
平潭海岛黑松群落特征与多样性研究
2023-11-22 08:34:18熊宽洪刘子琳何雅琴郑建忠邓传远
安徽农业大学学报 2023年5期
熊宽洪,肖 兰,刘子琳,何雅琴,魏 凯,郑建忠,邓传远*
平潭海岛黑松群落特征与多样性研究
熊宽洪1,肖 兰2,刘子琳1,何雅琴3,魏 凯1,郑建忠4,邓传远1*
(1. 福建农林大学园林学院,福州 350002;2. 自然资源部海岛研究中心,平潭 350400;3. 广西大学林学院,南宁 530004;4. 福建林业职业技术学院园林系,南平 353000)
为探究平潭海岛黑松()群落特征、多样性水平,科学管理黑松防护林,保护物种多样性,在2018—2020年内,采用样方法对平潭地区8个小型海岛黑松群落进行调查,研究其物种组成、植物区系、群落类型和多样性水平,并采用冗余分析对多样性指数与环境因子关系进行分析。结果发现:(1) 研究区域共有54科117属138种维管束植物,裸子植物与蕨类植物种类较少。(2) 生活型中,高位芽植物(68种)与一年生植物(30种)数量较多。生长型中,乔木8种,灌木39种,草本76种,藤本15种,草本与灌木的植物种类多于乔木。(3) 在地理区系方面,科与属均呈现较强热带性质,但均缺少中国特有分布类型。(4) 通过组间聚类可将黑松样方的植被类型划分为3个群系,9个群丛。(5) 在群系多样性对比中,黑松+台湾群系高于黑松+滨柃群系与黑松群系。在不同层片多样性对比上,草本层与灌木层高于乔木层。(6) 多样性指数与环境因子的冗余分析中,土壤全磷、全钾含量以及坡向对多样性影响显著,总解释量为42.3%。总之,在构建以黑松为主的海岛植物群落时需要注意混交和磷肥使用,根据地区微气候状况合理配置。
黑松;群落特征;物种多样性;冗余分析
物种多样性是物种丰富度与分布均匀度的综合反映,能体现群落的结构类型、组织程度、发展阶段、稳定性以及生境差异[1]。物种多样性是生态学研究的热点,主要内容为物种多样性的发生机制、与环境因子关系、对群落功能的生态影响等[2],研究对象则以大陆地区为主,岛屿地区研究较少。岛屿具有一定封闭性,对保护物种多样性上具有重要作用[3],但岛屿种群分布小,生境恶劣,较大陆种群具有更高灭绝风险[4]。
黑松是松科松属植物,抗逆性强,能适应恶劣的海岛气候,常被用于沿海防护林建设。平潭的自然条件恶劣,生态环境脆弱,原有植被曾多次遭受破坏。20世纪,平潭陆续引种黑松作为防护林,在保持水土、恢复植被等方面取得了良好效果[5],但结构单一黑松群落也导致种群自然更新差,易受病虫害、物种多样性不高等问题。目前有关黑松的研究主要集中在苗木培养[6-7]、病虫害治理与防护[8-9]、外生根菌[10-11]等方向,而针对黑松群落的研究并不多,主要有黑松群落演替特征[12]、黑松群落功能多样性与纬度变异[13],黑松群落物种多样性状况[14]等,平潭海岛的黑松群落物种多样性水平及其与环境因子关系并不明确。本研究对平潭地区的黑松群落进行实地调查,了解黑松群落的物种组成特征及多样性水平,理清影响黑松群落多样性的主要环境因子,为营造海岛黑松防护林、保护黑松群落多样性提供科学参考。
1 研究区概况
平潭位于福建省东部,经纬度为119°32′ ~ 120°10′ E,25°15′ ~ 25°45′ N,面积371 km2,海拔最高438 m。该地区属于南亚热带海洋性季风气候,年降水量900 ~ 1 200 mm,年均温19 ℃左右,气候温暖湿润,雨旱季较分明,降水多集中在3—9月[15],主要植被为黑松、台湾相思()和木麻黄()林等。选取的研究地点为平潭地区的8个小型岛屿,分别为北宫屿、姜山岛、龙母岛、大练岛、大嵩岛、光幼屿、红山屿和小嵩岛。
2 材料与方法
2.1 样方设置与调查内容
在2018—2020年内对平潭地区的8个小型岛屿采用样方法进行实地调查,设置19个面积为20 m ×20 m样方,每个样方四角设置1个5 m × 5 m的灌木和1 m × 1 m草本样方,样方情况如表1所示。分别记录各样方信息如种类、胸径和高度等植物信息以及海拔、坡度、坡向和坡位等地形数据。为了便于数据分析,对坡向与坡位进行转化[16],坡向以北为起点(0°),按照顺时针每45°划分等级并标注数字,如1北坡(337.5° ~ 22.5°)、2东北坡(22.5° ~ 67.5°),数字越大代表越干热,坡位则用1、2、3和4分别代表上坡、中坡、下坡及山谷。对每个样方四角及中心位置使用取土钻进行土壤采样,采样深度为25 cm。根据《土壤农业化学分析方法》[17],使用电位法检测pH值、高温外热重铬酸钾氧化-容量法检测有机质含量、碱解扩散法检测碱解氮含量、碳酸氢钠法检测有效磷含量、乙酸铵提取法检测速效钾含量、凯氏法检测全氮含量、酸融-钼锑抗比色法检测全磷含量、氧化钠熔融法检测全钾含量和电导法检测水溶性盐含量。
2.2 数据处理与分析
按照《中国植物志》[18]与《福建植物志》[19]对植物进行鉴定,根据吴征镒等[20]对植物的科、属分布进行划分。参考Raunkiaer[21]和宋永昌[22]等文献,统计生长型与生活型类别。利用SPSS 25.0的组间连接聚类法对黑松群落样方进行分类。采用Canoco 5对黑松群落多样性与环境因子进行冗余分析(RDA)。
黑松群落的重要性与多样性指数参考张金屯[23]和郑心炫[24]等的方法,计算Patrick丰富度指数()、Pielou均匀度指数(J)、Simpson多样性指数()、Shannon-Wiener多样性指数()与重要值,计算公式如下:
乔木重要值=相对多度+相对频度+相对显著度/3(1)
灌木与草本重要值=相对多度+相对显著度/2 (2)
=(3)
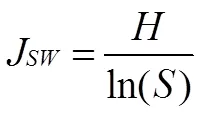
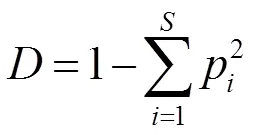
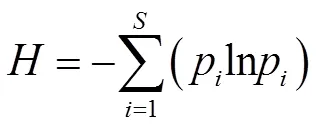
式中,表示样方出现的物种数,p为物种的重要值和全部物种的重要值之和的比值。群落整体多样性的计算参考袁丛军等[25]的方法。
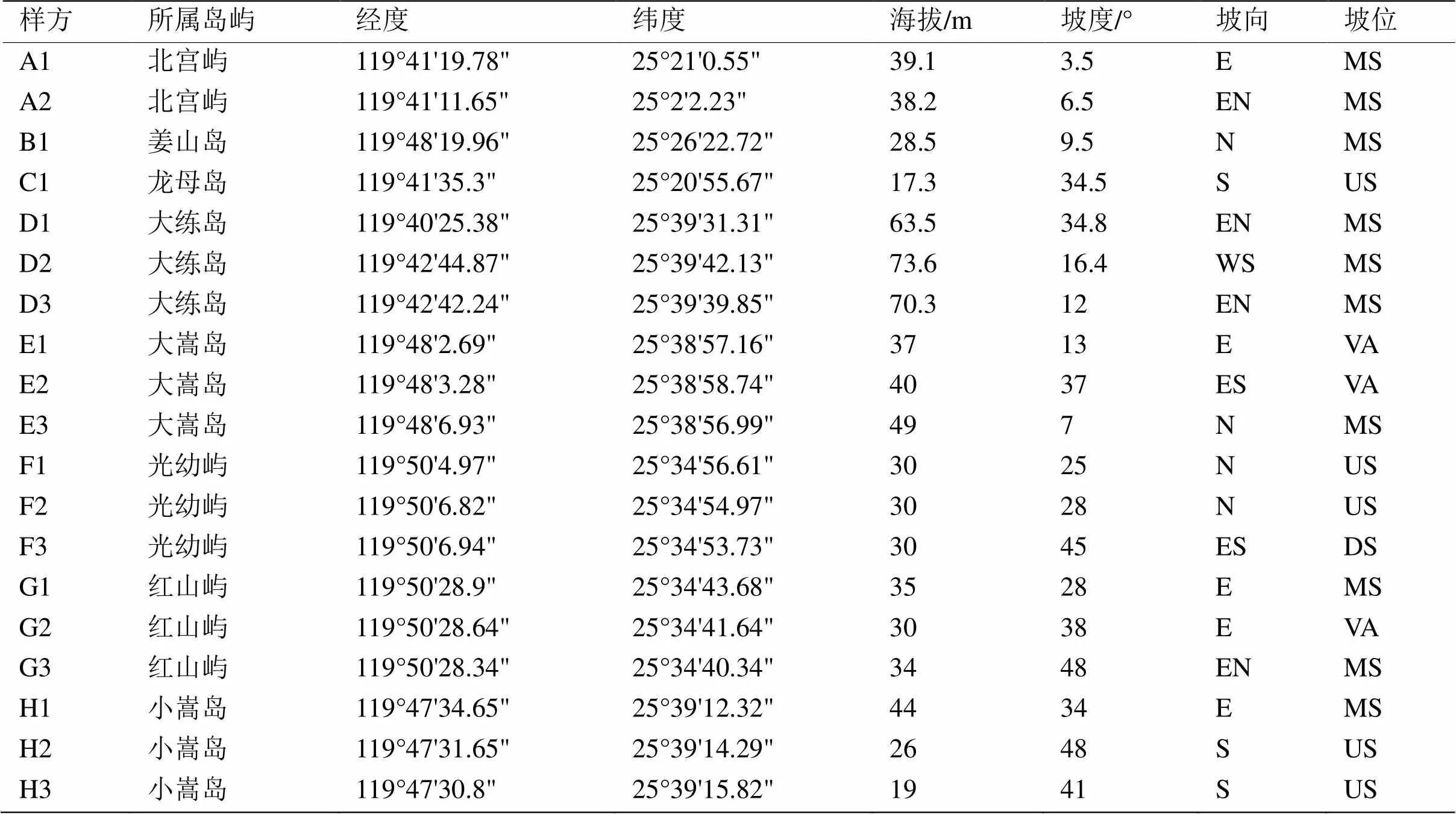
表1 样方概况
注:坡向字母代表, N:北坡;E:东坡;W:西坡;S:南坡;EN:东北坡;ES:东南坡;WS:西南坡;WN西北坡。坡位字母代表,US:上坡;MS:中坡;US:下坡;VA:谷地。
3 结果与分析
3.1 物种组成
样方中包括138种植物,隶属54科117属,其中单子叶4科22属29种,双子叶植物43科88属102种植物,裸子植物仅1科1属1种,蕨类植物6科6属6种。按照植物来源地划分,原生植物共有118种,外来植物20种。以科组成看,禾本科(Gramineae)、菊科(Compositae)和豆科(Leguminosae)等数量较多;从属组成看,则是葡萄属()、鸭嘴草属()等数量较多,仅含1种的属的数量有97个属,优势属不明显。按生活型划分,高位芽植物68种(49.28%),地上芽植物17种(10.14%),地面芽植物14种(12.32%),隐芽植物9种(6.52%),一年生植物30种(21.74%),其中高位芽植物可以细分为小高位芽植物5种(3.62%)与矮高位芽植物63种(45.65%)。按照生长型划分,乔木8种(5.80%),灌木39种(28.26%),草本76种(55.07%),藤本15种(10.87%)。按生活习性划分,阳性植物85种(61.59%),中性植物24种(17.39%),阴性植物29种(21.01%)。
3.2 种子植物地理区系成分
剔除样方中蕨类植物,将剩余48科111属132种种子植物进行区系地理分析,结果如表2所示。除世界分布科外,数量最多的是泛热带分布科,共有18科,占比64.29%,如大戟科(Euphorbiaceae)、葡萄科(Vitaceae)和防己科(Menispermaceae)等,其他热带性质的科仅占个位数。温带性质科的数量也较少,北温带科只有3科,占比10.71%,如松科(Pinaceae)、百合科(Liliaceae)和忍冬科(Caprifoliaceae),北温带和南温带间断分布科只有胡颓子科(Elaeagnaceae),占比为3.57%。
从属的水平看,泛热带性质属的数量最多,有35属,占比为35.35%,如榕属()、蟛蜞菊属()和狗尾草属()等;第2是北温带分布属,具有14属,占比为14.14%,如画眉草属()、蒲公英属()和葡萄属等;其次是旧世界热带分布属,有11属,占比为11.11%,如合欢属()、木麻黄属()和海桐属()等。科与属的热带性质均十分明显,科的R/T值为6.0,属的R/T值为2.0,科的热带成分相比属的热带成分占比更高,但科的分布类型不及属的分布类型丰富,科与属均缺少中国特有分布这一类型。
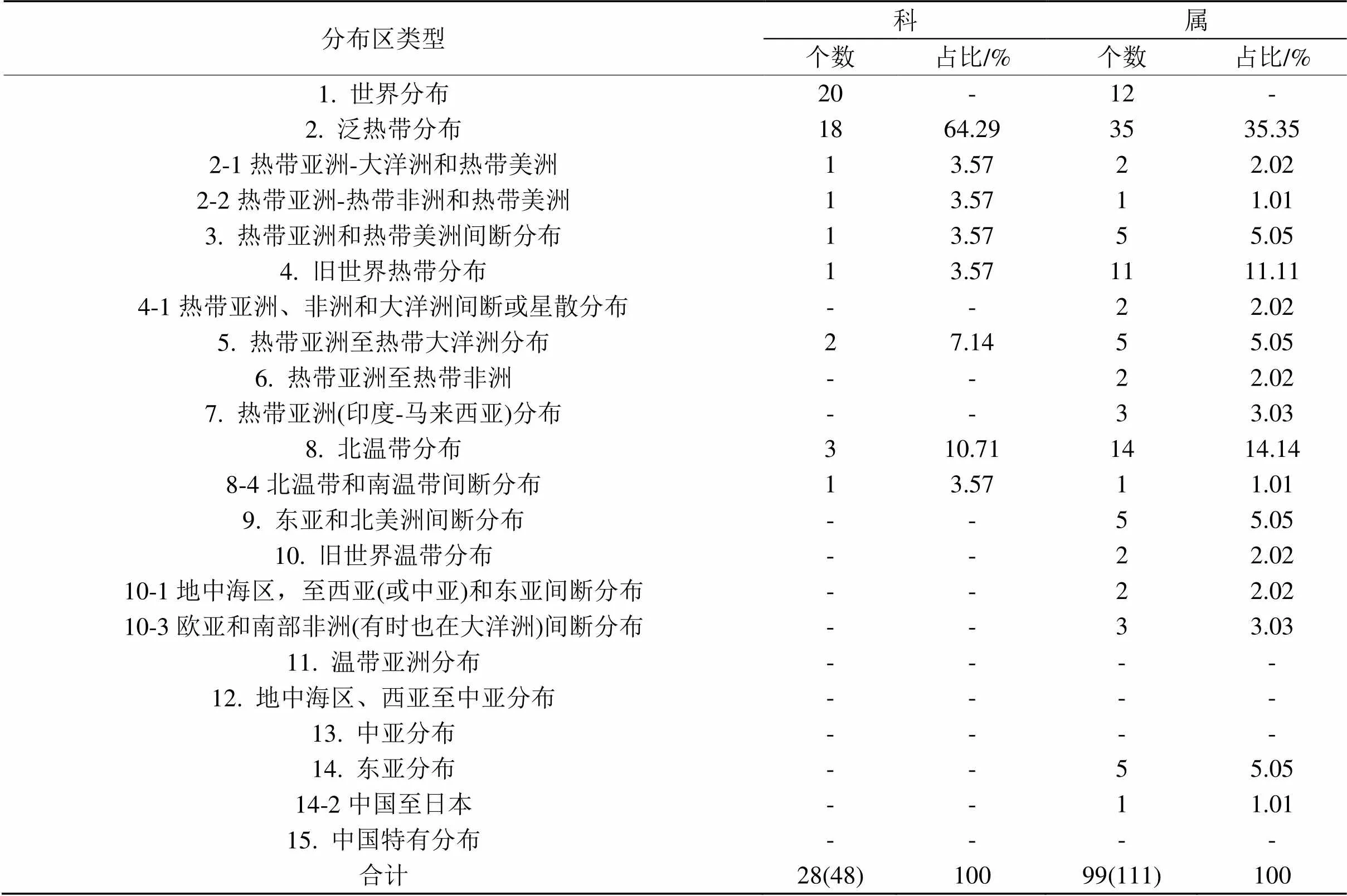
表2 种子植物地理区系分布
注:括号内数量代表包含世界分布类型的科、属总数。

表3 植被分类
3.3 植被的数量分类
剔除样方中频次少于3和重要值小于1的偶见种[26],利用组间连接聚类法进行聚类 (图1)。当欧式距离取21.5时,可以将19个样地划分为3个群系;当欧式距离取10.5时,可以划分为9个群丛(表3)。调查的样地中乔木层优势种主要为黑松与台湾相思,灌木层优势种主要为黑松、滨柃()、铁包金()、算盘子()、车桑子()、雀梅藤()、木麻黄、野牡丹()、山芝麻()和铁包金()等,草本层优势种主要为狗牙根()、华南狗娃花()、天门冬(s)、五节芒()、山菅()、鬼针草()、积雪草()、鸭嘴草()和叶下珠()等。
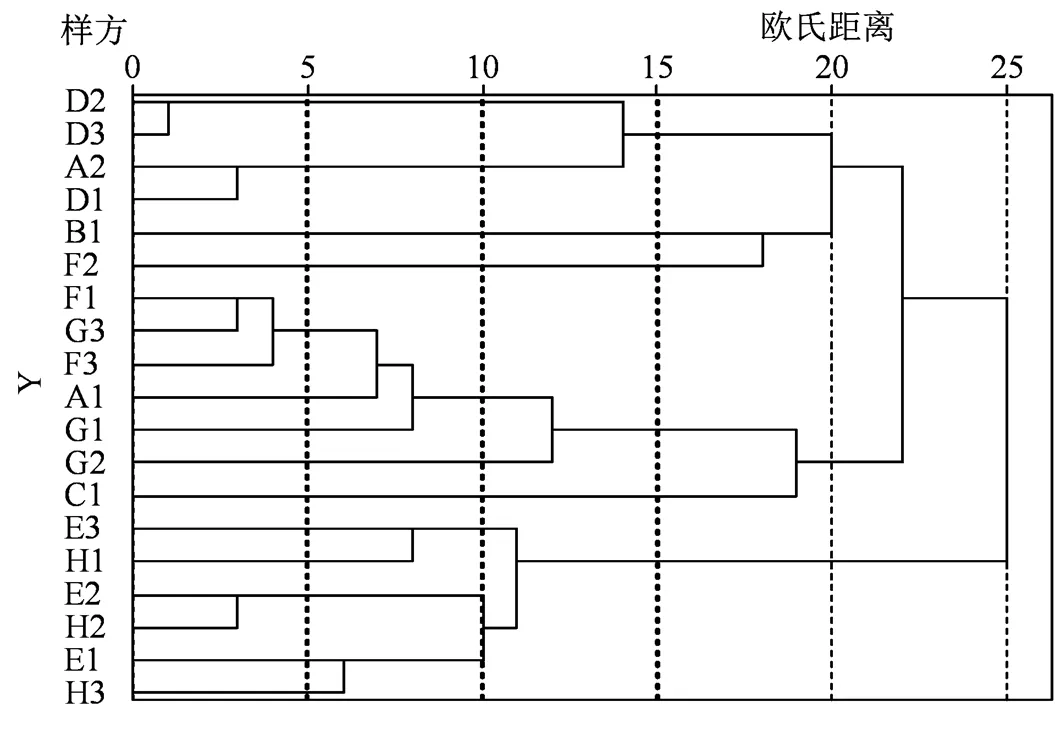
图1 黑松样方聚类图
Figure 1 Clustering map ofsamples
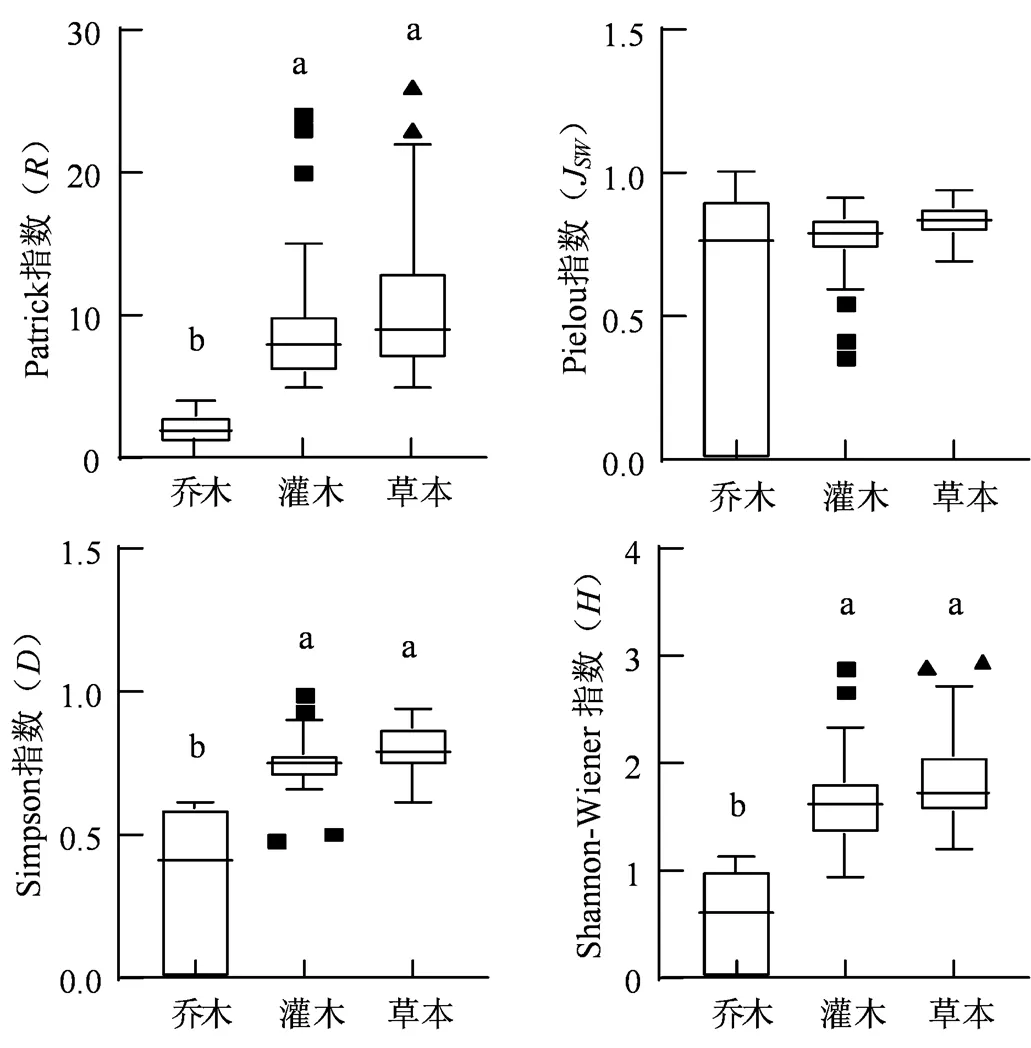
不同小写字母表示差异性显著(P < 0.05)。下同。
Figure 2 Plant diversity of different synusia
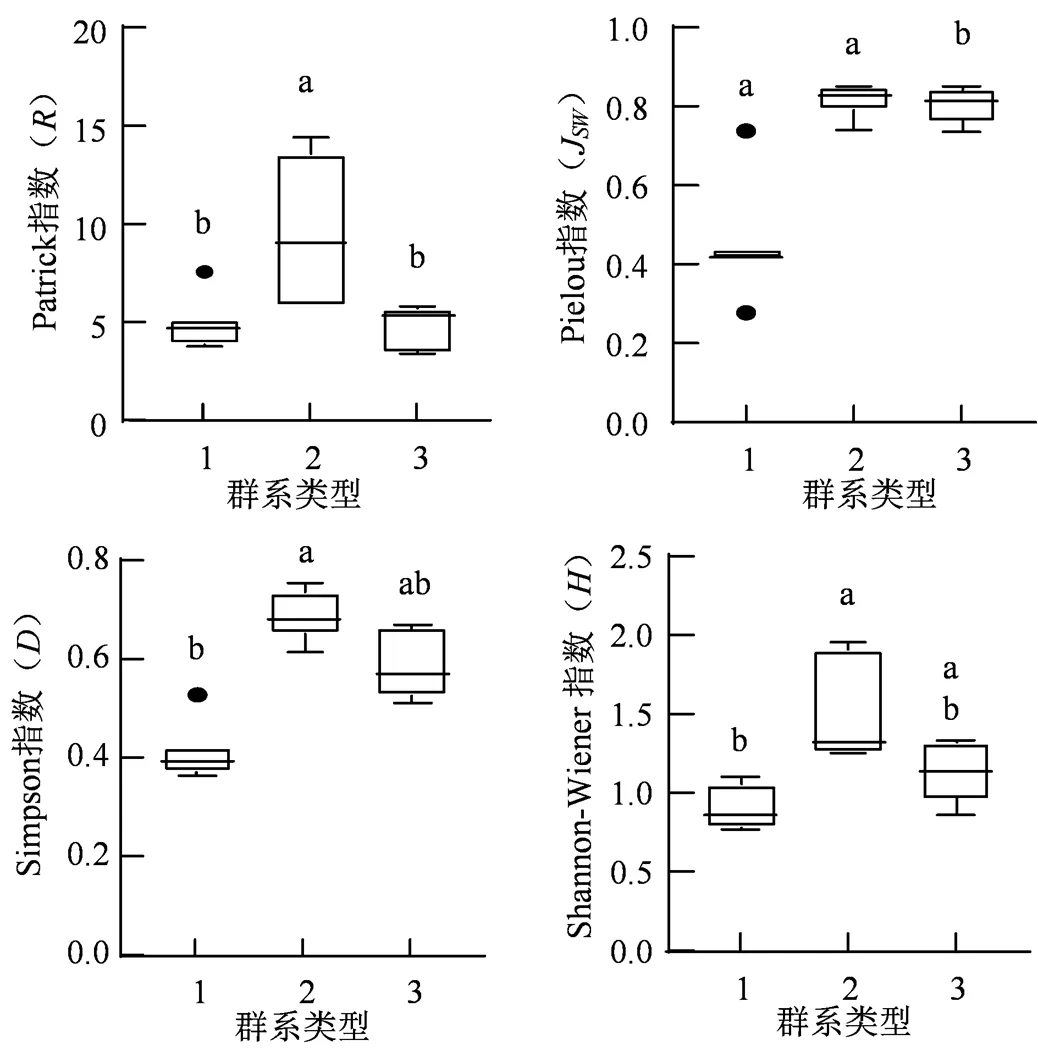
1. 表示黑松群系;2. 表示黑松+台湾相思群系;3. 表示黑松+滨柃群系。
Figure 3 Plant diversity of different formations
3.4 黑松样方不同层片与不同群系多样性水平
为探究黑松样方中不同层片与不同群系的多样性水平,对黑松样方中乔灌草各层的多样性指数和黑松3个群系的多样性指数进行 Kruskal-Wallis检验和多重比较。样方各层片的多样性指数结果(图2)显示,乔木、灌木与草本层在、和指数均存在显著差异(< 0.05),灌木与草本层的多样性水平总体上大于乔木层,乔木、灌木和草本层在J指数上并无显著差异,说明各层片的不同物种数量之间较为均匀,不同层片的与指数主要受指数的物种数()影响。图3为黑松群系、黑松+台湾相思群系以及黑松+滨柃群系的多样性水平,3个群系在、J、和指数均存在显著差异(< 0.05),不同群系的多样性水平差异较大,可能是生境异质性引起的。黑松+台湾相思群系在、J、和指数均明显高于黑松群系与黑松+滨柃群系,黑松+滨柃群系的J、和指数在总体上高于黑松群系,但在丰富度指数上两者水平相近,这是由于黑松群系与黑松+滨柃群系、指数的差异主要受J指数的影响。
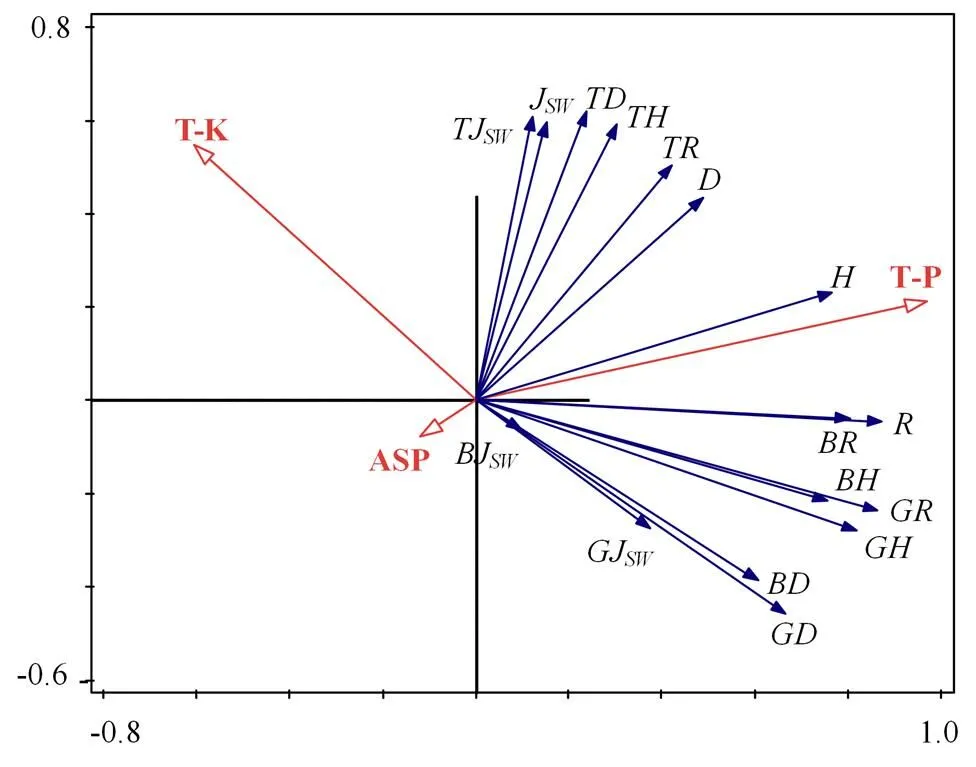
T-P:全磷含量;T-K:全钾含量;ASP:坡向;R:Patrick群落丰富度指数;JSW:Pielou群落均匀度指数;D:Simpson群落多样性指数;H:Shannon-wiener群落多样性指数;TR、TJSW、TD和TH分别代表乔木层Pielou、Pielou、Simpson和Shannon-wiener指数,同理灌木层与草本层的Pielou、Pielou、Simpson和Shannon-wiener多样性指数分别为BR、BJSW、BD、BH、GR、GJSW、GD和GH。
Figure 4 RDA ordination diagram of diversity indexes and environmental factors

图5 环境因子方差分解图
Figure 5 Variance decomposition diagram of environmental factors
3.5 黑松多样性与环境因子的关系
经Canoco5软件分析,第一主轴长度(0.5SD)小于3SD,表明黑松群落多样性指数与环境因子为线性关系。将群落、各层片的多样性指数与环境因子进行冗余分析并通过向前筛选法与蒙特卡洛置换检验,结果(图4)显示T-P(全磷)、T-K(全钾)与ASP(坡向)对黑松群落多样性影响最大(< 0.05)。经方差分解分析(图5),三者的总解释量为42.3%,T-P单独解释量为23.9%,T-K单独解释量为16.1%,ASP单独解释量为6.7%,三者共同解释量为0.2%,存在57.7%未知因素解释量。T-P与乔灌草各层及群落的J和多样性指数与呈正相关。T-K与群落的J均匀度指数及乔木层的J和多样性指数呈正相关,与灌木和草本层的、J、和多样性指数以及群落的、和多样性指数呈负相关。ASP与乔灌草层及群落的、J、和多样性指数呈负相关。群落多样性指数与主要环境因子(表4)显示群落的丰富度指数在3.4 ~ 14.4之间,群落的J均匀度指数在0.28 ~ 0.85之间,群落的多样性指数在0.37 ~ 0.75之间,群落的指数在0.77 ~ 1. 97之间,不同样方之间多样性指数差异较大。

表4 多样性指数及主要环境因子
4 讨论
4.1 黑松群落的组成与特征
样方中共包括54科117属的138种植物,其中裸子植物(1种)与蕨类植物(6种)种类较少。郑凤英等[27]调查发现威海市黑松群落也仅有2种裸子植物与1种蕨类植物,张俊艳等[28]认为针叶林相比阔叶林光照条件好,这导致黑松群落的生境较为干热,不利于蕨类植物生长,而调查结果发现阳性植物占比最大(61.59%)也能从侧面印证这一点。从生长型看,草本(55.07%)与灌木(28.26%)种类最多,乔木(5.80%)种类最少,与浙江无居民海岛[29]中草本(62.91%)和灌木(29.58%)占比相近。过去平潭地区植被遭到严重破坏,原有乔木数量少,后期人工引种的黑松、台湾相思等防护植物占据了较大优势,影响了其他乔木种类生长,而草本、灌木植物凭借繁殖快速以及对恶劣环境具有较好抗性,能快速抢占林下生存空间,因此两者占比较乔木高。从生活型看,黑松群落中高位芽植物(45.65%)与一年生植物(21.74%)占比高,高位芽植物中矮高位芽(45.65%)与小型高位芽植物(3.62%)最多,这与张坚强等[30]调查结果相似,淇澳岛高位芽植物中矮高位芽植物(23.26%)和小型高位芽植物(33.72)数量也是最多。他认为这是由于人工引进植物进行群落构建而影响了群落中植物生活型分布;另外平潭地区旱雨季节分明,降水量小于蒸发量,这在一定程度上也限制了大型高位芽植物的生长。从地理区系成分看,科的热带成分占比85.71%,属的热带成分占比66.67%,科的R/T值为6.0,属的R/T值为2.0,相比外马廊山岛[31]的热带成分科(71.40%)和属(49.00%)更高,这主要是由于纬度引起的,平潭地区的纬度更低,科属热带性质更强。黑松群落中泛热带分布科(64.29%)与泛热带分布属(35.35%)最多,其原因是平潭所处位置为中亚与南亚热带过渡区,属于泛热带分布区域,生长的植物符合该气候类型。黑松群落中优势科与优势属不明显,存在大量科内只包含一属以及属内只包含一种的现象,从侧面反应海岛植物种类分布更偏重生境条件,以能适应恶劣气候、抗性强的科、属分布较多。
4.2 多样性及其与环境因子相关性
不同层片在、和指数均存在显著差异,草本和灌木层多样性指数总体高于乔木层,这种差异与不同层片的物种数量()有关,而不同层片的物种数量则与前文提到的人为影响、不同生长型的抗性及生长繁殖策略是相关的。黑松群系、黑松+台湾相思群系在多样性水平具有显著差异(< 0.05)。从图3的箱线图来看,黑松+台湾相思群系的、J、和多样性指数相较黑松群系更高。郑凤英等[14]的研究也发现,黑松混交林在丰富度与多样性指数上要高于黑松纯林。有学者认为阔叶林多样性高于针叶林,针阔混交林大于纯林[32],不同植物在营养需求、根系特点、生长快慢等方面存在差异,混交林较纯林具有更高的养分利用率[33];梁文斌等[34]认为阔叶林的凋落物分解速度较针叶林快,对土壤有改善作用,有利植物生长发育。通过对黑松群系与黑松+台湾相思群系的土壤成分进行Kruskal-Wallis检验及多重比较发现,黑松+台湾相思群系土壤中的全磷含量(< 0.05)显著高于黑松群系,推测黑松+台湾相思群系相较黑松群系对土壤改善作用好,生境条件更加优越,因而具有更高的多样性水平。
从多样性与环境因子排序结果看,土壤全磷(T-P)、全钾含量(T-K)和坡向(ASP)对于黑松群落总解释量为42.3%,全磷为主要影响因子,单独解释量为23.9%;其次是全钾,单独解释量为16.1%,坡向单独解释量为6.7%。全磷与群落及各层多样性指数呈正相关。徐明锋等[35]发现全磷与多样性指数之间存在显著正相关性,在磷含量满足植物所需临界点前,磷含量越高,对植物生长发育越好。平潭地区土壤磷含量较为缺乏[36],样方中全磷含量均值(0.16 g·kg-1)要远小于全国土壤表层全磷均值(0.78 g·kg-1)[37],而磷对植物生长发育具有重要作用,海岛土壤多为肥力低的沙壤土,加上高温雨淋的气候条件,磷元素容易流失,植物生长所需的磷元素成为了限制性因素,从而影响多样性水平。全钾与乔木J、和多样性指数呈正相关而与灌草层多样性呈负相关,全钾并不能为植物所吸收,这一点还有待进一步研究。坡向与乔灌草及群落多样性指数呈负相关,如邹文涛等[38]认为阳坡多样性要小于阴坡,阳坡温度高、湿度低、蒸发量大,平潭地区本身相较大陆同纬度地区而言降水少、蒸发量大,土壤含水量与空气湿度的减少导致阳坡的植物多样性低于阴坡。环境因子对黑松群落的多样性的有效解释量只有42.3%,还有57.7%未知因素解释量,这可能与气候因子以及人为干扰因素有关。目前研究认为大尺度环境下,气候是影响群落分布的主要因素,中小尺度下,地形与土壤为主导因素[39],但平潭地区大风天数多、风力大,植物受风害较严重,气候对平潭地区植物群落可能具有较大影响。
5 结论
综上所述,平潭地区的黑松群落物种种类单一,缺少蕨类植物,乔木种类少,群落内部以阳性植物为主,区系成分的热带性质明显。物种多样性中,纯黑松林的多样性较混交林低,全磷、全钾与物种多样性呈正相关,坡向与其呈负相关,其余未知解释量可能与气候、人为干扰因子有关,未来需要对其进一步研究。在构建海岛黑松群落时需注意树种配置与地区微气候,尽量混交,避免单一树种成林,根据树种习性进行种植,同时需要加强磷肥使用,以提高海岛地区黑松群落多样性水平。
[1] 茹文明, 张金屯, 张峰, 等. 历山森林群落物种多样性与群落结构研究[J]. 应用生态学报, 2006, 17(4): 4561-4566.
[2] 汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述[J]. 生态学杂志, 2001, 20(4): 55-60.
[3] VALLI A T, KOUGIOUMOUTZIS K, ILIADOU E, et al. Determinants of alpha and beta vascular plant diversity in Mediterranean Island systems: the Ionian Islands, Greece[J]. Nord J Bot, 2019, 37(1): e02156.
[4] 魏娜, 王中生, 冷欣, 等. 海洋岛屿生物多样性保育研究进展[J]. 生态学杂志, 2008, 27(3): 460-468.
[5] 张巧, 黄义雄, 文华英, 等. 福建平潭岛沿海防护林综合生态效益评价[J]. 西南林业大学学报, 2015, 35(2): 63-67.
[6] 周永学, 樊军锋, 曹锋. 基质对奥地利黑松容器苗生长的影响[J]. 西部林业科学, 2017, 46(1): 130-133.
[7] MARUYAMA E, HOSOI Y, ISHII K. Somatic embryo production and plant regeneration of Japanese black pine ()[J]. J For Res, 2005, 10(5): 403-407.
[8] FENG Y Q, RUI L, WANG X Y, et al. Adaptation of pine wood nematode,, early in its interaction with twospecies that differ in resistance[J]. J For Res, 2022, 33(4): 1391-1400.
[9] 张治宇, 林茂松, 余本渊. 拟松材线虫对黑松苗的致病性[J]. 南京农业大学学报, 2004, 27(1): 46-50.
[10] 温祝桂, 朱小梅, 刘冲, 等. 两株外生菌根真菌对盐渍土壤中黑松幼苗生长的影响[J]. 中南林业科技大学学报, 2019, 39(4): 22-27.
[11] 吴小芹, 郑玲, 叶建仁. 黑松三种菌根苗根系构型差异及其与生长的关系[J]. 生态学报, 2009, 29(10): 5493-5499.
[12] HONG Y, EUIJOO K, LEE E, et al. Characteristics of vegetation succession on theforests in warm temperate regions, Jeju Island, South Korea[J]. J Ecol Environ, 2019, 43(1): 1-16.
[13] 石娇星, 许洺山, 方晓晨, 等. 中国东部海岛黑松群落功能多样性的纬度变异及其影响因素[J]. 植物生态学报, 2021, 45(2): 163-173.
[14] 郑凤英, 杜伟, 苟学文. 威海市区黑松林群落的物种多样性特征[J]. 生态环境, 2008, 17(5): 1965-1969.
[15] 侯艳茹. 平潭气象水文要素变化趋势分析[J]. 水利科技, 2019(3): 18-20, 23.
[16] 郑建忠. 平潭6个海岛植物群落特征及相关环境因子研究[D]. 福州: 福建农林大学, 2019.
[17] 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000.
[18] 中国科学院中国植物志编辑委员会. 中国植物志-第一卷-总论[M]. 北京: 科学出版社, 2004.
[19] 福建省科学技术委员会. 福建植物志[M]. 福州: 福建科学技术出版社, 1982.
[20] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245-257.
[21] RAUNKIAER C, GILBERT-CARTER H, FAUSBØLL A, et al. The life forms of plants and statistical plant geography[M]. Oxford: The Clarendon Press, 1934.
[22] 宋永昌. 植被生态学[M]. 2版. 北京: 高等教育出版社, 2017.
[23] 张金屯. 数量生态学[M]. 3版. 北京: 科学出版社, 2018.
[24] 郑心炫, 张增可, 林华贞, 等. 平潭岛不同演替阶段植被群落物种多样性特征[J]. 林业资源管理, 2019(3): 66-73.
[25] 袁丛军, 刘娜, 谢涛, 等. 贵州青钱柳群落结构及物种多样性特征[J]. 江苏农业科学, 2022, 50(4): 131-138.
[26] 齐婷婷. 庙岛群岛森林植物群落结构及其土壤固碳的研究[D]. 天津: 天津理工大学, 2015.
[27] 郑凤英, 王法明. 威海市区黑松林群落分析的研究[J]. 生态环境, 2006, 15(4): 787-790.
[28] 张俊艳. 海南岛热带天然针叶林—阔叶林交错区的群落特征研究[D]. 北京: 中国林业科学研究院, 2014.
[29] 张凯迪, 魏艳艳, 龚元, 等. 浙江沿海无居民海岛植物群落物种组成及多样性[J]. 地球环境学报, 2019, 10(1): 58-68.
[30] 张坚强, 张琳婷, 赵东铭, 等. 珠海淇澳岛次生植被特征及物种多样性[J]. 西北植物学报, 2019, 39(1): 173-184.
[31] 郑俊鸣, 方笑, 朱雪平, 等. 外马廊山岛植被特性与植物多样性[J]. 广西植物, 2017, 37(3): 271-279.
[32] 邵水仙, 李红丽, 董智, 等. 退化砂石山地人工林林下植物群落特征与物种多样性[J]. 水土保持研究, 2015, 22(5): 146-151, 157.
[33] HARTLEY M J. Rationale and methods for conserving biodiversity in plantation forests[J]. For Ecol Manag, 2002, 155(1/2/3): 81-95.
[34] 梁文斌, 刘文胜, 张合平, 等. 青羊湖林场不同林分物种多样性与水源涵养能力的研究[J]. 西北植物学报, 2015, 35(9): 1875-1881.
[35] 徐明锋, 胡砚秋, 李文斌, 等. 土壤养分对亚热带天然林物种分布的影响[J]. 中南林业科技大学学报, 2014, 34(9): 91-97.
[36] 张巧. 平潭岛四种主要沿海防护林生态效益评价[D]. 福州: 福建师范大学, 2015.
[37] TIAN H Q, CHEN G S, ZHANG C, et al. Pattern and variation of C:N:P ratios in China's soils: a synthesis of observational data[J]. Biogeochemistry, 2010, 98(1): 139-151.
[38] 邹文涛, 姜艳, 尹光天, 等. 石门森林公园不同海拔或坡向林地物种多样性的比较[J]. 中南林业科技大学学报, 2014, 34(4): 77-81.
[39] 张鸽, 王琦枝, 何媛, 等. 北川滑坡迹地植物群落多样性变化及环境解释[J]. 浙江农林大学学报, 2022, 39(1): 50-59.
Characteristic and species diversity ofcommunities in Pingtan island
XIONG Kuanhong1, XIAO Lan2, LIU Zilin1, HE Yaqin3, WEI Kai1, ZHENG Jianzhong4, DENG Chuanyuan1
(1. College of Landscape Architecture, Fujian Agriculture and Forestry University, Fuzhou 350002; 2. Island Research Center of Ministry of Natural Resources, Pingtan 350400; 3. College of Forestry, Guangxi University, Nanning 530004; 4. Department of Landscape Architecture, Fujian Forestry Vocational Technical College, Nanping 353000)
To explore the community characteristics and diversity ofcommunity, scientifically manage.shelterbelt and protect the diversity of.community, we investigated the species composition, flora, and community type of the.community with the quadrat method in eight affiliated islands of Pingtan from 2018 to 2020, and analyzed the relationship between the diversity indexes and environmental factors by redundancy analysis. The results showed that: (1) There were 138 species of vascular plants belonging to 117 genera and 54 families in the study area, with few gymnosperms and ferns in total. (2) Phanerophytes (68 species) and therophytes (30 species) were high in the life form. In terms of growth form, there were 8 species of arbors, 39 species of shrubs, 76 species of herbs, and 15 species of vines, with more species of herbs and shrubs than arbors. (3) Both families and genera had the characteristics of tropical traits but lack Chinese endemic distribution. (4) Group average clustering method was used to divide the vegetation in the research area, which consisted of three formation types and nine association types. (5) In the comparison of the overall diversity of the community,+plant formation was higher than the+plant formation andplant formation. The diversity indexes in the herb layer and shrub layer of each plant community were all higher than that in the tree layer. (6) The redundancy analysis showed that the soil total phosphorus, soil total potassium and slope aspect had significant effects on the diversity of.communities, with a total explanation of 42.3%. In conclusion, when building an island plant community dominated by, it is necessary to pay attention to the use of mixed and phosphate fertilizers and to allocate reasonably according to the regional microclimate conditions.
; community characteristic; species diversity; redundancy analysis
S718.54; S791.256
A
1672-352X (2023)05-0764-08
10.13610/j.cnki.1672-352x.20231030.006
2023-10-31 09:56:24
[URL] https://link.cnki.net/urlid/34.1162.S.20231030.1112.046
2022-09-24
福建省区域发展科技项目计划(2018Y3006),福建农林大学科技创新专项基金(CXZX2019086)和 2019年度福建省科技特派员工作经费项目(KTP19186A)共同资助。
熊宽洪,硕士研究生。E-mail:xiongkuanhong@qq.com
邓传远,博士,副教授。E-mail:dengchuanyuan@fafu.edu.cn
猜你喜欢
安徽农业科学(2023年2期)2023-02-25 04:02:08
美食(2022年12期)2022-12-09 02:08:48
现代农村科技(2022年3期)2022-03-14 06:09:04
海峡姐妹(2020年7期)2020-08-13 07:49:14
美食(2019年7期)2019-09-10 07:22:44
走向世界(2019年9期)2019-06-11 01:47:45
现代园艺(2018年11期)2018-06-15 10:46:20
海峡姐妹(2018年3期)2018-05-09 08:20:49
科学大众·小诺贝尔(2018年12期)2018-02-22 12:05:38
海峡姐妹(2017年7期)2017-07-31 19:08:20
