但随着干旱胁迫时间增加表达量呈现上升的趋势;GoanoNF-YA14基因在干旱5 d时表达量最高,而在其他时期表达量较低;GoanoNF-YA7基因在干旱胁迫期间表达量均较低,GoanoNF-YA3基因在干旱胁迫期间均不表达。
3 讨论
NF-YA为NF-Y复合体的一个亚基蛋白,虽进化上较为保守,但具有多样性的生物功能[28-31]。本研究在二倍体野生种异常棉基因组中鉴定到17个GoanoNF-YA成员,同其他单子叶植物水稻、谷子和玉米以及双子叶植物拟南芥、油菜、大豆、鹰嘴豆、桃、白菜和茶树等[7-15]一样,NF-YA基因在异常棉基因组中也发生了扩张。这在一定程度上表明,NF-YA家族成员在进化过程中可能存在功能分化和遗传冗余,从而帮助植物在长期的进化过程中形成灵活的、复杂的和多功能的转录调控系统以适应复杂多变的环境。系统进化分析结果表明,拟南芥和异常棉NF-YA家族成员可分为6类,每类2个物种的基因数目不同,且2个物种都存在单独聚类的情况。如Ⅳ类中仅有异常棉NF-YA成员,没有拟南芥相关成员,而Ⅵ类中仅有拟南芥2个成员AtNF-YA8和AtNF-YA3,没有异常棉NF-YA成员。这说明NF-YA基因在物种进化中发生了适应性进化,这与以往的报道[7-15]是一致的。通过保守基序和基因结构预测,系统进化树上分支近的NF-YA家族成员具有相同或相似类型和排列顺序的基序,推测分支较近的NF-YA家族成员间的功能可能存在着相似性。异常棉GoanoNF-YA家族基因中除GoanoNF-YA4外,其余16个成员均包含3~5个外显子,与其他作物具有相似的外显子数目[9-11]。因此,可以认为NF-YA外显子数目在进化上较为保守。
Zhao等[4]研究结果表明,拟南芥NF-YA基因能调控胚胎发生、根系生长、种子萌发、叶片发育、花期和逆境胁迫等生物学过程,这种基因的多样性功能与其启动子的顺式作用元件有关。启动子顺式作用元件通过调控目的基因的转录进而影响植物的生长发育、非生物胁迫响应和光反应等[28-30]。本研究发现异常棉GoanoNF-YA基因5’上游2 000 bp的序列中存在胁迫、植物激素、生长发育和光响应等相关顺式作用元件。与胁迫相关的元件包括干旱胁迫响应元件MBS、脱落酸响应元件ABRE、干旱、低温和盐胁迫响应元件DRE和抗氧化作用响应元件GC-motif等;与生长发育相关的元件有CAT-box、GCN4_motif、HD-Zip、RY-element、O2-site和MBSI等,分别响应分生组织发育、胚乳发育、栅栏组织分化、种子发育、玉米醇溶蛋白代谢调节和类黄酮生物合成等过程;与光反应相关的顺式作用元件有ATC-motif、ATCT-motif、Box4和MRE等。这些结果表明异常棉GoanoNF-YA基因具有多样的生物功能。
NF-YA基因在植物体中呈现多样化的组织表达模式。拟南芥10个NF-YA基因中,有8个基因在胚胎中表达,5个基因(AtNF-YA1、AtNF-YA2、AtNF-YA3、AtNF-YA7和AtNF-YA9)在胚乳中表达[17]。亚洲棉(G.arboretum)的GaNF-YA4基因在种子和纤维发育过程中表达量较高[32]。陆地棉(G.hirsutum)的GhNF-YA18基因在茎、叶、花瓣和雄蕊中表达量较高,而GhNF-YA27基因在茎中表达量较高[33]。小麦中TaNF-YA3、TaNF-YA4、TaNF-YA7和TaNF-YA9等基因优先在胚乳组织中表达[34]。本研究中,异常棉17个GoanoNF-YA家族基因,有5个基因在根、茎、叶和花中均高表达,11个基因在根、茎、叶均较高表达,但在花中表达量较低。这种基因的多样化组织表达模式说明NF-YA基因参与了较多的生物学过程,具有多样化的生物功能,在植物的生长发育和逆境响应中扮演着非常重要的角色。拟南芥AtNF-YA5基因在干旱胁迫14 d后表达量上调,其异位表达增强了植株抗旱性[35]。在拟南芥中过表达大豆GmNF-YA3基因可提高植株的抗旱性[36]。本研究中,异常棉GoanoNF-YA基因在干旱胁迫后,除了GoanoNF-YA7和GoanoNF-YA3外,其余15个基因均对干旱胁迫产生了响应。进化树上与AtNF-YA5同源性较近的GoanoNF-YA13在异常棉干旱胁迫处理下表达量均较高,且随着干旱胁迫时间增加表达量呈现上升的趋势。
4 结论
本研究通过生物信息学手段从棉花野生种异常棉基因组中鉴定出了17个NF-YA家族基因,分布在10条染色体上,编码的蛋白质长度为190~603个氨基酸,理论等电点为7.87~10.07,16个基因包含3~5个外显子。异常棉NF-YA基因与拟南芥NF-YA基因在系统进化树上可分为6类,每类成员具有相似或者相同类型和排列顺序的基序。异常棉NF-YA家族基因具有组织特异性表达模式且参与了干旱胁迫响应,与AtNF-YA5同源性较近的GoanoNF-YA13在干旱胁迫处理下表达量较高,推测其在调控干旱胁迫响应中发挥重要作用。本研究结果为进一步开展异常棉NF-YA基因家族功能研究奠定了基础。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17
电子科技大学学报(2022年5期)2022-10-29
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国生殖健康(2020年4期)2021-01-18
三农资讯半月报(2020年15期)2020-08-25
中国生殖健康(2018年4期)2018-11-06
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
山东农业工程学院学报(2016年6期)2016-12-01
