
鄱阳湖地区扶桑绵粉蚧寄主植物调查及寄生蜂鉴定
2023-11-22 04:46乔艳艳吴珍平魏洪义
江西农业大学学报 2023年5期
乔艳艳,吴珍平,吴 洁,魏洪义
(1.江西农业大学 农学院∕昆虫研究所,江西 南昌 330045;2.江西省经济作物研究所,江西 九江 332100)
【研究意义】扶桑绵粉蚧Phenacoccus solenopsis(Hemiptera:Pseudococcidae)最早于1898 年在美国新墨西哥州公园两种杂草Boerhavia spicata和Kallstroemia brachystylis根部的红火蚁Solenopsis geminata巢穴中被发现并命名[1-2],之后近一个世纪在美国各州之间传播,鲜有报道。1991 年在美国得克萨斯州首次发现扶桑绵粉蚧危害棉花[3-4];21 世纪初期,通过苗木、接穗及果品的调运等方式迅速蔓延至周边国家和地区。2005 年该虫扩散至亚洲、大洋洲、非洲部分国家[6-7],印度、巴基斯坦等主要产棉国受害严重[8-9]。2008 年6 月,我国广州市园林植物扶桑树上首次发现扶桑绵粉蚧危害[10],很快入侵我国大部分省份[11],危险性综合评价值为0.886[12]。【前人研究进展】扶桑绵粉蚧是多食性害虫,截至2015 年,危害全球范围内55 个属的202 种植物,包括大田作物、蔬菜、观赏植物、果树甚至杂草[13]。棉花受害最严重,其次是秋葵、茄子、番茄、芝麻、向日葵[14]。天敌是调节和控制害虫种群数量的重要因子,而寄生蜂在自然条件下控制粉蚧种群数量起着重要作用。报道显示,康长索跳小蜂Anagyrus kamali对木槿曼粉蚧Maconellicoccus hyrsutus的寄生率为66%~100%[15],在26 个非洲国家,劳氏跳小蜂Apoanagyrus lopezi成功将木薯绵粉蚧Phenacoccus manihoti种群密度降低到可接受范围内[16]。全世界已报到的扶桑绵粉蚧的寄生天敌有24 种[14],而我国报道的扶桑绵粉蚧寄生天敌种类不多,调查区域较窄。陈华燕等[17]在广东和海南调查发现,其寄生蜂有4 种,分别是松粉蚧抑虱跳小蜂Acerophagus coccois、班氏跳小蜂Aenasius bambawalei、长崎原长缘跳小蜂Prochiloneurus nagasakiensis和粉蚧广腹细蜂Allotropasp.;李金峰等[18]在广西地区发现扶桑绵粉蚧寄生蜂9 种。其中,班氏跳小蜂是绵粉蚧的伴迁寄生蜂,是目前大部分地区绵粉蚧的优势寄生蜂,2~3 龄若虫和雌成虫的寄生率20%~70%,也是研究扶桑绵粉蚧天敌防治比较集中的方向。而随着扶桑绵粉蚧入侵时间的推移,本地天敌的适应能力会随之增强,天敌种类和数量往往增加[11]。害虫寄生性天敌的分类鉴定是其人工饲养及生物防治的基础,不断挖掘与利用寄生蜂资源,是控制扶桑绵粉蚧种群的主要途径。DNA 条形码技术的发展和普及为物种分类鉴定工作提供了快速有效的研究方法和手段[19]。该技术主要利用线粒体细胞色素C氧化酶亚单位I(mtCOⅠ)的特定标准区域做模板进行物种鉴定,该段基因序列长度适中,相对保守又有足够的变异,且不存在内含子,是蛋白编码基因,便于比对,可通过翻译检测错误,极少出现重组、插入和缺失。最早由Hebert等[20]利用COⅠ作为DNA 条形编码的基础,对11 个门13 320 个物种的mtCOⅠ基因序列进行分析,发现此基片区段能够很好地对被研究物种进行鉴定,对200多个鳞翅目昆虫种类进行DNA 条形码识别,鉴定结果认为Ⅰ基因的特定序列可以成功鉴定每个昆虫个体[21]。随后,该技术也对逐渐应用到膜翅目寄生性昆虫的鉴定,Zaldívar-Riverón 等[22]利每用Ⅰ对墨西哥Chamela-Cuixmala 自然保护区内采集到的468 个矛茧蜂亚科(Doryctinae)寄生蜂进行了分类研究,Darsouei等[23]在对伊朗Mashhad 梨树果园蚜虫寄生蜂和重寄生蜂展开调查,mtCOⅠ序列的结果与形态鉴定结果一致,张泽源等[24]利用mtCOⅠ基因对中国不同地区紫薇绒蚧Acanthococcus(=Eriococcus)lagerstroemiae寄生性天敌进行研究,快速、准确鉴定了9 种跳小蜂,弥补形态分类学的不足。DNA 条形码技术还可以对寄生蜂的隐存种、近缘种、地理分化和寄主转化性的研究[25-28]。
【本研究切入点】鄱阳湖地区是江西省棉花、芝麻等经济作物的主要种植区,而扶桑绵粉蚧及其天敌在江西省的发生情况尚未有系统的研究报道。【拟解决的关键问题】基于扶桑绵粉蚧对主要经济作物棉花、芝麻等经济作物、蔬菜等的严重威胁,在其尚未产生巨大为害前,明确其在鄱阳湖地区发生、分布情况及寄主范围,准确掌握其寄生天敌的种类及寄生特点,可为充分发挥天敌的生态调控作用,降低杀虫剂的使用量,有效防治绵粉蚧提供科学依据。
1 材料与方法
1.1 扶桑绵粉蚧及寄主植物的调查方法
分别于2020 年8 月17—19 日、2020 年9 月6—7 日、2021 年8 月20—21 日、2021 年9 月21—22 日在鄱阳湖地区组织扶桑绵粉蚧专项普查共4 次,采用随机抽样调查方法,结合目测,调查农田及其周边田埂杂草、城镇主干道及河边绿化带、花草种植基地及其周围区域,重点调查经济作物棉花、芝麻以及花卉、蔬菜、杂草。调查时仔细观察植株各部位,当植株上发现扶桑绵粉蚧若干不同虫态的聚集,即认定该植株为其寄主植物。参考张莉丽等[29]记载其危害程度,即按虫口密度划分轻发生(+),每m2寄主植物<7.5 头;中发生(++):寄主植物上虫量少,每m2寄主植物7.5~45 头;重发生(+++),每m2寄主植物>45 头。使用元道经纬相机APP 拍摄记录寄主植物的为害部位,并记录已确定寄主种名或请相关专家鉴定寄主植物。
1.2 寄生性天敌的采集及形态学鉴定
采集僵硬或变色扶桑绵粉蚧的虫体,装入1.5 mL 离心管,带回实验室密闭饲养,每天观察记录寄生性天敌的孵化情况。将羽化的寄生蜂用无水乙醇浸泡,-20 ℃保存备用。在体视显微镜下观察收集到的标本,参考前人研究结果[30-31]进行外部形态初步分类。每个调查点不同形态种选取一定的样本用于开展DNA条形码分子鉴定,并对所选样本拍照,便于后期进一步核对。
1.3 寄生蜂DNA条形码鉴定
选则不同调查点不同形态学物种寄生蜂共52 头,无菌水冲洗后于滤纸上晾干,使用试剂盒DNeasy Blood &Tissue Kit(Qiagen)提取寄生蜂基因组总DNA,过程严格按照试剂盒说明书操作。凝胶电泳检测DNA 质量。将合格的DNA 样品-20 ℃备用,-80 ℃长期保存。目的mtCOⅠ基因片段扩增选用引物对LCO1490:GGTCAACAAATCATAAAGATATTGG,HCO2198:TAAACTTCAGGGTGACCAAAAAATCA[32]。PCR扩增体系25 µL:2.5 µL 10×Ex Taq Buffer(Mg2+plus),2 µL dNTP混合液,0.5 UTaqDNA聚合酶,正反向引物各5 pmol和2.5 µL DNA模板。PCR扩增反应程序为:95 ℃预变性 3 min,94 ℃变性30 s,47 ℃退火45 s,72 ℃延伸1 min,35 个循环;72 ℃延伸10 min。PCR 产物用1%琼脂糖凝胶电泳检测,由上海生工用ABI3730 测序仪双向测序完成。所测序列在Genebank 中进行Blast,根据序列一致性确定对应物种种名或属名,应用MEGA5 计算4 种寄生蜂mtCOⅠ基因序列的种内与种间遗传距离,邻接法构建发育树(Neighbour-joining tree)对样品进行聚类分析。
2 结果与分析
2.1 扶桑绵粉蚧的寄主植物
2020—2021 年,在鄱阳湖周边3 个市级范围的合计16 个县区共43 个调查点开展调查,其中有21 个调查点上发现扶桑绵粉蚧的危害,占调查点的48.83%,该虫在鄱阳湖地区的发生较为普遍,为害的寄主植物共26科59种,主要涉及大田作物、杂草、花卉、木本植物等,危害植物种类较多的依次为菊科Asteraceae、苋科Amaranthaceae、锦葵科Malvaceae 和茄科Solanaceae 等(表1)。其中,棉花Gossypium hirsutum、胡麻Sesamum indicum、马齿苋Portulaca oleracea、辣椒Capsicum annuum、木槿Hibiscus syriacus等植物上发生虫口密度较大,属重发生,非洲菊Gerbera jamesonii、藿香蓟Ageratum conyzoides、茄Solanum melongena、番茄Lycopersicon esculentum、黄花稔Sida acuta和毛马齿苋Portulaca pilosa等植物上中等发生;其他植物上轻发生或零星发生。
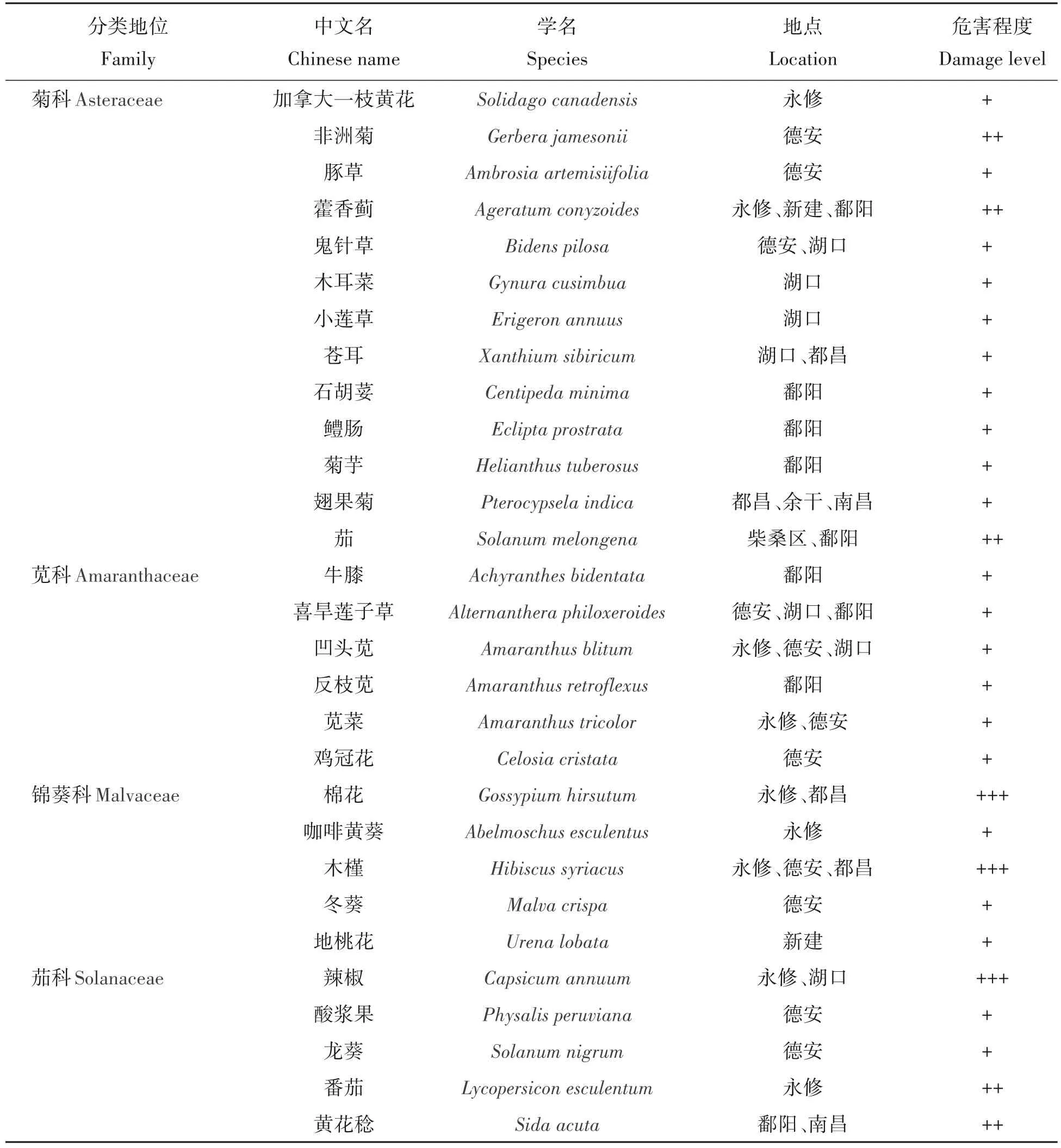
表1 鄱阳湖周边扶桑绵粉蚧发生及寄主植物Tab.1 Occurrence and host plants of P.solenopsis in Poyang Lake
普查中发现,扶桑绵粉蚧在九江市德安县发生面积较大,寄主种类也最多,其次是上饶市鄱阳县、九江市柴桑区等地。在农田如棉花、胡麻等作物上绵粉蚧的种群数量在不同年份间波动较大,其中2020年度在棉田发生危害明显重于2021年度,而在路边、农户自留地和院内花卉等用药较少的区域其发生动态在年度间变化不大。
2.2 扶桑绵粉蚧的寄生蜂种类及发生特点
2020—2021年,各调查点田间采集扶桑绵粉蚧僵蚧共500余头,其中顺利出蜂僵蚧383头,孵化出寄生蜂合计950头(表2),根据孵化特点及镜检,初步归为4个形态种。
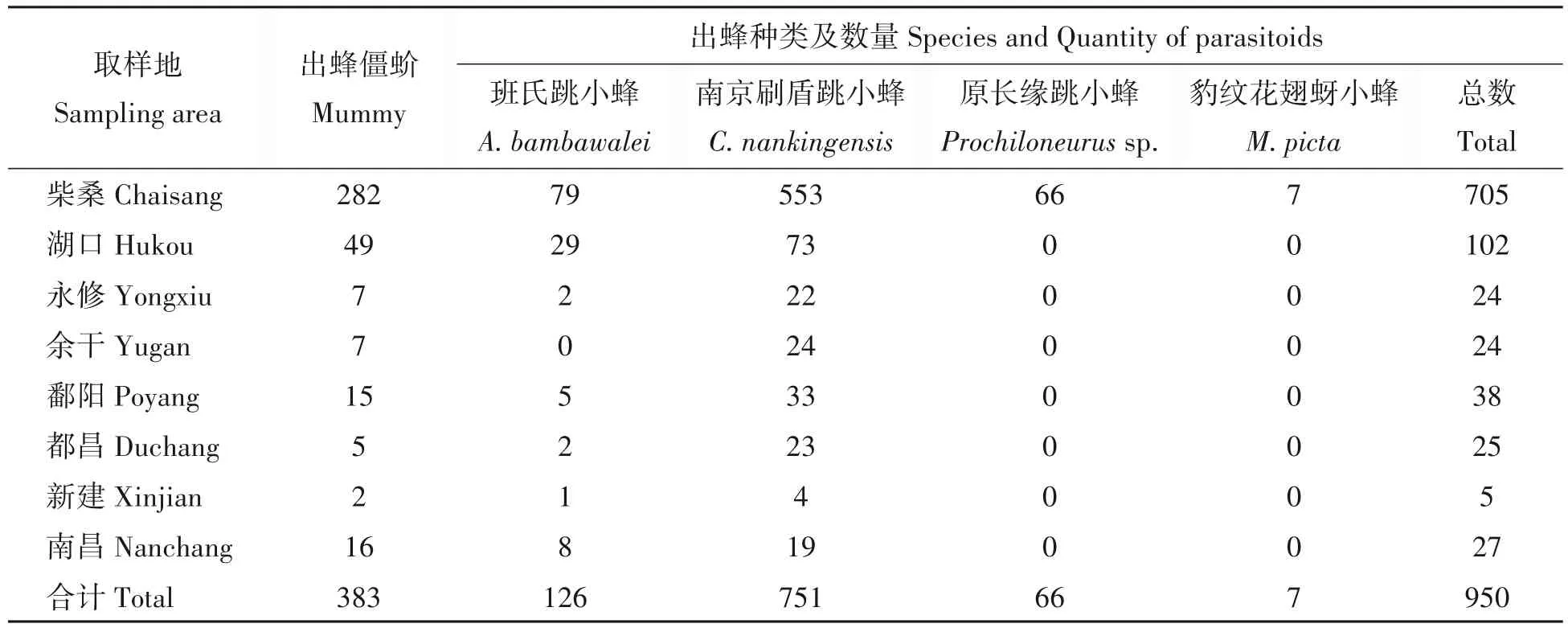
表2 不同取样地扶桑绵粉蚧寄生性天敌种类及孵化数量Tab.2 Species and quantity of parasitoids of P.solenopsis in different sampling area
2.2.1 班氏跳小蜂A.bambawalei(图1a)主要形态特征:雌蜂体长约2.1 mm,体黑色,显微镜下具有金属光泽。复眼大,卵圆形。触角棒状,柄节黄褐色,梗节、第1~2 索节黑色,第3~6 索节、棒节黄褐色至暗褐色,前翅基部褐色,端部透明,后翅全透明。

图1 扶桑绵粉蚧的寄生峰与重寄生蜂Fig.1 The parasitoids and hyperparasitoids of P.solenopsis
2.2.2 南京刷盾跳小蜂Cheiloneurusnankingensis(图1b),主要形态特征:雌性体长约1.5 mm,大部分为黄褐色,头部大,深棕色,具有紫色光泽。触角棒状,柄节、梗节、第1 索节黄褐色,第2~6 索节、棒节黑色。前足淡黄色。
2.2.3 原长缘跳小蜂Prochiloneurussp.(图1c)主要形态特征:雌蜂体长约1.6 mm,体大部分呈黑色;触角柄节为淡褐色 梗节深褐色;第1~5索节白色,第6索节和棒节黑色;前翅基部和端部透明,中间为褐色,后翅透明;产卵器伸出体外部分大于腹部长度的1∕2,为蜡黄色。
2.2.4 豹纹花翅蚜小蜂Marietta picta(图1d)主要形态特征:雌蜂体长约1.0 mm,身体和前翅具暗色和淡色斑纹,呈豹纹状。头部背面宽约等于胸部宽。复眼大,光滑无毛。
2.2.5 寄生蜂的寄生与孵化特征A.bambawalei、Prochiloneurussp.和M.picta为扶桑绵粉蚧的单寄生蜂,每头僵蚧只孵化出1 头寄生蜂;而C.nankingensis为群寄生蜂,每头僵蚧最少孵化1 头寄生蜂,大部分僵蚧孵化2 头及以上,最多孵化15 头,雌雄比大于2∶1。由表2 可知,刷盾跳小蜂和班氏跳小蜂为优势种,二者寄生的僵蚧所占比例分别为47.91%、32.89%,孵化出的寄生蜂分别为751头、126头,出蜂数占全部寄生蜂总量的79.05%、13.26%,这2种蜂分布也最广;原长缘跳小蜂寄生的僵蚧所占比例为17.23%,总出蜂率6.95%,豹纹花翅蚜小蜂的寄生的僵蚧比例为1.83%,出蜂率仅为0.74%,这2 种寄生蜂仅在九江市柴桑区采集到,主要于9月中下旬出现,而班氏跳小蜂与刷盾跳小蜂则伴随着绵粉蚧的出现而出现,寄生时间生态位较长。
2.3 寄生蜂的分子鉴定
从表2 中选取不同地区不同形态种类的寄生蜂共52 头,成功提取DNA 47 条,通用引物PCR 扩增mtCOⅠ基因序列,于Genebank 中进行blasts 搜索,MEGA5 中进行Clustal 多重序列比对,计算碱基组成、变异位点等,用Kimura2-Parameter(K2P)计算序列遗传距离并构建NJ树,结果显示:47条DNA 序列聚类为4个分支,与形态学初步分类结果一致(图2)。
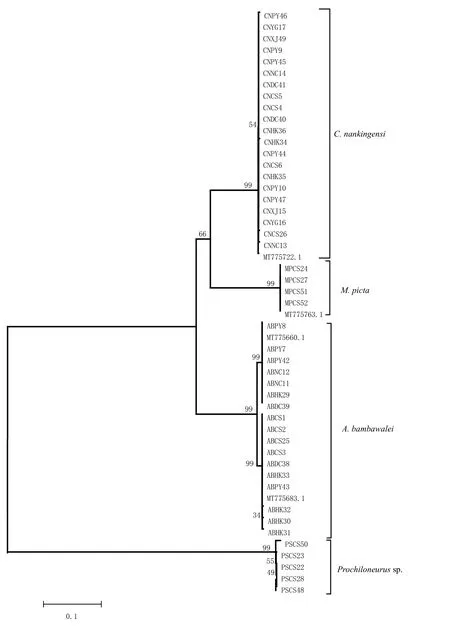
图2 基于寄生蜂和重寄生蜂mtCOⅠ基因序列的NJ树Fig.2 Neighbour-joining tree of parasitoids and hyperparasitoids mtCOⅠsequences,using Kimura-2-parameter distance.Bootstrap values 100 for each haplogroup were calculated in MEGA4.0 with 1 000 replicates
17 条mtCOⅠ基因序列比对截齐后获得609 bp 长度的序列,所测序列的保守位点596 个,变异位点13 个,其中简约信息位点10 个,单一变异位点3 个;序列中A、T、C、G 平均含量为27.5%、42.3%、12.1%、18.1%,A+T 含量69.8%,偏倚A∕T。17 条序列与Genebank 中序列号为MT775660.1 和MT775683.1 的线粒体COⅠ基因序列同源性都高于98.3%以上,聚为一支,鉴定为班氏跳小蜂,是扶桑绵粉蚧的优势寄生蜂,种内遗传距离为0~0.016 0,平均遗传距离0.007 7;21条mtCOⅠ基因序列比对截齐后获得609 bp,所测序列保守位点604个,变异位点5个,其中简约信息位点0个,单一变异位点5个;序列中A、T、C、G平均含量为26.7%、45.3%、12.8%、15.1%,A+T 含量72%,偏倚A∕T。21条序列与Genebank 中序列号为MT775722.1的线粒体COⅠ基因序列同源性都高于99.68%以上,聚为一支,鉴定为南京刷盾跳小蜂,是一种重寄生蜂[33-34],种内遗传距离为0~0.003 5,平均遗传距离0.000 8;4 条mtCOⅠ基因序列比对截齐后获得609 bp,所测序列保守位点609 个,变异位点0 个;序列中A、T、C、G 平均含量为28.9%、47.1%、11%、13%,A+T 含量76%,偏倚A∕T。4 条序列与Genebank 中序列号MT775763.1 的线粒体COⅠ基因序列同源性都高达100%,聚为一支,鉴定为豹纹花翅蚜小蜂,是一种重寄生蜂[33],种内遗传距离为0;5 条形态学为Prochiloneurussp 的寄生蜂mtDNA COⅠ基因序列比对截齐后获得652 bp,所测保守位点644 个,变异位点8 个,其中简约信息位点0 个,单一变异位点8 个;序列中A、T、C、G 平均含量为28.4%、44.8%、14.7%、12.1%,A+T 含量73.2%,偏倚A∕T;种内序列同源性高于99.2%,种内遗传距离0~1.106,平均遗传距离0.004 2,该组在Genebank 中未找到与之匹配的物种,该属种类主要重寄生于半翅目蚧科、粉蚧科的部分跳小蜂上[35]。
3 讨论与结论
扶桑绵粉蚧是一类自身迁移能力较弱的害虫,能够于20年间在全球大范围传播危害,主要与不同地域间种子、苗木、花卉等的频繁调运、绵粉蚧寄主范围广、自身适生性强有关。此次调研发现,扶桑绵粉蚧在江西省鄱阳湖地区发生已较为普遍,关注的各类生境中都有发生,寄主植物涉及大田作物、花卉、园林植物、杂草等多种类别,其中除2019 年在部分棉花种植区域较发生较重外,尚未大面积发生。即仅个别年份的个别田块及地段发生较重,其余调查点均呈点状分布而非成片,散落分布于村庄自留地、路边杂草、家养花卉等化学防治比较薄弱的区域,但存在向周边农田扩散的风险。该害虫尚未在鄱阳湖地区爆发成灾,分析很可能与该虫的寄生蜂天敌有关。
在调研过程中,发现粉蚧危害的调查点,均能采集到被寄生的僵蚧,而且这些僵蚧大部分都成功孵化出寄生蜂。经过形态学初步分类结合DNA 条形码分子鉴定,共发现4 种寄生蜂,其中班氏跳小蜂为初级寄生蜂,也是优势寄生蜂,其余3 种蜂为重寄生蜂。虽然全世界已报道的扶桑绵粉蚧寄生性天敌数量超过24 种[14],很可能有部分种类为重寄生蜂,被误认为是寄生蜂。随着研究的不断深入,部分种类的寄生关系已被重新确认,如本研究中的刷盾跳小蜂,李金峰等[18]在对广西绵粉蚧寄生蜂种类调查研究中曾认定其为绵粉蚧的优势寄生蜂,而陈华燕等[17]对南方9 省粉蚧寄生蜂研究后,将其修正为扶桑绵粉蚧重寄生蜂的优势种,该蜂由Li 等[34]将其描述并命名为南京刷盾跳小蜂Cheiloneurusnankingensis,并证实其寄生于扶桑绵粉蚧的初级寄生蜂班氏跳小蜂,且其为群寄生蜂,每只僵蚧孵化2 头以上的后代,最多达18 头后代,本文调查结果与上述研究结果一致。在整个调研过程中,该蜂几乎都伴随班氏跳小蜂的出现而出现,甚至孵化出该蜂的僵蚧率(47.91%)高于孵化出班氏跳小蜂寄的僵蚧率(32.89%),远远高于另外2 种重寄生蜂豹纹花翅蚜小蜂(0.74%)和原长缘跳小蜂(1.83%)的寄生蜂僵蚧率,在江西余干地区,采集的僵蚧则只孵化出南京刷盾跳小蜂,说明南京刷盾跳小蜂对班氏跳小蜂的种群繁衍存在较大威胁,对其生防控害功能起到巨大的干扰作用。因此,研究南京刷盾跳小蜂、班氏跳小蜂和扶桑绵粉蚧三者之间的相互作用,阻断其对班氏跳小蜂的寄生影响,将是发挥班氏跳小蜂生防功能的关键环节,有必要深入研究。
本研究利用形态学与DNA分子鉴定相结合,清晰准确的将3种膜翅目昆虫鉴定到种,且与Genebank中的物种序列同源性高于98%以上,但仍有一种重寄生蜂mtCOⅠ基因序列无法在Genbank 中获得相似序列,主要原因是Genebank 中收录的mtCOⅠ基因的物种数量相对自然界生物种类还十分有限,这也是DNA条形码在物种鉴定应用的局限之一。
致谢:中国科学院动物研究所张彦周副研究员在部分寄生蜂形态学分类方面给予了帮助,谨致谢意!
猜你喜欢
国画家(2022年1期)2022-03-29
小福尔摩斯·上下五千年(2021年2期)2021-02-21
河南农业科学(2018年6期)2018-01-18
江苏农业科学(2017年7期)2017-05-23
书屋(2017年1期)2017-02-17
西藏科技(2015年3期)2015-09-26
科学大众·小诺贝尔(2015年5期)2015-05-04
湖南农业科学(2012年6期)2012-08-15
西部(2011年5期)2011-08-15
湖南中医药大学学报(2011年11期)2011-03-20
