
水分添加量对高水分挤压大豆粕植物蛋白肉品质及结构特性的影响
2023-11-20 07:45岳程程佟丽凤邢维臣段玉敏王致力肖志刚
食品工业科技 2023年22期
岳程程,王 哲,佟丽凤,邢维臣,段玉敏,王致力,肖志刚,王 鹏,
(1.沈阳师范大学,辽宁沈阳 110034;2.东方集团哈尔滨福肴食品有限公司,黑龙江哈尔滨 150001)
大豆是重要的油料作物和植物蛋白来源之一,在农业中占有重要的地位。低温脱脂大豆粕(简称大豆粕)是大豆榨油后的剩余副产物,其含有35%~55%的蛋白质以及丰富的功能因子,如大豆异黄酮、大豆膳食纤维等[1],常作为饲料行业的加工原料。虽然大豆粕含有较丰富的植物蛋白质资源,但由于其经济价值低,造成了优质植物蛋白资源的浪费,因此,有必要对大豆粕开展高值化开发利用。近年来,作为动物肉替代品的高水分挤压植物蛋白肉受到了越来越多的关注[2-3],由于大豆分离蛋白中含有近90%的球蛋白和5%的白蛋白,使其成为加工植物蛋白肉的理想原料[4]。大豆分离蛋白主要是从大豆粕中分离提取的,其使用成本要远高于大豆粕,同时,以大豆分离蛋白作为植物蛋白肉的加工原料也会直接推高产品的生产成本。此外,大豆粕中的部分不溶性膳食纤维在挤压过程中会转变为可溶性膳食纤维,这会影响挤压产品的流变性能及品质[5],因此,探究利用大豆粕替代大豆分离蛋白作为高水分挤压植物蛋白肉加工原料的可能性,对生产企业控制生产成本及提高大豆粕经济价值都具有重要意义。
高水分挤压技术是新兴的植物蛋白重组技术[6],挤压原料的水分含量通常达到60%以上,采用该技术制成的产品质地紧密、口感细腻,具有类似于动物肌肉的纤维组织[7],而且无需复水即可直接食用。由于水分具有增塑、导热、降黏、熟化和利于成型的作用,因此,在挤压过程中的水分添加量对植物蛋白肉的顺利挤出和最终产品特性的形成有着十分重要的作用。产品原料被水浸润软化后,其熔融温度低于热分解温度,易于后续的挤压加工和营养成分的保护[8]。此外,水分还可以作为反应溶剂降低物料的黏度和在机筒内的停留时间,进而降低蛋白质亚基之间的聚合交联,增加二硫键与氢键、疏水作用之间的协同作用,并影响产品的色泽,提高产品的纤维化程度[9]。Lin 等[10]以大豆分离蛋白和小麦粉为原料,通过研究发现物料含水率是影响挤压过程最重要的因素。张玉洁等[11]以大豆分离蛋白(SPI)和金线鱼鱼糜为原料,证实了水热参数组合作用显著(P<0.05)影响SPI-鱼糜混合物的微观结构以及质构特性,并且物料水分的影响更显著。
为了探究大豆粕能否作为高水分挤压植物蛋白肉的加工原料,本文以低温脱脂大豆粕为原料,固定进料速度、螺杆转速、机筒温度等条件,考察水分添加量对挤压植物蛋白肉理化性质和微观结构的影响。本文的研究结果将为大豆粕作为原料在高水分挤压植物蛋白肉中的应用提供依据和参考。
1 材料与方法
1.1 材料与仪器
低温脱脂大豆粕(水分含量10.62%;蛋白含量38.5%(干基);纤维含量4.33%(干基);灰分含量4.28%(干基))黑龙江冬雪生物科技有限公司;磷酸盐博迈德生物化学试剂有限公司;尿素 天津市瑞金特化学品有限公司;巯基乙醇 麦克林试剂有限公司;十二烷基硫酸钠 Coolaber Science &Technology。
UVTE-36 双螺杆挤压机 长沙创享食品科技有限公司;CT3 4500 型质构仪 美国Brookfield 公司;CR-400 型色彩色差计 日本柯尼卡美能达有限公司;TA Q20 差式扫描量热仪 美国TA 公司;贝克曼冷冻离心机 美国贝克曼库尔特公司;SU3500 扫描电子显微镜 日本Hitachi Ltd 公司。
1.2 实验方法
1.2.1 大豆粕植物肉挤压实验 根据挤压机的构造特点和预实验结果,将水分添加量分别设置为10.5、11、11.5、12 和12.5 kg/h(通过预实验测得上述水分添加量范围内制备的植物肉含水量皆大于60%,所对应的含水量分别为61%、62%、63%、64%和65%),其他挤压参数不变,将粉碎且过100 目筛的低温脱脂大豆粕以8 kg/h 的速度从喂料区加入到双螺杆挤压机机筒中,设定螺杆转速为220 r/min,机筒共开启六段加热区,温度分别设定为(40±1)、(60±1)、(90±1)、(120±1)、(150±1)和(160±1)℃,冷却装置温度设定在(55±1)℃,进行挤压试验。将挤出的植物蛋白肉冷却至室温后,对一部分试样进行质构和颜色测试,另一部分试样经真空冷冻干燥后粉碎,并过100 目筛后进行后续分析。
1.2.2 豆粕植物肉色泽测定 采用色差仪测定植物蛋白肉的L*、a*、b*的数值。对每个样品3 个不同位置的色泽进行测定,每个实验测定至少进行3 次,去掉异常值,取平均值。
1.2.3 豆粕植物肉质构特性测定 参考肖志刚等[12]的方法并做适当修改。将植物蛋白肉样品裁剪为长宽均为25 mm 高为10 mm 的长方体,设置质构仪操作条件:TPA 模式,探头选用TA-5,测试前速度1.0 mm/s,测试速度1.0 mm/s,测试后速度1.0 mm/s,下压程度50%,测定样品硬度、弹性、咀嚼性的数值,每个样品进行3 次平行测定,取平均值。
1.2.4 豆粕植物肉持水性、持油性测定
1.2.4.1 豆粕植物肉持水性 参考Mazaheri 等[13]的方法并加以改进。称取1.5 g 植物蛋白肉样品放入到50 mL 的离心管中,然后加入30 mL 的去离子水,室温下涡旋10 min 后于4000 r/min 离心20 min,然后倒掉上清液,植物蛋白肉的持水性根据下列公式进行计算:
式中:m 为样品的质量,g;m1为样品的质量+离心管质量,g;m2为离心管除去上清液后的质量,g。
1.2.4.2 豆粕植物肉持油性 参考Mazaheri 等[13]的方法并加以改进。称取1.5 g 植物蛋白肉样品放入到50 mL 的离心管中,然加入30 mL 的葵花籽油,室温下涡旋10 min 后于4000 r/min 离心20 min,然后倒掉上清液,植物蛋白肉的持油性根据下列公式进行计算:
式中:m 为样品的质量,g;m1为样品的质量+离心管质量,g;m2为离心管除去上清液后的质量,g。
1.2.5 豆粕植物肉热特性分析 参考张金闯[14]的方法并加以改进。称取5 mg 样品置于耐高压铝盘中,密封。使用TA Q20 差式扫描量热仪,扫描温度区间25~130 ℃、升温速率5 ℃/min,N2保护,N2流速为50 mL/min。空铝盘做参比。记录热转变起始温度(To)、热转变峰值温度(TP)、热转变焓值(ΔH)和峰宽(ΔT),每个样品重复测定3 次,结果取平均值。
1.2.6 豆粕植物肉SDS-PAGE 凝胶电泳测定 利用SDS-PAGE 凝胶电泳测定植物蛋白肉的相对分子量,具体操作参考张金闯[14]的方法并加以改进。将不同水分添加量挤出样品经过冷冻干燥后,粉碎,过100 目筛网。将过筛样品(2 mg)溶于0.5 mL 样品缓冲液(0.08 mol/L Tris-HCl 缓冲液,pH6.8),1%(w/v)SDS,2%(v/v)2-ME,5%(v/v)甘油和0.025%(w/v)溴酚蓝并充分混合。然后将样品在沸水浴中加热10 min,12000 r/min 离心10 min。进样时取上清5 μL进样,Marker 分子量范围为10000 至250000 Da。在Mini-PROTEAN 系统上进行电泳,所配制电泳胶为5%的浓缩胶和12%的分离胶。电泳选取恒压模式,首先电压选用80 V,溴酚蓝条带跑至浓缩胶底端时换用110 V 电压,等溴酚蓝条带跑至底端停止。采用考马斯亮蓝R250 振荡染色60 min。染色完成后,用脱色液进行振荡脱色10 h,每隔1 h 更换一次脱色液。使用Gel DocTMXR+凝胶成像系统拍摄蛋白质电泳光谱的图像。
1.2.7 豆粕植物肉扫描电镜观察 将冷冻干燥后的植物蛋白肉样品进行横纵方向切片并固定在样品台上。采用Hitachi IB-5 离子镀膜仪进行喷金处理后,置于SU8010 扫描电子显微镜下放大300 倍观察并拍照。
1.2.8 豆粕植物肉化学交联作用测定 参考肖志刚等[15]的方法并加以改进。取粉末状样品0.25 g(精确至0.1 mg)于25 mL 烧杯中,分别加入10 mL 八种溶剂:a.0.035 mol·L-1pH7.6 磷酸盐缓冲溶液(P);b.磷酸盐缓冲液中尿素含量为8 mol·L-1尿素(P+U);c.磷酸盐缓冲溶液中2-巯基乙醇(2-ME)0.1 mol·L-1(P+M);d.磷酸缓冲液中十二烷基硫酸钠(SDS)1.5 g·100 mL-1(P+S);e.磷酸盐缓冲溶液中尿素和2-ME 分别为8 和0.1 mol·L-1(P+U+M);f.磷酸盐缓冲液中SDS 和尿素分别为1.5 g·100 mL-1和8 mol·L-1(P+U+S);g.磷酸盐缓冲液中SDS 和2-ME 分别为1.5 g·100 mL-1和0.1 mol·L-1(P+S+M);h.磷酸盐缓冲溶液中尿素为8 mol·L-1,SDS 为1.5 g·100 mL-1,2-ME 为0.1 mol·L-1(P+U+S+M),常温下搅拌浸提1 h 后放入冰箱4 ℃保存,离心(12000×g,10 ℃,15 min),取上清液。沉淀中再分别加入10 mL 上述八种溶剂,重复浸提两次,离心。取1 mL 上清液于1.5 mL 离心管中(36×3 个),混匀(摇床160 r/min,1 h)。加20 μL 到96 孔酶标板的样品孔中,采用Lowery法测得(测定波长650 nm)上清液中可溶性蛋白含量,原料和挤出样品中总的蛋白质含量采用凯氏定氮法测定,蛋白质溶解度计算方法为上清液中可溶性蛋白与样品中总蛋白的比值。每次测量重复三次。蛋白质溶解度,总蛋白质量浓度根据下列公式进行计算:
根据不同溶液中的溶解度计算各种化学键:天然状态蛋白:a;氢键作用:b-a;二硫键:c-a;疏水作用:d-a;氢键和二硫键交叉作用:e+a-b-c;氢键和疏水作用交叉作用:f+a-b-d;疏水和二硫键交叉作用:g+a-c-d;氢键和二硫键和疏水作用交叉作用:h+b+c+d-a-e-f-g。
1.2.9 豆粕植物肉蛋白质二级结构分析 参考Guo等[16]的方法并加以改进,将1 mg 样品和100 mg 溴化钾称重制成透明片剂,扫描范围设置为1600~1700 cm-1,分辨率为4 cm-1和信号扫描64 次。使用Peakfit 版本4.12 软件处理光谱。
1.3 数据处理
数据结果分析采用SPSS 20.0 统计软件和Origin 2018 软件进行分析及图形绘制。通过单因素方差分析(ANOVA)确定显著性,然后进行Duncan 检验,通过Pearson 相关系数法对实验数据进行相关性分析,所得试验结果均采用三次重复试验的平均值和标准差表示。
2 结果与分析
2.1 水分添加量对植物蛋白肉外观色泽的影响
水分添加量对植物蛋白肉外观色泽的影响如表1 所示,随着水分添加量从10.5 kg/h 增加到11.5 kg/h时,植物蛋白肉亮度值(L*)逐渐增加,而红绿值(a*)逐渐下降,这表明适当增加水分含量有利于提高植物蛋白肉产品的外观色泽。水分添加量的增加促进了组织化蛋白形成更多的定向纤维和多孔结构,并且随着纤维化程度的提高,在植物蛋白肉的表面形成了一层光滑的薄膜结构[8];同时水分含量增加也会导致物料粘度降低,这减少了推动物料通过机筒所需的力以及物料与螺杆和机筒之间的摩擦,缩短了物料在挤压机中的停留时间,降低了美拉德反应引起的褐变程度[17],从而提高了植物蛋白肉的光泽度。此外,随着水分含量的增加,水分子与受热断开的蛋白质链上的亲水结合位点的结合更加紧密,塑化作用增强,这可能与颜色L*的增加有关。陈锋亮等[18]和张汆[19]也发现了物料水分的增加会促使大豆蛋白素肉或花生蛋白素肉变得更亮白。然而,当水分添加量从11.5 kg/h进一步升高到12.5 kg/h 时,植物蛋白肉的亮度值呈下降趋势,这可能是因为当水分添加量过多时,物料的黏度也随之升高,物料在机筒内的滞留时间变长,因此加剧了物料之间发生美拉德反应的程度[20],从而使植物肉的外观色泽下降。实验结果表明,适量的物料水分含量有助于提高植物蛋白肉产品的外观色泽。
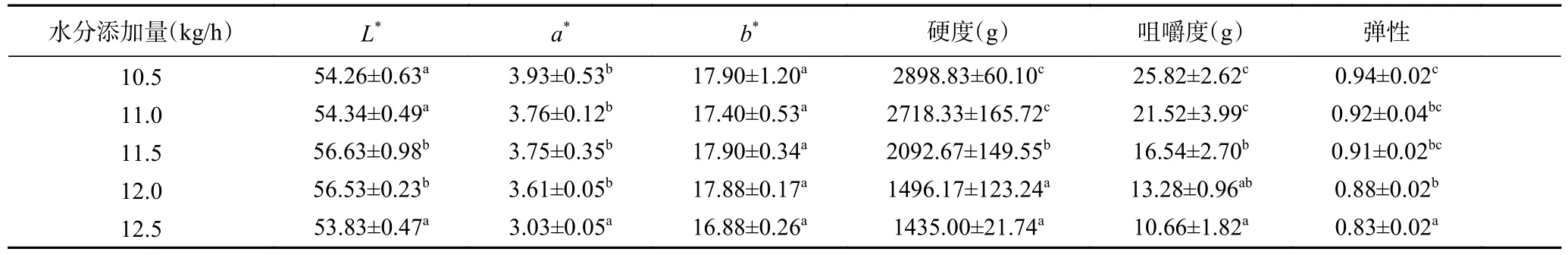
表1 水分添加量对植物蛋白肉外观色泽、质构特性的影响Table 1 Effect of water addition on appearance and color and texture properties of plant protein meat
2.2 水分添加量对植物蛋白肉质构特性的影响
表1 列出了水分添加量对植物蛋白肉质构特性的影响结果。从表1 中可以看出,不同的水分添加量对植物蛋白肉的硬度、咀嚼度和弹性具有显著影响(P<0.05)。随着水分添加量的逐渐增加,物料在机筒内结合了过多的水分,导致植物蛋白肉的质地变得愈加柔软,其硬度、咀嚼度和弹性均显著降低(P<0.05),尤其以硬度变化最为明显,植物蛋白肉的硬度由2898.83 g 显著降至1435.00 g(P<0.05),而咀嚼度和弹性则分别由25.82 g 和0.94 降至10.66 g和0.83,由此可知,通过调节水分添加量能够调控植物蛋白肉产品的口感。
2.3 水分添加量对植物蛋白肉持水性、持油性的影响
水分添加量对植物蛋白肉持水性、持油性的影响如图1 所示,随着水分添加量的增加,植物蛋白肉的持水性、持油性均呈现出先升高后降低的趋势。当水分添加量低于11.5 kg/h 时,植物蛋白肉持水性和持油性的升高主要是因为水分在挤压过程中起到了增塑的作用[21],一定量的水分可以加快聚合物分子的运动,促进蛋白质-蛋白质及蛋白质-水的相互作用,从而促使更多的疏水基团暴露,这加速了蛋白质的变性,并引起蛋白质的凝聚度下降,蛋白质的溶胀特性和形成网络结构的能力进一步得到增强,产品组织化程度下降,从而导致植物蛋白肉的持水和持油能力均得到提高[22]。此外,豆粕中的不溶性膳食纤维在挤压的作用下会转变为可溶性膳食纤维,这也有助于提高植物蛋白肉的持水能力[23]。水分添加量由12.0 kg/h增加至12.5 kg/h 时,植物蛋白肉的持水性和持油性均呈下降趋势,这可能是由于过量的水分阻碍了蛋白质与蛋白质之间的相互作用,导致形成了较弱的凝胶网络[24],从而造成植物蛋白肉持水性和持油性的下降。可以看出,适当提高水分添加量有利于增强植物蛋白肉的持水和持油能力。
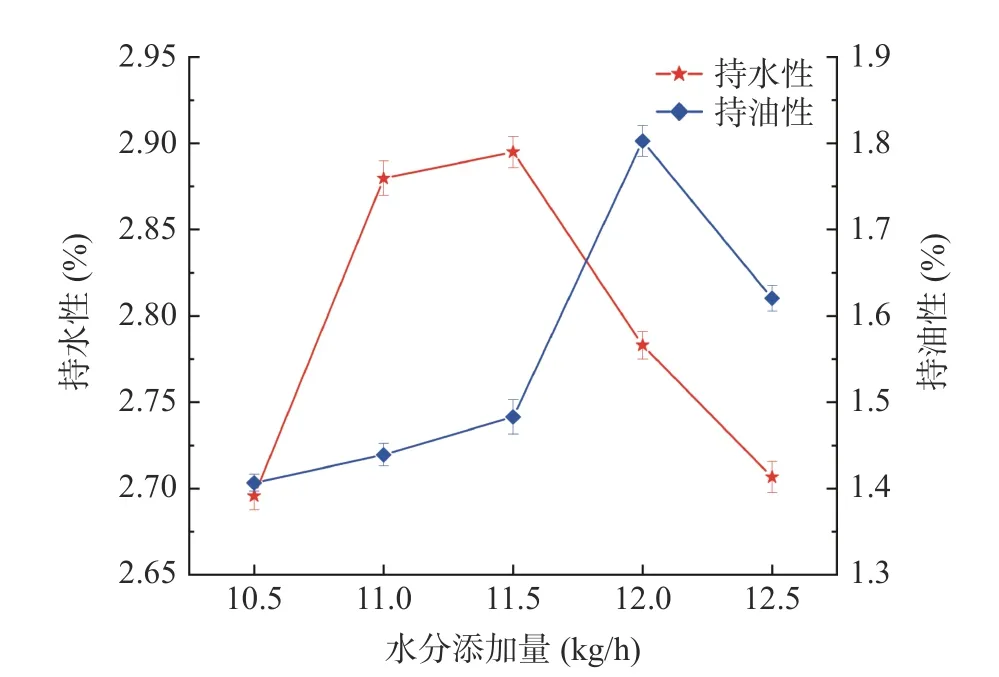
图1 水分添加量对植物蛋白肉持水性、持油性的影响Fig.1 Effects of water addition on water absorption capacity,oil absorption capacity of plant protein meat
2.4 水分添加量对植物蛋白肉热特性的影响
水分添加量对植物蛋白肉热特性的影响如表2所示,由于豆粕中大豆蛋白11S 亚基的热转变焓值明显高于7S 亚基,其热转变峰更明显,因此,在本研究中,植物蛋白肉的热特性变化仅以11S 亚基为代表进行分析[18]。当水分添加量由10.5 kg/h 增加到11.0 kg/h 时,植物蛋白肉发生热转变的起始温度(To)和峰值温度(Tp)均呈现出上升的趋势,而峰宽(ΔT)和热转变焓值(ΔH)却出现明显下降,这可能与水分含量的突然增大有关。物料水分含量的突然增大会使蛋白分子结构变得灵活[25],分子结构更易于被打开[26],从而降低了植物蛋白肉的热稳定性。当水分添加量由11.0 kg/h 继续升高到12.5 kg/h 时,植物蛋白肉发生热转变的起始温度(To)和峰值温度(Tp)随之逐渐下降,但峰宽(ΔT)和热转变焓值(ΔH)却逐渐升高,To和Tp分别从78.84 和101.21 ℃降低至67.23和96.31 ℃,ΔT 和ΔH 分别从34.15 ℃和26.75 J/g升高至58.71 ℃和57.88 J/g,这表明水分含量进一步的升高加快了植物蛋白肉中大豆蛋白的热转变速率。而植物蛋白肉中大豆蛋白的热焓值△H 显著增大,说明其分子结构更加稳固,打开其分子结构所需要的能量明显增加。
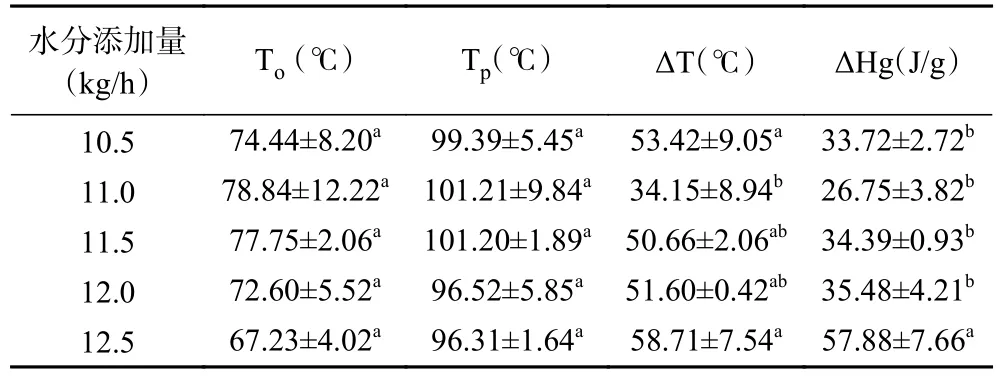
表2 水分添加量对植物蛋白肉热特性的影响Table 2 Effect of water addition on thermal properties of plant protein meat
2.5 水分添加量对植物蛋白肉相对分子量的影响
豆粕中的大豆蛋白主要组分包括7S 亚基(分子量分别为78.41、73.00、54.05 kDa)和11S 亚基(分子量分别为42.18、37.62、34.03、31.58、22.05、19.02 kDa)[18]。从图2 的SDS-PAGE 电泳结果中可以看出,随着挤压过程中水分添加量的升高,植物蛋白肉样品中的7S/11S 球蛋白比例也发生了显著的变化。当水分添加量为10.5 kg/h 时,植物蛋白肉样品电泳条带中亚基的颜色与条带顶端颜色均很浅。随着水分添加量的进一步增加,7S 中的α′亚基(78.41 kDa)、α亚基(73.00 kDa)、β亚基(54.05 kDa)以及11S 中的A 组分(42.18 kDa)和B 组分(22.05 kDa)的亚基条带逐渐变宽,并且亚基条带颜色以及挤出样品条带顶端的颜色逐渐变深,这可能是因为随着水分添加量的增加,豆粕中的大豆蛋白经挤压蒸煮后发生聚合、交联等变性反应的程度增强,形成了分子量较大的聚合物[27]。然而,当水分添加量由12 kg/h 增加到12.5 kg/h 时,植物蛋白肉样品条带顶端的颜色略微变浅,这可能是因为形成的较大分子量的聚合物可以透过浓缩胶,但却不能透过分离胶的网络孔隙,因此在条带的顶端发生了聚集[28]。可以看出,适当提高水分添加量有利于提高蛋白质聚合交联变性的程度。

图2 水分添加量对植物蛋白肉相对分子量的影响Fig.2 Effect of water addition on relative molecular weight of plant protein meat
2.6 水分添加量对植物蛋白肉微观形貌的影响
扫描电子显微镜观察植物蛋白肉的形态如图3所示。在挤压过程中,由于豆粕中含有的纤维成分与蛋白质流动性能不同,在高温、高剪切条件下两者发生相分离,从而在冷却装置中逐渐形成具有拉丝效果的纤维结构[29]。当水分添加量为10.5 kg/h 时,植物蛋白肉拉丝效果并不明显,所形成的纤维结构内部孔洞较大,出现了明显分层的现象,这主要是因为在相对较低的水分含量下,植物蛋白肉在冷却模具中受到较高摩擦力和剪切力所导致[30]。随着水分添加量增加至11.5 kg/h 时,植物蛋白肉表面形成了更加致密的纤维细丝结构,组织蛋白层状结构清晰且整齐,具有精细的定向结构,这主要是由于合适的水分含量促进疏水基团暴露[31],加速了蛋白质的变性[32],使豆粕中的蛋白质分子由最初的无序多孔结构变为结构致密且具有一定规则的晶型排列结构,这有利于三维网络结构的形成,从而使植物蛋白肉形成更多具有类似于肉类纤维的条状纤维结构。然而随着水分添加量继续升高至12.5 kg/h 时,植物蛋白肉形成了更多层状而非纤维状的结构且疏松多孔,这主要是因为随着水分添加量的升高,机筒的高温导致更多的水蒸汽产生,蛋白质形成了更多的膜状气孔结构[9],这加剧了蛋白质内部疏水基团的暴露和蛋白质交联变性程度的增大,造成产品气孔结构更加明显。此外,过量的水分促使豆粕中更多的糖类物质溶于水中,加剧了蛋白质的溶胀程度[28],使得蛋白质-蛋白质相互作用减弱,产生了不相容的聚合物网络[33],从而导致植物蛋白肉形成了更多分离的层状结构。
2.7 水分添加量对植物蛋白肉化学交联作用力的影响
水分添加量对植物蛋白肉化学交联作用力的影响如表3 所示,植物蛋白肉的结构是由疏水作用、氢键、二硫键以及彼此之间的交互作用共同维持的。从表3 可以看出,随着水分添加量的增加,植物蛋白肉中的氢键数量和疏水作用均随之升高,这表明提高水分含量能够促进蛋白质与水混合,从而形成更多的氢键[16]。当水分添加量由10.5 kg/h 增加到11 kg/h时,植物蛋白肉中二硫键显著地增加(P<0.05),而当水分添加量进一步增加时二硫键数量则无显著变化。Sheard 等[34]认为当物料水分含量较高时,疏水作用是维持蛋白聚集的主要作用力;而随着水分含量的降低,二硫键逐渐形成并成为稳定结构的主要作用力。此外,当水分添加量由10.5 kg/h 增加到12.5 kg/h时,氢键、疏水相互作用、二硫键三者交叉作用呈现先升高后下降的趋势,且在水分添加量为11.5 kg/h时,氢键、疏水相互作用、二硫键三者交叉作用达到最强,由此可知,适当地提高水分添加量有利于蛋白质变性,对氢键、疏水相互作用、二硫键三者交互作用具有促进作用。
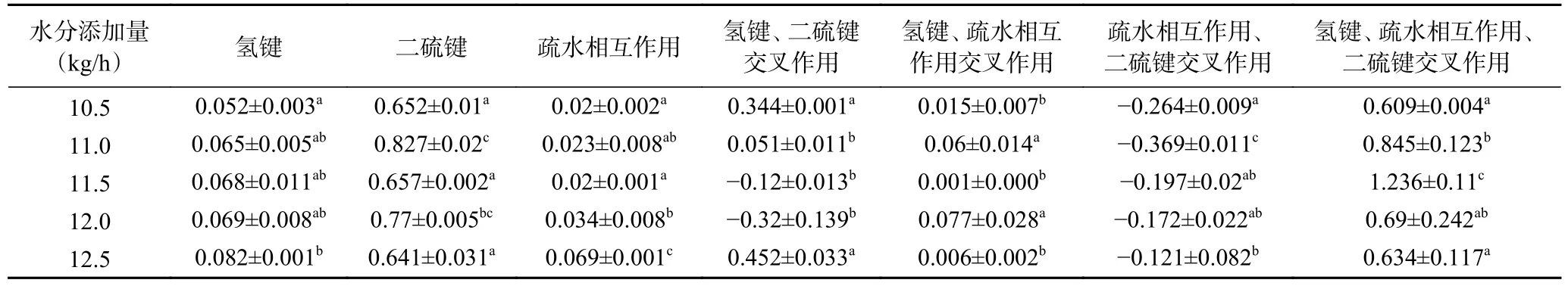
表3 水分添加量对植物蛋白肉化学交联变化的影响Table 3 Effect of water addition on the chemical cross-linking force of plant protein meat
综合上述结果可知,植物蛋白肉的结构由共价键与非共价键的交互作用共同维持。在所考察的水分添加量范围内,共价键与非共价键的交互作用对维持植物蛋白肉结构的贡献最大,非共价键作用和共价键作用次之,并且非共价键作用大于共价键作用,这与张金闯[14]的研究结果一致。
2.8 水分添加量对植物蛋白肉二级结构的影响
水分添加量对植物蛋白肉红外光谱图的影响如图4a 所示。在3700~3100 cm-1内,5 种水分添加量的植物蛋白肉均出现较宽的特征条带,这主要由O-H、N-H 伸缩振动和氢键伸缩振动引起的,在3000~2700 cm-1处出现的尖锐吸收峰,这归因于C-H 收缩振动[35]。植物蛋白肉在2500~2300 cm-1处出现的吸收峰主要与O=C=O 和R-N=C=O 伸缩振动有关[36],此处吸收峰强度随着水分含量的增加先增强后减弱,并且在水分添加量为11.0 kg/h 时达到最强,这表明植物蛋白肉的稳定性随着水分添加量的升高呈先升高后降低的趋势,这与前面热特性分析结果一致。而在1500~1700 cm-1处的吸收峰与美拉德反应产物如羰基化合物、希夫碱和吡嗪类相关的C=O、C=N 和C-N 伸缩振动有关[37],吸收峰强度均随着水分含量增加而减弱,表明挤压过程中水分含量对美拉德反应的发生程度具有一定影响。
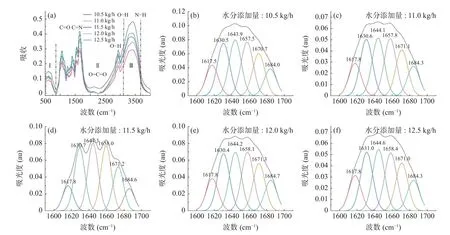
图4 水分添加量对植物蛋白肉二级结构的影响Fig.4 Effect of water addition on the secondary structure of plant protein meat
由图4a 可知,随着水分添加量的变化并未有新峰出现,但结合傅里叶自卷积、二阶导数和高斯曲线拟合(图4b~图4f),通过定量分析频谱的二阶导数发现蛋白质二级结构随着水分添加量的增加发生显著变化。根据文献[16]报道可知,α螺旋由分子内氢键维持,位于1646~1662 cm-1;β转角为松散有序结构,位于1662~1681 cm-1;无规则卷曲结构由C=O和H2O 之间形成的氢键维持,位于1637~1645 cm-1;β1 为反向平行式折叠的β层状结构,由分子间氢键维持,位于1608~1622 和1682~1700 cm-1。β2 为平行式折叠的β层状结构,由分子内的氢键维持,位于1622~1639 cm-1。
水分添加量对植物蛋白肉二级结构的影响如表4 所示。由表4 与图4b~图4f 可知,随着水分添加量的增加,植物蛋白肉的二级结构分别出现在不同位置,并且具有不同峰面积。β折叠为植物蛋白肉二级结构的主要作用力,其中α螺旋与β2 所占含量高于β1,表明分子内氢键作用力更强,这与Guo 等[16]研究的结果一致。随着水分添加量由10.5 kg/h 增加到11.5 kg/h 时,β1、β2 与α螺旋含量均呈降低趋势,这表明植物蛋白肉中维持β1、β2 与α螺旋结构的氢键作用力减弱,这可能与水分含量的突然增大有关。结合DSC 与化学交联结果可知,物料水分添加量的突然增加会使蛋白分子结构变得灵活,其分子结构更易于被打开[26],从而使植物蛋白肉的热稳定性降低。此外,当水分添加量由10.5 kg/h 增加到11.5 kg/h时,β转角和无规则卷曲分别呈现出先升高后降低与先降低后升高趋势,这表明在适当的挤压水分条件下,分子二级结构中松散有序结构逐渐增加,这有利于β折叠、α螺旋、无规则卷曲结构向β转角的转化。当继续增大水分含量时,加剧了蛋白质内部疏水基团的暴露和蛋白质交联变性程度的增大,蛋白质形成了更多的膜状气孔结构,这与扫描电镜观察到的结果一致。
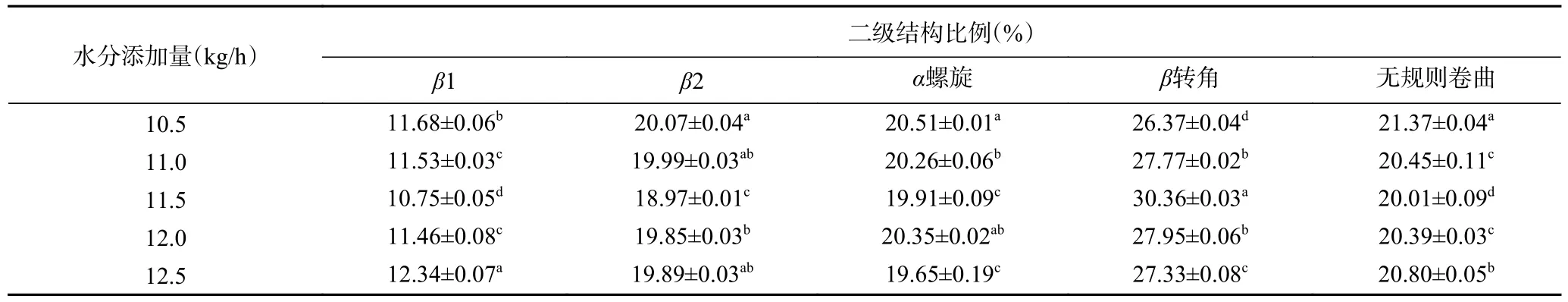
表4 水分添加量对植物蛋白肉二级结构的影响Table 4 Effect of water addition on the secondary structure of plant protein meat
2.9 植物蛋白素肉的相关性分析
对不同水分添加量植物蛋白肉的质构特性、色差、二级结构、变性热焓、化学交联键进行相关性分析,结果如图5 和表5 所示。从图5 可知,12.5 和12.0 kg/h 水分添加量与10.5、11.0、11.5 kg/h 水分添加量方向不同,并且11.5 kg/h 水分添加量与其他4 组所在象限均不同,表明水分含量对植物蛋白肉作用效果显著。同时从载荷可以看出,硬度与咀嚼度这两个特性彼此接近,表明它们之间存在正相关关系。此外,二硫键所处正交方向,表明其独立变化,对其他性能没有影响。由表5 可知硬度、咀嚼度、弹性指数与氢键、疏水相互作用、二硫键和疏水相互作用均呈负相关,而咀嚼度、弹性指数与α螺旋含量呈正相关。P<0.01 时,L*与β转角含量呈正相关,L*与氢键+二硫键呈负相关且皮尔逊系数最大。P<0.01 时,持水性与β1 呈负相关,P<0.05 时,持水性与β转角和氢键+二硫键+疏水相互作用呈正相关。持油性在P<0.01 时与疏水相互作用呈正相关且皮尔逊系数最大为0.743。

表5 植物蛋白肉的相关性分析Table 5 Correlation analysis of soybean meal simulated meat
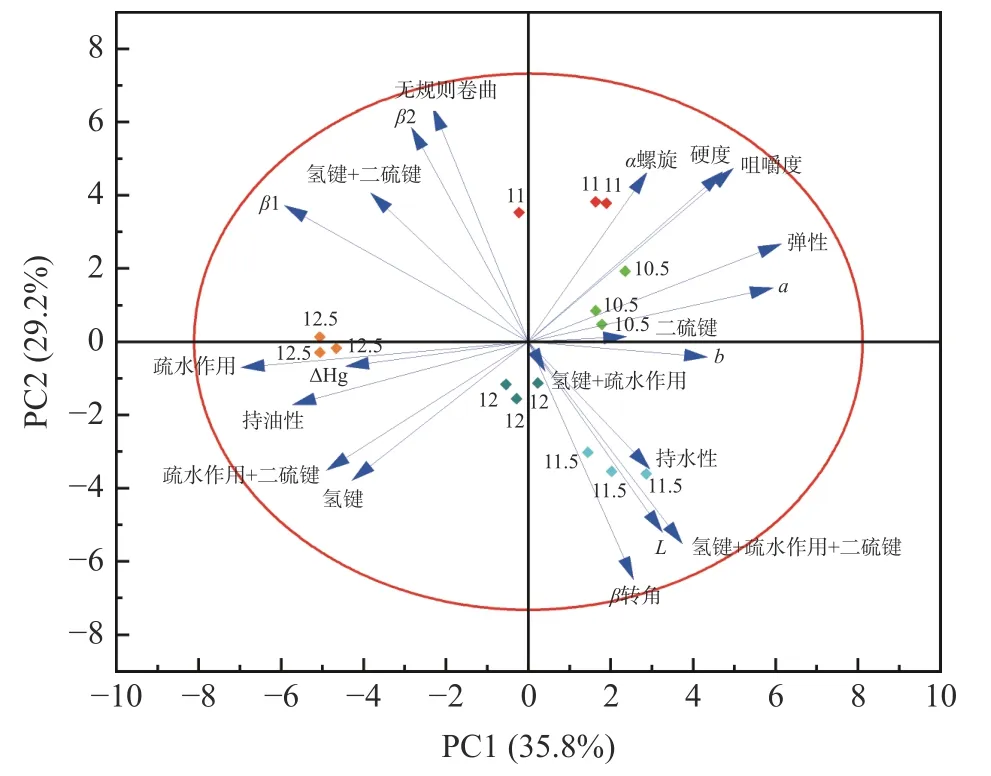
图5 植物蛋白肉的相关性分析载荷图、分值图及双标图Fig.5 Loading plot and scora plot and biplot correlation analysis of soybean meal simulated meat
综上所述,植物蛋白肉质构特性主要是化学交联力中的氢键、二硫键、氢键+疏水相互作用及二级结构中α螺旋共同作用的结果。二级结构中β1、β2 和β转角的变化对植物蛋白肉的亮度影响显著。此外植物蛋白肉持水性是β1、β转角、无规则卷曲、氢键+二硫键+疏水相互作用的结果,而持油性变化主要受疏水相互作用影响。
3 结论
本文以低温脱脂大豆粕为原料,探究了水分添加量对高水分挤压植物蛋白肉理化性质和微观结构的调控作用。研究结果表明,水分添加量作为挤压过程中的重要参数对植物蛋白肉的品质特性影响效果显著,随着挤压过程中水分添加量的增加,植物蛋白肉的硬度、弹性、咀嚼性均显著下降,而亮度值、持水性和持油性均呈先升高后降低趋势。水分添加量在11.5 kg/h 时,植物蛋白肉具有较好的品质特性与纤维结构。适当提高水分添加量有利于α-螺旋、β-折叠、无规则卷曲结构向β-转角的转化,同时促进共价键与非共价键的交互作用,对植物蛋白肉松散有序结构的增加,形成具有一定规则的晶型排列结构具有促进作用。高水分挤压植物肉具有更接近于动物肉的纤维结构和质构等特性,对其开发在减少全球碳排放、满足民众日益增加的肉类需求、减少动物性食品的消费以及确保中国和世界食物供给安全等方面具有重要意义。
猜你喜欢
食品科学技术学报(2022年5期)2022-10-11
质谱学报(2021年6期)2021-12-06
中国生物制品学杂志(2021年10期)2021-10-20
当代水产(2019年11期)2019-12-23
当代水产(2019年7期)2019-09-03
当代水产(2019年6期)2019-07-25
河南畜牧兽医(2017年8期)2017-11-24
中国饲料(2016年17期)2016-12-01
中学化学(2015年12期)2016-01-19
原子与分子物理学报(2014年3期)2014-02-28
