
云南水稻新品系重要农艺性状全基因组关联分析
2023-11-22 04:20:42杨丹丹李小林徐雨然奎丽梅
西南农业学报 2023年9期
杨丹丹,李小林,吕 莹,邓 伟,徐雨然,奎丽梅,涂 建,谭 静
(1.云南大学资源植物研究院,昆明 650504;2.云南省农业科学院粮食作物研究所,昆明 650205;3.云南省主要农作物种子加工工程技术研究中心,昆明 650205;4.云南省稻作遗传改良重点实验室,昆明 650205)
【研究意义】水稻是重要的粮食作物,全世界一半以上的人口以之为主食[1],中国是人口大国,水稻生产对中国粮食安全具有十分重要的战略意义。随着人口增长,进一步提高水稻单产十分重要。有效穗数、每穗粒数、千粒重是构成水稻产量的3个重要因素,剑叶长、剑叶宽、株高、穗长和结实率等重要农艺性状对水稻产量的形成也具有重要作用。这些重要农艺性状大部分都是受多个基因所控制的数量性状[2],通常会被环境所影响。基于SNP的全基因组关联分析已成为研究数量性状的主要方法,通过发掘水稻重要农艺性状的QTL位点及其关联标记,并筛选携带优良农艺性状的育种材料,是提高云南水稻产量最经济有效的方法。【前人研究进展】识别重要农艺性状基因位点是进行水稻遗传育种改良的重要手段,对于新育成的品系,要充分了解其农艺性状与基因位点的关联情况以便更好地开展育种工作。水稻的许多重要农艺性状已被定位,鲁清[3]利用5291个SNP标记,对523份稻种资源进行遗传结构和重要农艺性状分析,检测到6个与株型相关性状显著性位点、6个种子颜色相关性状显著性位点、2个直播出苗相关性状显著性位点、127个稻瘟病显著性位点。Huang等[4]以517份水稻为试验材料,选取14个农艺性状进行表型调查,先通过重测序对参试材料进行基因组高密度单倍型图谱构建,在此基础上再对参试材料进行全基因组关联分析,共筛选到80个显著关联位点。Begum等[5]选用363份热带优质水稻群体进行GBS测序,利用测序得到的71 710个SNPs对产量及产量构成性状进行全基因组关联分析,结果筛选到52个相关的QTLs。Zhou等[6]以超级杂交稻协优9308为供体材料创制的138份水稻新品系为试验材料,经过重测序获得79万个SNPs,结合全基因关联分析筛选出31个有关株高以及抽穗期的QTLs,其研究结果为进一步利用138份水稻新品系奠定了一定理论基础。【本研究切入点】全基因组关联分析(Genome-wide association study, GWAS)以连锁不平衡为基础,借助上万甚至百万个SNP标记,鉴定群体内目标性状和遗传变异之间的相关性,现在已广泛应用于水稻[4]、玉米[7]、大豆[8]、花生[9]、小麦[10]等农作物。本研究选用云南省农业科学院育成的124份水稻新品系为试验材料,对其剑叶长、剑叶宽、株高、有效穗数、穗长、每穗粒数、结实率、千粒重8个重要农艺性状进行GWAS分析。【拟解决的关键问题】以期鉴定出一批具有优良性状的水稻育种材料,并筛选出具有控制重要农艺性状的QTL位点,研究结果可为今后云南省水稻遗传改良、育种利用和分子标记辅助选择提供有用信息。
1 材料与方法
1.1 试验材料
试验材料为云南省农科院粮食作物研究所水稻种业科技创新团队选育的F8代水稻新品系124份,其中籼稻有82份,粳稻有42份(表1),该群体来源广泛且遗传背景较为丰富,在一定程度上能反映云南水稻种质资源情况。

表1 供试水稻材料
1.2 田间试验
将新育成的124份水稻品系于2020年3月在云南省农业科学院富民试验基地进行播种,4月单株移栽,试验设置3次重复,每个重复种3行,每行种10株材料,株行距为15 cm × 30 cm。
1.3 表型调查
待水稻成熟后,在田间每小区选择3株长势一致的单株,测量其剑叶长、剑叶宽和株高;收割材料进行自然风干,将材料晾干后,再考察每份材料的各个单株的有效穗数、穗长、每穗粒数、结实率和千粒重。
1.4 数据统计与分析
1.4.1 表型数据分析 用软件Excel 2017对获得的表型数据进行初步整理,再用软件SPSS statistic对各个性状进行方差分析和基本的描述性统计分析,统计参试材料8个农艺性状的最小值、最大值、平均值、标准差、变异系数、偏度和峰度,并绘制各个农艺性状的频数分布直方图。
1.4.2 SNP基因型分析 研究所使用的SNP标记来自于华智生物技术有限公司的SNP 56K芯片,将测序数据与参考基因组日本晴IRGSP1.0[11]进行比对并基于严格的质量控制,剔除基因型频率<5%的等位基因标记、缺失标记数>10%的标记和相关性>0.95的标记,筛选出高质量SNP标记进行分析,共筛选到26 707个SNP用于后续分析[12]。
1.4.3 聚类分析 利用软件Tassel V5.0的Cladogram模块对124份水稻的基因型数据进行Neighbor-Joining聚类分析[13]。
1.4.4 全基因组关联分析 利用软件Tassel V5.0的混合线性模型,结合群体SNP基因型数据,对水稻的剑叶长、剑叶宽、株高、有效穗数、穗长、每穗粒数、结实率、千粒重用MLM模型进行全基因组关联分析,本研究以-log10(P)>3为筛选阈值,P=1.0×10-5作为显著标记选取的标准[14]。用R2评价关联位点对表型性状的总体贡献率,鉴定出的QTL并采用CGSNL命名法命名[15]。
2 结果与分析
2.1 124个新品系8个主要农艺性状变异性分析
从表2可以看出,参试群体呈现出极广泛的表型变异,124份水稻各性状均存在极显著差异。参试材料的8个农艺性状变异系数均在10%以上,表明该8个农艺性状在群体中的离散程度较大,其中变异系数最大的是有效穗数,为29%;变异程度较高;穗长和千粒重变异系数较小,分别为0.12、0.14。从图1可以看出,参试材料表型值的偏度与峰度绝对值系数大多小于1,呈现典型数量性状特点,可用于后续进行关联分析。对比《水稻新品种测试原理与方法》标准,共68份水稻的农艺性状具有较高值,其中有10份水稻的剑叶长超过45.1 cm,12份水稻的剑叶宽超过2.6 cm,有13份水稻的株高超过121 cm;参试水稻结实率较高,每穗粒数在250粒以上的有33份,小穗结实率在60%以上的有84份,占参试材料的68%;小穗结实率在81%以上的有21份;千粒重超过30 g的有8份,其中既是大穗又是大粒的有2份。

图1 参试材料农艺性状表型值频数分布Fig.1 The frequency distribution of agronomic traits phenotypic values of test materials
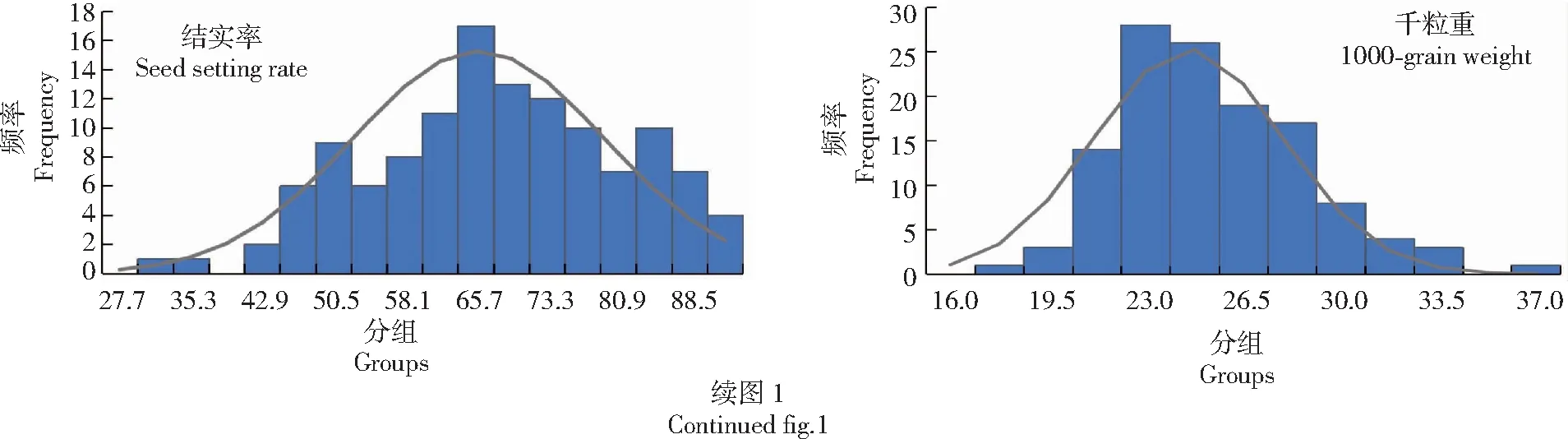

表2 124份水稻新品系重要农艺性状的表型鉴定
2.2 124份新品系SNP及聚类分析
通过对124份水稻新品系8个重要农艺性状进行全基因组关联分析,共检测到204个SNP位点(P≤0.001),分布于水稻的12条染色体上,阈值在P=1.0×10-5的位点有39个,对39个SNP标记进行QTL分析。依据124份水稻材料的基因型数据计算遗传距离并进行聚类分析,如图2所示,水稻群体分化十分明显,124份水稻被分成群体I、群体II、群体III,分别包括18、34和72份材料。通过对材料系谱信息和已知品种的籼粳亚种属性分析,籼稻品系主要聚集在群体I和群体II中,群体III是籼稻与粳稻的混合群体,其中粳稻占主要部分。

图2 124份材料聚类分析Fig.2 Cluster analysis of 124 materials
2.3 全基因组关联分析
从表3可见,检测到与剑叶长显著相关的QTL有5个,分别是位于1号染色体上的qFLL1,11号染色体上的qFLL11.1、qFLL11.2、qFLL11.3、qFLL11.4,表型解释率为9.5%~17.35%;与剑叶宽显著相关的QTL位点有1个,是位于1号染色体上的qFLW1,表型解释率为14.00%;与株高相关的显著位点4个,分别是位于2号染色体上的qPH2.1、qPH2.2,6号染色体上的qPH6.1、qPH6.2,表型解释率为12.01%~13.82%;与有效穗数相关的显著位点3个,是位于9号染色体上的qEPN9.1、qEPN9.2、qEPN9.3,表型解释率为11.49%~14.85%;与穗长相关的显著位点4个,分别是位于2号染色体上的qPL2,3号染色体上的qPL3,11号染色体上的qPL11,12号染色体上的qPL12,表型解释率为10.03%~13.77%;与每穗粒数相关显著位点6个,分别是位于2号染色体上的qGNPP2.1、qGNPP2.2,12号染色体上的qGNPP12.1、qGNPP12.2、qGNPP12.3、qGNPP12.4,表型解释率为9.9%~15.33%;与结实率相关的显著位点2个,是位于1号染色体上的qSSR1.1、qSSR1.2,表型解释率为9.8%~11.7%;筛选到14个位点与千粒重相关联,分别是位于3号染色体上的qTGW3.1、qTGW3.2、qTGW3.3、qTGW3.4,位于4号染色体上的qTGW4.1、qTGW4.2,位于6号染色体上的qTGW6,位于7号染色体上的qTGW7,位于8号染色体上的qTGW8.1、qTGW8.2、qTGW8.3、qTGW8.4,位于10号染色体上的qTGW10,位于11号染色体上的qTGW11,表型解释率为13.52%~19.50%。

表3 水稻主要农艺性状全基因组关联分析检测到的位点
本研究共鉴定出8份千粒重超过30 g的材料,通过对这8份材料千粒重的位点进行分析,从表4可见,在8号染色体的qTGW8.2、qTGW8.3、qTGW8.4 3个位点上有7份大粒材料集中出现,分别是材料D326、D543、D546、D553、D556、D558、D1059;在11号染色体的qTGW11位点上集中出现5份材料,分别为材料D45、D326、D543、D546、D553;在其他位点上均没有筛选到相关材料。

表4 8份材料千粒重位点分析
3 讨 论
3.1 关联分析群体表型性状研究
水稻是云南省重要的粮食作物,是农民主要经济收入来源之一。在现代水稻育种策略中,大穗、大粒、矮秆的理想株型育种是提高水稻产量的重要方法[16-17],叶片是水稻进行光合作用的最主要器官,剑叶长宽能通过影响水稻光合效率,从而影响水稻产量[18],鉴定出的D45、D326、D556、D558、D1125等优势材料是提高云南水稻产量的重要资源。前人在对株高的研究中,主要对矮秆、半矮秆基因研究较多,对高秆基因的研究较少[19]。试验鉴定出高秆材料13份(株高>121 cm),可以在后续试验定位材料中与株高相关的QTL位点,通过鉴定出的QTL改良材料株型,以此来更好地利用水稻材料资源。
3.2 关联分析群体的群体结构
聚类分析将124份材料分成3个群体,经过对亲本品种的籼粳亚种属性分析可知,群体I和群体II为籼稻群体,群体III为粳稻/部分籼稻品系,并且亲缘关系近的材料聚集的位置较近,分群结果与材料类型基本一致。但在分群时会出现亲本组合完全一样的2个品系被分到2个群体中,如D820和D824这2个材料亲本组合都为云恢290×辐恢669,但D820被聚在群体II中,D824被聚在群体I中,可能因为材料在长期选育过程中由于品系间发生基因交流或者基因重组引起。简单重复序列SSR分子标记技术因具多态性好、共显性、技术简便等优点被广泛应用于植物遗传多样性的研究[20-21]。在今后的研究中,可以考虑进行多年多点试验,结合SSR分子标记技术进行鉴定。
3.3 水稻农艺性状的GWAS研究
本研究表明,剑叶是影响水稻产量的重要因素,在1、11号染色体上共检测到6个与剑叶相关的QTL位点,其中qFLW1与鲁清[3]定位到的QTL较相近,其余的QTL位点未见报道,需进行进一步验证。
Gramene数据库中已报道1011个关于株高的QTL[22],许多基因已被克隆,如半矮秆基因sd1[23],矮秆基因d35[24],OsPNH1[25],本试验在2、6号染色体上定位4个QTL,其中位于6号染上体上的qPH6.1、qPH6.2与基因OsPNH1较接近;位于2号染色体上的2个QTL是需进一步验证的新位点。
水稻穗部性状主要包括穗长、穗粒数、有效穗数等,与产量密切相关,在Gramene数据库中已报道了253个关于穗长的QTL,353个关于每穗粒数的QTL,213个关于有效穗数的QTL[26];Li等[27]克隆了一个水稻短穗突变体基因SP1。Li等[28]、Zhang等[29]克隆了长穗基因LP、qPL6。穗粒数基因Gn1a、gnp4、OSEB等[30-31]、一些控制分蘖的基因(如MOC1[32]、D3[33]等)已被克隆。本研究定位到与穗长相关位点4个,与每穗粒数相关位点6个,与有效穗数相关位点3个均未见报道,但穗部性状大多是数量性状,容易受环境影响,本研究定位到的位点需进一步进行遗传验证。
本研究筛选到与千粒重相关的显著位点14个,与结实率相关位点2个,分别位于1、3、4、6、7、8、10、11号染色体上。其中位于3号染色体上的4个位点与魏秀彩[34]的研究结果相近,7号染色体上的位点与魏秀彩等[35]研究一致。其余位点未见报道,需进行进一步验证。
本研究通过对124份参试材料表型进行鉴定,检测到10份长剑叶材料、12份宽剑叶材料、13份高杆材料、21份高结实率材料、8份大粒材料、33份大穗材料,但除了千粒重以外,其他性状位点遗传较复杂。通过对8份材料的千粒重位点进行分析,有7份大粒材料在8号染色体的3个位点上集中出现,除D326为杂合外,其余6份材料都为纯合,并且这3个位点的距离较近,推测这3个SNP位点是属于控制同一个基因的主效QTL新位点;在11号染色体上的位点上也出现了5份大粒材料,这个位点可能是控制千粒重的非主效基因QTL位点,具体遗传效应需进行进一步验证。
4 结 论
选用云南省农业科学院育成的124份水稻新品系为试验材料,对其剑叶长、剑叶宽、株高、有效穗数、穗长、每穗粒数、结实率、千粒重8个重要农艺性状进行测定,通过表型共鉴定出33份大穗材料、8份大粒材料;通过全基因组关联分析共筛选到204个相关位点(P≤0.001),其中显著位点39个。
猜你喜欢
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
农技服务(2023年2期)2023-03-15 00:43:08
中国水稻科学(2021年6期)2021-11-18 12:06:30
落叶果树(2021年6期)2021-02-12 01:29:20
天津农林科技(2020年3期)2020-08-13 03:16:56
园林科技(2020年2期)2020-01-18 03:28:18
浙江农业科学(2019年1期)2019-01-25 05:25:34
中国蔬菜(2016年8期)2017-01-15 14:23:37
扬子江(2016年1期)2016-05-19 22:06:26
中国棉花加工(2015年4期)2015-12-19 06:40:58
