
不同类型抗菌肽的结构及作用机制
2023-11-20 12:50宋雪莹
当代畜禽养殖业 2023年5期
宋雪莹
(黑龙江省农业科学院畜牧兽医分院,黑龙江齐齐哈尔 160000)
抗菌肽(antimicrobial peptides, AMPs)也称为宿主防御肽,是生物机体对抗入侵微生物过程中形成的防御系统。 AMPs 大小在40 个氨基酸残基以内,通常呈阳离子[1]。 AMPs 作为先天免疫系统的一部分,对革兰阴性和革兰阳性细菌都表现出广谱抗菌活性。 迄今为止,已确定AMPs 发挥抗菌活性的主要作用机制包括破坏膜完整性和抑制细胞内活性。 不同类别AMPs都有独特的结构和微生物细胞内活性抑制机制,渗透到脂质双分子层[2-3],通过不同机制杀灭细菌。 例如,影响转录和DNA 复制、细胞质分裂、细胞壁生物合成以及抑制酶活性和蛋白质合成[4]。
1 牛乳铁蛋白肽(LfcinB)
LfcinB 是由乳铁蛋白N-末端结构域 (17~41 个残基)酸水解产生的一种阳离子抗菌肽,具有抗细菌、抗真菌和抗病毒活性。 LfcinB 含有25 个残基的多肽片段,包括α 螺旋结构[5]、反向平行β 结构[6]和由2 个半胱氨酸残基之间的二硫键形成的环状结构。 LfcinB具有两亲性,因为肽的一个表面呈疏水性;另一表面呈亲水性。 LfcinB 破坏细菌质膜的能力被认为是影响抗菌活性的主要因素,膜电位在病菌质膜局部脂质双分子层破裂和快速渗透中起重要作用[7]。 LfcinB(4~9)(RRWQWR) 残基被认为是乳铁素B 的抗菌活性中心,当C 端的羧基被酰胺化时,显示出与LfcinB 相当的抗菌活性[8]。
LfcinB(21~25)Pal(RWQWRWQWR)的净电荷为4+,具有两亲性。 肽有一个由精氨酸残基形成的带电面;另一侧含有谷氨酰胺残基,这两侧都含有2 个色氨酸残基。 对大肠杆菌ATCC 25922、 粪肠杆菌ATCC 29212、 肠炎菌ATCC 13076、 铜绿假单胞菌ATCC 27853 和金黄色葡萄球菌ATCC 25923 具有抑菌活性[9]。
2 天蚕素(Cecropin)
Cecropin 是一种带正电荷的α 螺旋肽,具有广谱抗菌活性,有一定的抗病毒和抗真菌特性,由节肢动物先天免疫系统产生,可作为理想的天然抗生素替代品。 Cecropin 具有强碱性的N 端和高疏水的C 端,表现出两亲性。 根据氨基酸序列不同,分为天蚕素A、B、C 和D 等。 不同种类天蚕素的氨基酸序列见表1[10]。

表1 同种类天蚕素的氨基酸序列[10]
Cecropin A 是最早发现的天蚕素之一,是从蛾类Hyalophora cecropia 中分离出来的[11],由37 个氨基酸残基组成,呈α 螺旋结构[12]。 Cecropin A 通过快速渗透病原体外膜,致使其破裂死亡。
Cecropin B 是一种线性带正电荷多肽, 由35 个氨基酸组成。 目前研究表明,在天蚕素家族的所有成员中,Cecropin B 对革兰阴性菌具有较强的抗菌活性,并且对人体细胞具有较低的细胞毒性[13]。
Cecropin C 由37 个氨基酸组成,目前在抗细菌活性方面研究较少。 因其与Cecropin A 相比,缺少C 端阻断基团,被认为是Cecropin A 的前体或降解产物[14]。
Cecropin D 由19 个氨基酸残基组成, 末端缺乏大部分赖氨酸残基, 不带电荷。 Cecropin D 比Cecropin A、Cecropin B 疏水性更强。Cecropin D 对革兰阳性和革兰阴性细菌具有抗菌作用,同时也表现出强大的抗病毒特性[15]。
3 组织蛋白酶抑制素(Cathelicidin)
Cathelicidin 由哺乳动物固有宿主防御系统上皮细胞产生,主要包含一个非活性抗菌结构域和一个信号肽。 Cathelicidin 除具有抗菌作用外,还参与许多重要的生理现象, 如细胞凋亡、 免疫调节和伤口愈合。Cathelicidin 基因CAMP 编码18 kDa 前体, 人类阳离子抗菌蛋白hCAP18 经蛋白裂解得到成熟的生物活性肽LL-37[16]。LL-37 已在包括角质形成细胞、单核细胞、肥大细胞和中性粒细胞在内的许多细胞中被鉴定出来, 在先天免疫细胞反应中起着多方面的作用,包括诱导白细胞介素8(IL-8),上调趋化因子受体4(CXCR4)、趋化因子受体2(CCR2)和白介素8 受体(IL-8RB),并可作为中性粒细胞和单核细胞的趋化剂[17]。
4 作用机理
目前,抗菌肽的抗菌机制尚无明确结论,普遍认可以下4 种模型:凝集模型、桶壁模型、环面孔隙模型和地毯模型(图1)。
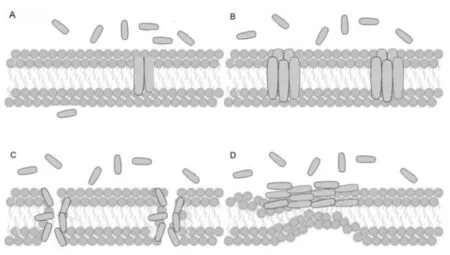
图1 肽的作用机理[22]
凝集模型:抗菌肽在静电作用下,与细菌细胞膜上带负电的磷脂酰肌醇相互作用, 凝集在细胞膜两侧,使得细胞膜去极化[18]。
桶壁模型:该模型主要假设抗菌肽是在“杆”中形成的,并且可以相互结合。 由于这种结合形成了桶状孔隙。 这些孔隙的形成是基于抗菌肽的残基与脂质双分子层脂肪酸链相互作用,并且亲水区域排列在形成的孔隙内部[19]。
环面孔隙模型:有些短链结构的抗菌肽,不能同α 螺旋结构一样在脂质双分子层中积累,因此提出了环面孔隙模型。 其中,环形孔被认为是由多肽和脂质组成的,因此该类抗菌肽分子不需要穿过细胞膜的两层,与地毯机制类似,聚集的多肽可能会影响膜的厚度,使其变薄并形成孔隙[20]。
地毯模型:该模型显示抗菌肽对细胞膜完整性的影响。 在细胞外的肽与细胞膜结合并覆盖其表面,以与脂质双分子层平行的方向积累,由此产生的抗菌肽聚合体斑块类似于地毯,因此而得名[21-22]。
5 结语
目前, 对抗菌肽抑菌活性研究较多。 通过测试LfcinB、Cecropin 和Cathelicidin 等对不同抗革兰阴性菌和革兰阳性菌的最小抑菌浓度(MIC),证明这些抗菌肽均具有较好的抑菌活性。 在畜禽生产中虽然也开展了一些应用研究,但针对不同畜禽品种、不同生长阶段的研究还不够深入。 因此,将不同来源抗菌肽应用在畜禽生产中应该是未来研究的方向之一。
猜你喜欢
生物化学与生物物理进展(2022年6期)2022-07-21
现代畜牧科技(2021年9期)2021-10-13
中国动物保健(2021年4期)2021-07-21
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
数学小灵通·3-4年级(2018年12期)2018-01-26
中国蚕业(2017年1期)2017-04-08
吉林农业(2016年23期)2016-12-22
广东饲料(2016年5期)2016-12-01
国外医药(抗生素分册)(2016年5期)2016-07-12
池州学院学报(2015年3期)2016-01-05
