
限制性内切酶Bsa I 的分离纯化与结晶及其硒代衍生物的制备
2023-11-20 07:45郑梦泽贾玮玮刘子昂张传志王伟玲李婷婷
食品工业科技 2023年22期
朱 海,郑梦泽,贾玮玮,周 倩,谈 帅,刘子昂,张传志,王伟玲,李婷婷,
(1.江苏海洋大学,江苏省海洋药物活性分子筛选重点实验室,江苏连云港 222005;2.江苏省海洋生物产业技术协同创新中心,江苏连云港 222005;3.正大天晴药业集团股份有限公司,江苏连云港 222005)
限制性内切酶是分子生物学中最常用的工具酶之一[1],其中,Ⅱ型限制性内切酶在识别序列的内部或附近确定位点对DNA 进行特异性切割,由于其稳定的裂解模式及简便的催化方式而被广泛应用于基因重组等分子生物学领域,具有重要的商业价值[2]。根据Ⅱ型限制性内切酶在识别序列上的不同,可以将其分为不同的亚类:例如,以EcoR I、Hind III 为代表的ⅡP 型内切酶,可以识别回文序列,并在识别序列内部进行对称切割;以Bsa I、Fok I 为代表的ⅡS 型,可以识别不对称序列,并在识别序列一端的固定位置进行切割[3-4]。
Bsa I 是一种来源于嗜热芽孢杆菌的ⅡS 型限制酶,可以特异性识别不对称DNA 序列5’-GGTCTC-3’,并在识别序列3’端第1 个碱基N1 后,以及识别的互补序列3’-CCAGAG-5’的5’端第5 个碱基N5后切割,从而留下4 个碱基的5’突出端[5-6]。常用于Golden Gate 组装(Golden Gate Assembly)以及mRNA疫苗用质粒的线性化或其它体外转录质粒的线性化[7-10]。Bsa I 作为基因敲除的工具,在揭示植物基因功能和作物改良方面具有举足轻重的作用[11-13]。天然来源的限制酶使用往往具有局限性,比如活性低,星号活性明显,使用条件苛刻等[14-15]。这些局限性极大地影响了内切酶在商业化中的应用,所以研究者对于天然限制酶的研究和改造一直都没有停止。为了克服天然酶存在的问题,针对Bsa I 进行结构研究具有重要意义。
X 射线晶体衍射是解析生物大分子三维结构的主要手段之一。其主要过程包括数据收集、相位解析、初始电子密度优化以及原子模型的构建[16-18]。对于生物大分子,一般情况下难以直接从衍射数据求得相位。结构解析中解决相位问题的一般方法有分子置换法,同晶置换法和反常散射法[19-20]。分子置换法需要同源性较高的同源结构来解析相位,而Ⅱ型限制酶同源性较低,无法利用分子置换法解析Bsa I 的相位问题。同晶置换法需要首先获得大量蛋白晶体进行重金属浸泡,因此本实验尝试在重组蛋白表达时制备硒代蛋白衍生物,利用反常散射法推算晶体相位[21-22]。
本研究以pBAD 质粒为载体,利用大肠杆菌ER2566(E.coli.)为宿主菌,制备ⅡS 型限制性内切酶Bsa I 及其硒代蛋白衍生物,利用坐滴法进行初步的晶体生长研究,为后续阐明Bsa I 的三维结构及作用机理提供实验基础,进而为其定向改造和功能优化提供信息。
1 材料与方法
1.1 材料与仪器
感受态菌株B834(DE3)、ER2566 武汉淼灵生物科技有限公司;λDNA 莫纳(武汉)生物科技有限公司;酵母提取物、蛋白胨 Thermo Fisher Scientific 公司;Ni 亲和层析填料、UniGel-80Q 阴离子层析填料 江苏纳微生物科技有限公司;其余化学试剂均为分析纯 国药集团化学试剂有限公司;基础培养基配方 LB 培养基(g/L):酵母提取物 5.0,胰蛋白胨10.0,氯化钠 10.0;M9 培养基(g/L):十二水合磷酸氢二钠 17.09,磷酸二氢钾 3.0,葡萄糖 4.0,硫酸镁0.24,氯化钠 0.5,氯化钙 0.011,氯化铵 1.0。
凝胶成像系统 上海勤翔科学仪器有限公司;超高压连续流细胞破碎仪 广州聚能生物科技有限公司;AKTATm Pure 蛋白纯化仪 GE 公司。
1.2 实验方法
1.2.1 重组菌株的获取 将编码Ⅱ型限制酶Bsa I序列(Q6SPF4)及甲基化酶 Bsa I-M 基因序列(Q6SPF6)经人工合成构建在pBAD 载体上(安徽通用生物公司)。将重组质粒通过热激法转入到ER2566感受态细胞内(冰浴30 min,42 ℃热激1 min,冰浴5 min,加入500 μL 无抗LB,37 ℃,200 r/min 培养1 h),将培养液在固体平板划线,37 ℃培养箱培养过夜,获得重组菌株[23]。
1.2.2 重组Bsa I 蛋白的表达与纯化 将重组菌株ER2566-Bsa I 在含有氨苄霉素的LB 平板上划线稀释,37 ℃培养箱培养过夜。挑取数个单克隆在含有氨苄霉素的LB 液体培养基中活化,37 ℃ 200 r/min摇床培养至OD600值为0.6~1.0。进行小量诱导实验(实验组1),取活化液10 mL,加入20% L-ara,37 ℃诱导5 h 后,收集菌体,加水溶解,100 ℃煮沸15 min,取上清,SDS-PAGE 点胶验证蛋白表达情况,通过实验组1 确定蛋白表达后进行大量诱导实验(实验组2)。按1:50 比例在500 mL LB 培养基中扩大培养,其OD600值为0.6~0.8 时加入20% L-ara,在37 ℃下进行诱导5 h 后,收集发酵菌体。发酵菌体重悬至裂解液(25 mmol/L Tris-HCl,pH8.0,150 mmol/L NaCl),用细胞破碎仪1000 bar 破碎至菌液澄清透亮,4 ℃下13000 r/min 离心1 h,收集上清;然后将上清液与镍填料结合过夜;利用15 mmol/L 低浓度咪唑洗杂蛋白,再利用250 mmol/L 高浓度咪唑洗脱目的蛋白;目的蛋白洗脱液用UniGel-80Q 阴离子层析柱进行下一步纯化,低盐溶液(25 mmol/L Tris-HCl,pH8.0)进行柱平衡,盐浓度梯度洗脱目的蛋白。蛋白浓度的确定:通过测量样品在280 nm 处的吸光度。
1.2.3 Se-Bsa I 蛋白的表达与纯化 活化方法见1.2.2,首先进行小量诱导实验(实验组1),取活化液200 μL 加入10 mL 过度培养基中(LB 培养基:M9培养基体积比为1:4),37 ℃ 200 r/min 摇床培养至OD600值为0.6~1.0,2500×g 离心5 min 后,收集菌体,置换到M9 培养基中培养1 h,加入20% L-ara,37 ℃诱导5 h,分2 次加入硒代甲硫氨酸,诱导开始时及诱导2 h 后各添加一次硒代甲硫氨酸,每次加入量为0.25 mg,诱导完成后,收集菌体,加水溶解,100 ℃煮沸15 min,取上清,SDS-PAGE 点胶验证蛋白表达情况。
通过实验组1 确定蛋白表达后进行大量诱导实验(实验组2)。按照1:50 比例将活化液接种于500 mL 过渡培养基扩大培养(实验组2),待其OD600值为0.6~0.8 时,将菌体置换到完全M9 培养基中培养1 h。加入20% L-ara,37 ℃诱导5 h,分2 次加入硒代甲硫氨酸,诱导开始时及诱导2 h 后各添加一次硒代甲硫氨酸,每次加入量为0.0125 g,使其终浓度为0.05 g/L。诱导完成后,收集菌体,硒代蛋白纯化过程与重组蛋白一致,见方法1.2.2。
1.2.4 重组Bsa I 蛋白和Se-Bsa I 蛋白的质谱检测 为验证 Se-Bsa I 蛋白中硫原子被硒原子的取代情况,采用AB Sciex 5800 MALDI-TOF/TOF 质谱仪对 Bsa I 重组蛋白和其硒代蛋白进行分析鉴定,质谱检测在华中科技大学完成。
1.2.5 重组Bsa I 蛋白和Se-Bsa I 蛋白圆二色谱检测 为初步检验重组Bsa I 蛋白与Se-Bsa I 蛋白二级结构的区别,将重组蛋白和硒代蛋白稀释至0.5 mg/mL,并使用圆二色谱仪对其进行扫描,样品池光程为0.5 mm,近紫外区波长范围为190~240 nm,分析其二级结构有无差异。
1.2.6 重组Bsa I 蛋白和Se-Bsa I 蛋白酶活检测重组Bsa I 蛋白(1 mg/mL)或Se-Bsa I 蛋白(1 mg/mL)各1 μL,底物为1 μg 的λDNA,配制成20 μL 的反应体系,在37 ℃的条件下反应30 min,反应结束后高温失活。使用1 %琼脂糖凝胶电泳,鉴定Bsa I 蛋白或Se-Bsa I 蛋白对λDNA 切割条带。
1.2.7 重组Bsa I 蛋白结晶条件初筛 将蛋白浓缩到10 mg/mL,通过气相扩散坐滴法对9 种结晶试剂盒(Crystal Screen Cryo-HR2-112、PEGRx 1 -HR2-082、Index-HR2-144、Crystal Screen Lite-HR2-128、Crystal Screen Cryo -HR2-122、SaltRx 1 -HR2-107、Crystal Screen-HR2-110、Natrix-HR2-116、PEGRx 2-HR2-084)条件进行蛋白晶体生长条件的初步探索。使用NT8 机械手臂将蛋白溶液和生长池液按照1:1 的比例混合,点样至晶体板的样品槽内,并使用透明塑封纸封装。放入全自动晶体生长观察成像分析仪,设置温度为20 ℃,间隔12 h 拍照,观察晶体在不同条件下的生长情况。蛋白在2 种情况下生长出晶体,条件a:0.2 mol/L 醋酸镁四水合物,0.1 mol/L二甲胂酸钠三水合物 pH6.5,20%聚乙二醇8000;条件b:0.1 mol/L 三水醋酸钠 pH4.6,2.0 mol/L 硫酸铵。
2 结果与分析
2.1 重组表达载体的构建
利用原核生物表达体系表达内切酶,首先要考虑的问题是避免重组内切酶对宿主DNA 的切割伤害。为解决这个难题,本研究在载体pBAD 上同时构建了Bsa I 基因和Bsa I 的甲基化酶基因。如图1所示,在Bsa I 基因和Bsa I-M 基因的上下游分别构建了启动子和终止子,使两个蛋白分别表达。另外,在Bsa I 基因的上游加入6×His 标签序列,使目的蛋白Bsa I 可以带有融合标签6×His。将构建成功的重组质粒通过热激化法转化到ER2566 感受态细胞中,获得Bsa I 重组蛋白的表达菌株。

图1 pBAD-Bsa I 质粒图谱Fig.1 Schematic of pBAD-Bsa I construct
2.2 Bsa I 重组蛋白的表达和纯化
首先将构建成功的Bsa I 重组表达菌株ER2566进行诱导实验,以确定目的蛋白表达情况。限制性内切酶Bsa I 重组蛋白的分子质量约为65 kDa。将表达菌株在37 ℃下诱导5 h,小量实验验证(实验组1)泳道2 比泳道1 多了一条诱导后条带。与预期65 kDa 大小相仿,证明重组Bsa I 蛋白在ER2566 菌株中有诱导表达。然后进行大规模发酵实验(实验组2),将诱导后的发酵液进行超高压破碎,如图2a泳道3,在目的蛋白与镍基至结合后,利用高浓度咪唑洗脱,如图2a泳道4 所示,经过Ni 亲和层析后重组Bsa I 蛋白的纯度明显提升。但重组Bsa I 蛋白的纯度还未达到蛋白晶体生长所需的纯度,因此对重组Bsa I 蛋白利用阴离子交换层析进一步纯化,如图2c 所示,蛋白纯度可以达到90%以上。最终,每升发酵液得到了6.6 mg 的目的蛋白。

图2 重组蛋白Bsa I 的表达与纯化Fig.2 Expression and purification of Bsa I
2.3 Se-Bsa I 硒代蛋白的表达与纯化
实验室最常用的方法是使用甲氨酸缺陷型菌株制备硒代蛋白,利用M9 培养基发酵获得硒代蛋白,其中大肠杆菌常用甲硫氨酸缺陷型菌株是B834[24]。因此,本研究将构建成功的重组质粒转化导入B834(DE3)菌株中,并进行目的蛋白的小量表达试验。经过SDS-PAGE 检测发现没有硒代蛋白的目的条带。推测其结果可能是甲硫氨酸缺陷型菌株B834(DE3)不适合同时诱导表达Bsa I 基因和Bsa I-M 基因,B834 菌株中的甲基化修饰系统阻碍了表达Bsa I 的甲基化酶,而无法获得目的蛋白Bsa I。程艺等[25]在表达重组Se-Nco I 时,利用甲硫氨酸缺陷型菌株B834时蛋白未表达,之后利用表达重组Nco I 的BL21(DE3)pLysS 菌株和M9 培养基成功表达出硒代蛋白。
因此,本实验采用原ER2566 表达菌株,利用M9 培养基来替换甲硫氨酸为硒代甲硫氨酸,进行Se-Bsa I 蛋白的诱导表达,诱导后的重组Bsa I 蛋白在硒代培养基中有诱导条带产生,表明此方法是可行的。但是诱导后的菌体量少,无法获得足够的硒代蛋白。推测是由于硒代甲硫氨酸的细胞毒性[26]以及M9 培养基不是大肠杆菌的最适培养基,会严重影响菌体的生长,从而降低Se-Bsa I 重组蛋白的表达量。因此,本实验通过分批次加入硒代甲硫氨酸,减少硒代甲硫氨酸的浓度,从而降低硒代甲硫氨酸的细胞毒性。通过小量实验(实验组1)表明分批次加入可以提高诱导后菌体量,从而提高硒代蛋白的产量。大规模发酵(实验组2)经过Ni 亲和层析和阴离子交换层析的纯化,如图3a 泳道4 和图3c 所示。最终,每升发酵产物得到4 mg 目的蛋白。

图3 重组蛋白Se-Bsa I 的表达与纯化Fig.3 Expression and purification of Se-Bsa I
邵钰晨等[27]在制备硒代蛋白时也发现了此类问题,指出M9 培养基对于LB 培养基来说,营养成分相对贫瘠,因此邵钰晨等在培养过程中添加异亮氨酸等多种氨基酸,抑制大肠杆菌本底代谢,从而提高外源硒代甲硫氨酸利用率,提高硒代蛋白表达量。通过这些改进最终获得了浓度为10 mg/mL、纯度在95%以上的Se-Mlu I 蛋白。后续实验将针对硒代蛋白诱导条件进行优化,以达到满足晶体生长的蛋白浓度。
SDS-PAGE 检测结果显示Se-Bsa I 硒代蛋白的大小和Bsa I 蛋白大小类似,但电泳图中很难判断Se-Bsa I 蛋白是否被硒代,因此后续进行质谱检测Se-Bsa I 蛋白的分子量与Bsa I 蛋白的区别,从而判断Se-Bsa I 蛋白中甲硫氨酸的硒代概率。
2.4 重组蛋白Bsa I 和Se-Bsa I 的质谱检测
由于采用原ER2566 表达菌株,利用M9 培养基掺入硒代甲硫氨酸来替换目的蛋白中的甲硫氨酸,存在替换不完全的风险。为了检测Se-Bsa I 蛋白中硒原子的取代率,将纯化得到的Bsa I 和Se-Bsa I 取样进行质谱检测如图4 所示。经过质谱检测发现,重组Bsa I 蛋白的分子质量为65278.87 Da,这与Bsa I 理论相对分子质量65 kDa 相符。而重组Se-Bsa I 蛋白相对分子质量为65758.26 Da,与Bsa I 重组蛋白之间相差479.39 Da。已知硫元素相对分子质量为32.065 Da,硒元素的相对分子质量为78.96 Da,因此平均每分子Se-Bsa I 中被硒原子取代的硫原子数量为479.39/(78.96-32.065)=10.22。通过分析重组Bsa I 蛋白质的氨基酸序列发现其有11 个甲硫氨酸,存在11 个硫原子。因此,推测出本批次的Se-Bsa I 蛋白质中甲硫氨酸被硒代甲硫氨酸取代的概率约92.91%,样品适合进行后续X-射线衍射实验[28]。

图4 Bsa I 和Se-Bsa I 质谱图Fig.4 Serum mass spectrometry of Bsa I and Se-Bsa I
2.5 重组蛋白Bsa I 和Se-Bsa I 的圆二色谱图检测
为了解析限制性内切酶Bsa I 的三维结构,Se-Bsa I 重组蛋白样品与Bsa I 重组蛋白样品的结构必须高度相似。测定蛋白质二级结构最常用的方法是圆二色谱法,其广泛应用于蛋白质的构象研究中[29]。为了进一步明确硒代蛋白对重组蛋白的结构有无影响,本研究将重组蛋白样品Bsa I 和 Se-Bsa I 进行圆二色谱检测,解析其二级结构(图5)。CD 图谱均显示在192 nm 处有一个正峰,208、222 nm 处有两个负峰,这表明Bsa I 和Se-Bsa I 都是以α-螺旋的形式存在,说明蛋白折叠正确,构象稳定。结果表明,Bsa I 和Se-Bsa I 在二级结构上无差异,硒代不影响其结构。为后续功能以及结构研究奠定了很好的基础。
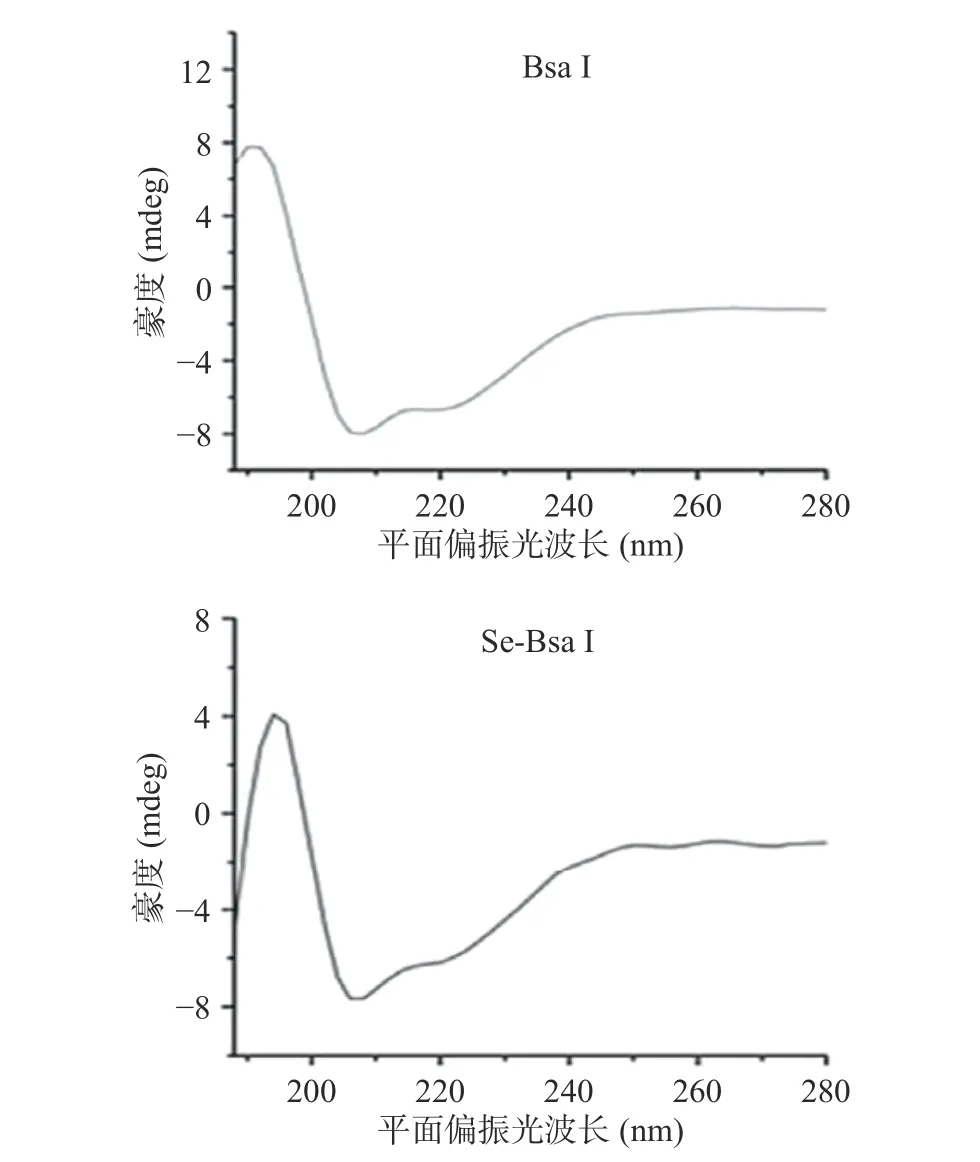
图5 Bsa I 和Se-Bsa I 圆二色谱图Fig.5 Circular dichromatogram of Bsa I and Se-Bsa I
2.6 蛋白及其硒代蛋白的酶活检测
本研究分别对Bsa I 重组蛋白与Se-Bsa I 重组蛋白进行酶活力检测,进一步检测硒代是否影响野生型蛋白的生物学功能。取1 μg Bsa I 和1 μg Se-Bsa I 蛋白,对1 μgλDNA 进行30 min 快速酶切实验,结果见图6。经琼脂糖凝胶电泳检测发现,Bsa I 和Se-Bsa I 都有明显的特征酶切条带,与Thermo Fisher公司网址中公布的Bsa I 特征酶切条带相似。酶切结果表明了Se-Bsa I 蛋白与野生型蛋白具有相似的生物学活性,即硒代蛋白活性并未受到硒代影响。

图6 Bsa I 和Se-Bsa I 的酶活验证Fig.6 Enzymes activity of Bsa I and Se-Bsa I
2.7 Bsa I 蛋白结晶生长条件筛选
本实验将重组后的Bsa I 蛋白利用坐滴法进行蛋白质晶体的初筛。初筛使用购自Hampton Research公司的商品化蛋白质晶体筛选试剂盒,蛋白质液滴与点晶体等体积混合后,于20 ℃的全自动晶体生长观察成像分析仪中进行晶体培养,每隔12 h 自动拍照采集数据[30-31]。在使用商品化蛋白结晶试剂盒进行蛋白结晶,最初得到的蛋白结晶条件往往不是最优条件,大概率会出现晶体过小,分辨率较差,衍射能力较低等情况[32-33]。筛选结果显示有2 个条件生长晶体,晶体如图7 所示。

图7 Bsa I 的结晶条件筛选Fig.7 Screening of crystallization conditions of Bsa I
蛋白晶体可以在a 条件下(0.2 mol/L 醋酸镁四水合物,0.1 mol/L 二甲胂酸钠三水合物 pH6.5,20%聚乙二醇8000)生长出较小的颗粒状结晶,并且可以在b 条件(0.1 mol/L 三水醋酸钠pH4.6,2 mol/L硫酸铵)下生长出不规则的球形晶体,但初步结晶条件所筛选到的蛋白结晶较为脆弱且边缘不规则,无法捞出进行衍射实验。针对这种情况,程焯等[34]在制备LxmX 蛋白时,通过利用最初筛选得到的晶体生长条件,设计正交试验,成功得到了边缘整齐,可用于衍射数据收集的LxmX 蛋白及硒代LxmX 蛋白晶体。因此后续实验将对初步得到的蛋白结晶条件进行优化,以获得高分辨率的晶体。此外,通过观察所有晶体的生长情况,发现大部分蛋白都发生了沉淀,推测Bsa I 的稳定性可能是影响其蛋白晶体生长的一个关键因素[35-37]。
3 结论
限制性内切酶是分子生物学最常用的工具之一,而内切酶的结构决定其功能。但大部分内切酶结构没有被解析,这限制了内切酶的发展。通过制备硒代衍生物进行衍射解析结构,其操作更为简单,适用性更高。但在制备硒代蛋白过程中,B834 甲硫氨酸缺陷型菌株往往易受到外源基因的影响,无法获得重组蛋白,具有局限性,同时硒代甲硫氨酸的毒性问题以及发酵条件鲜有报道,这也限制了制备硒代蛋白的成功率。本实验利用限制性内切酶Bsa I 和甲基化酶Bsa I-M 的大肠杆菌共表达系统,获得大量表达的Bsa I 蛋白,通过M9 培养基添加硒代甲硫氨酸的方式,获得了大小、性质和重组蛋白一致的硒代蛋白,并在两种结晶条件下培养出蛋白晶体,为后续Bsa I 蛋白的晶体衍射,解析其三维结构奠定了基础。
猜你喜欢
食品科学(2023年13期)2023-08-12
中国生物化学与分子生物学报(2022年7期)2022-09-07
草地学报(2022年3期)2022-03-28
中国药科大学学报(2021年6期)2021-12-31
今日农业(2021年11期)2021-11-27
人工晶体学报(2021年10期)2021-11-26
分析测试学报(2015年9期)2015-12-17
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年15期)2014-03-11
机械制造与自动化(2014年1期)2014-03-01
