
水稻和陆稻分化相关性状的QTL分析及水旱不同栽培条件下连续选择的影响
2023-11-15 10:42张培风李合顺王学军
种子 2023年8期
孙 佩, 张培风, 李合顺, 王学军
(新乡市农业科学院, 河南 新乡 453000)
稻种的抗旱性是由多基因控制的数量性状,与许多生理性状、农艺性状密切相关。因此,要对稻种抗旱性机理进行系统研究,必须对与抗旱性相关的主要农艺性状进行QTL定位,分析各性状对抗旱性的贡献及遗传方式。水稻株高、抽穗期、剑叶宽是多基因控制的数量性状。Monna等[1],Sasaki等[2],Spielmeyer等[3]克隆了水稻绿色革命基因sd1,sd1位于1号染色体上,参与赤霉素的生物合成。Liu等[4]用籼粳组合IR64×Azucena的DH群体,共检测到15个株高QTL,其中QTL1-15是主效QTL,其余的是微效QTL。王韵等[5]利用粳稻Lemont和籼稻特青相互导入构建的遗传背景基本一致的双向回交导入系群体,分别检测到16个影响抽穗期的主效QTL和17个影响株高的主效QTL,鉴定出一些在不同遗传背景和环境下稳定表达的主效QTL,如QHd3、QHd8a、QPh3和QPh4,适宜用于水稻抽穗期和株高的分子标记改良。冯玉涛等[6]利用金23B和CR071构建了BC3F1群体及其衍生的BC3F2群体,共定位到了12 个抽穗期相关QTL,8个株高相关的QTL,其中抽穗期和株高最大效应QTL均由CR07提供,且都定位于7号染色体同一位置。侯磊磊等[7]用粳型品种月之光与籼型品种明恢63的F2群体进行了抽穗期和株高的QTL定位分析,共定位到4个抽穗期QTL,分布于1,6,8,12号染色体上,8号染色体上的QHd8是主效基因,与已克隆的DTH8在相近位置,可能是DTH8;定位到4个与株高相关的QTL,分别位于1,3,8,12号染色体上,1号染色体上检测到的qPH1是主效基因,位于矮秆基因sd1附近,可能是sd1。彭伟业等[8]以粳稻魔王谷和籼稻CO39组配的重组自交系为材料,检测到12个控制剑叶形态性状的QTL,分布于1,3,4,6,7,9号染色体上,只检测到1个控制株高的QTL,位于1号染色体上,是一个主效 QTL。Lin等[9]利用BC4F2群体和BC4F3群体精细定位到影响抽穗期的QTL:Hd9位于3号染色体短臂(C721-R1468B)。谭禄宾等[10]以中国云南元江普通野生稻与籼稻品种特青构建的BC3F2群体和BC3F3群体,检测到6个控制抽穗期的QTL,分别位于1,3,7,8,11号染色体上。董国军等[11]以剑叶角度差异显著的籼稻窄叶青8号和粳稻京系17构建DH群体为材料,分别在1,2,3,12号染色体上检测到4个剑叶角度QTL。张向阳等[12]利用籼稻材料千粒稻与粳稻材料日本晴构建的F2群体检测到3个剑叶宽QTL,分别位于1,2,7号染色体上。
大约在一万年前,人类社会就已经开始从单纯的狩猎和采集型向农业型转变,并且在四千年前,古代人类就已经完成了所有主要作物的驯化。经过漫长的自然选择和人工选择,栽培稻发生了一系列农艺性状和生理特性上的重大改变,形成了品种类型复杂、栽培要求不一的生态类型[13]。
本研究以云南地方品种(水稻和陆稻作亲本)所构建的RIL9群体进行抗旱性相关农艺性状的QTL定位,为认识这些性状的遗传方式提供理论依据和实践基础。双亲所构建的F2群体通过Bulk方法进行繁殖,从F4代开始水、旱田两种栽培方式连续选择5代,分别得到CA、CB两个群体,调查抗旱性相关的农艺性状,以阐明人工选择的影响,为抗旱育种提供参考。
1 材料与方法
1.1 试验材料
双亲为云南省石林彝族栽培稻田Ch55群体中的2个个体:Ch5-10和Ch6-11(OryzasativaL.)。Ch5-10(母本、水稻、偏籼性)田间表现为茎秆粗壮、分蘖力较强、抽穗期较长。Ch6-11(父本、陆稻、偏粳性)田间表现为茎秆细弱、分蘖力较弱、抽穗期较短。双亲植株均高大繁茂,但株高差异明显。
双亲所构建的F2群体利用SSD方法得到RIL9群体(113个株系),进行QTL分析。
双亲所构建的F2群体利用Bulk方法进行繁殖。F4代开始进行水、旱田两种条件下种植。经过5代的筛选后,繁殖成株系,分别得到CA(289个株系)和CB(332个株系)两个群体。
1.2 表型鉴定
RIL9群体:试验于2009年分别在中国农业大学北京上庄试验站和安徽省农业科学院水稻研究所(合肥)进行,2010年在中国农业大学北京上庄试验站进行。主要考察始穗期、株高、剑叶宽等3个性状,每个株系选取有代表性的10株调查。
CA、CB群体:试验于2009年分别在中国农业大学北京上庄试验站和安徽省农业科学院水稻研究所(合肥)进行。主要考察最高分蘖数、有效分蘖数、始穗期、株高和剑叶宽等5个性状,每个株系选取有代表性的10株调查。
按照30 cm×30 cm的规格种植,每个株系各种植2行,每行12株,田间管理同常规大田。评价标准如下:
最高分蘖数:在拔节期间隔一周调查分蘖数,确定最高峰时的分蘖数。
有效分蘖数:在黄熟期时调查有效分蘖数(包括主茎),凡未抽穗或抽穗而不结实的为无效分蘖。
始穗期:10%的穗顶部露出剑叶鞘时为始穗期。
株高:黄熟期时,测量单株自地面到穗顶(不连芒)的高度(每株株高以该株最高穗的高度为准)。
剑叶宽:黄熟期时,主茎剑叶宽。
1.3 DNA提取及SSR分析
参照Rogers等[14]的CTAB法略加改动提取DNA。利用从Panaud等[15]、Chen等[16],Temnykh等[17]和McCouch等[18]所构建的高密度水稻连锁图谱中选出均匀覆盖12条染色体的660对SSR标记,及Wang等[19]开发的20对ILP标记,检测双亲多态性。
1.4 遗传图谱的构建及QTL分析
利用Map Manager QTXb20软件(Manly等[20];http://mapmgr.roswellpark.org/mmQTL.html)区间作图法进行分析。遗传图谱包括115对SSR标记及8对ILP标记。取概率值小于0.01作为判断QTL存在的阈值。采用SPSS17.0软件进行数据统计分析。
2 结果与分析
2.1 RIL9群体的QTL定位分析
2.1.1RIL9亲本及群体的表型变异与相关性分析
由表1可知,在始穗期方面,双亲在北京地区种植差异明显,Ch5-10和Ch6-11在2009年分别为119.0 d和80.0 d,2010年分别为123.0 d和90.0 d。双亲在安徽地区种植差异较小,Ch5-10和Ch6-11在2009年分别为98.0 d和95.0 d。双亲2009年北京剑叶宽差异达到极显著水平,Ch5-10和Ch6-11分别为1.6 cm和1.4 cm,2010年北京剑叶宽双亲间差异不显著,均为1.8 cm。株高双亲间差异达到极显著水平,2009年北京Ch5-10和Ch6-11分别为179.4 cm和128.9 cm,2009年安徽Ch5-10和Ch6-11分别为190.0 cm和160.0 cm,2010年北京Ch5-10和Ch6-11分别为200.5 cm和146.7 cm。从分离群体变异的分布范围来看,各性状均出现一定数量的超亲类型,而且多数是双向超亲分离。经单样本K-S检验,各性状均符合正态分布。

表1 亲本及RIL9群体的表型统计结果
田间表型间相关性如表2所示。极显著正相关的性状有20对,包括始穗期×株高、始穗期×剑叶宽、株高×剑叶宽;显著正相关的性状有2对,包括2009年安徽始穗期×2009年北京株高、2010年北京株高×2010年北京剑叶宽。

表2 田间表型的相关性分析
2.1.2遗传图谱的构建
本研究挑选均匀覆盖水稻基因组的680对标记检测双亲多态性。在作图群体中能稳定扩增出多态性条带并最终统计数据的有123对,构建了覆盖水稻12条染色体的遗传连锁图,部分染色体见图1。各染色体多态性标记数为6(10号染色体)~16对(3号染色体)。
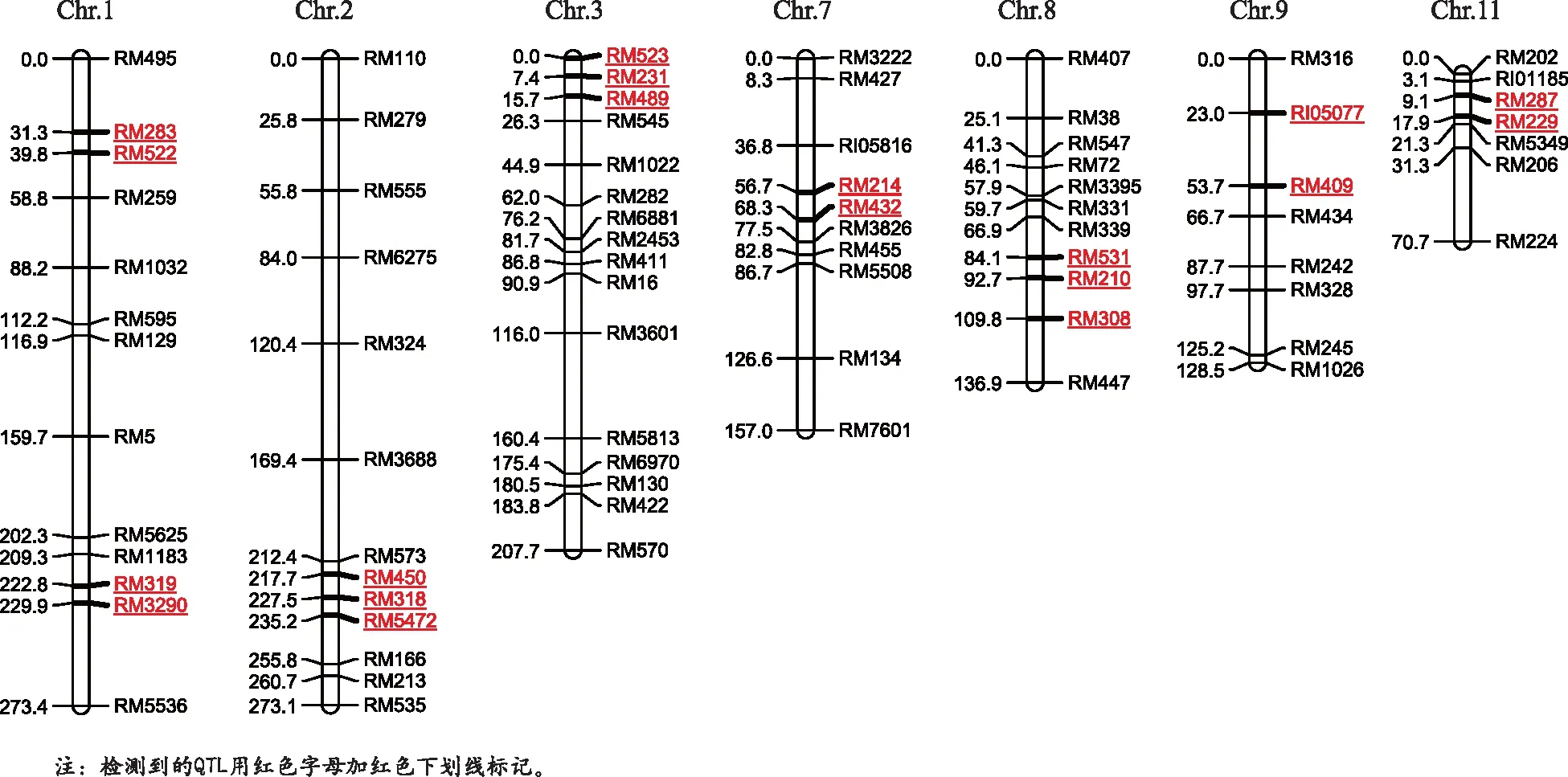
图1 检测到的QTL在染色体上的分布
2.1.3QTL分析
3个性状共检测到19个QTL(p<0.01),分布于1,2,3,7,8,9,11号染色体上。2010年北京始穗期的QTL最多,为4个。2009年北京剑叶宽的QTL最少,为1个。各性状QTL的数目和位置、效应见表3和图1。

表3 QTL分析结果
始穗期:2009年在北京种植的群体共检测到2个QTL,分别位于3号和7号染色体上。qHD-3-1位于3号染色体RM523-RM231标记区间,LOD值为3.4,贡献率为15%。qHD-7位于7号染色体RM214-RM432标记区间,LOD值为2.9,贡献率为11%。同时,对2009年在安徽合肥种植的群体分析共检测到了2个QTL,分别位于3号和7号染色体上。qHD-3-2位于3号染色体RM231-RM489标记区间,LOD值为4.7,贡献率为18%。qHD-7位于7号染色体RM214-RM432标记区间,LOD值为3.2,贡献率为12%。对2010年在北京种植的群体进一步分析,结果表明,在这一环境中检测到4个QTL,分别位于3,7,8号染色体上。qHD-3-1位于3号染色体RM231-RM489标记区间,LOD值为3.2,贡献率为12%。qHD-7位于7号染色体RM214-RM432标记区间,LOD值为2.6,贡献率为10%。qHD-8-1和qHD-8-2分别位于8号染色体RM531~RM210和RM210-RM308标记区间,LOD值分别为2.8和2.5,贡献率分别为11%和10%。
综合以上结果表明,qHD-7可以在 3个不同的生长环境中稳定表达。3号染色体的位点为主效QTL,但稳定性较差,表现为2009年安徽和2010年北京结果一致,2009年北京检测到的位置在相邻区间。3号和7号染色体的QTL均表现为来自亲本Ch5-10的基因增加始穗期。2010年北京在第8染色体检测到的2个QTL表现为来自亲本Ch6-11的基因增加始穗期。
株高:2009年在北京种植的群体共检测到4个QTL,分别位于1,3,9,11号染色体上。qPH-1位于1号染色体RM319-RM3290标记区间,LOD值为4.4,贡献率为16%。qPH-3位于3号染色体RM231-RM489标记区间,LOD值为2.6,贡献率为10%。qPH-9位于9号染色体RI05077-RM409标记区间,LOD值为2.4,贡献率为9%。qPH-11位于11号染色体RM287-RM229标记区间,LOD值为2.4,贡献率为10%。同时,对2009年在安徽合肥种植的群体分析共检测到了3个QTL,分别位于1,2,3号染色体上。qPH-1位于1号染色体RM319-RM3290标记区间,LOD值为6.2,贡献率为22%。qPH-2-1位于2号染色体RM318-RM5472标记区间,LOD值为2.4,贡献率为9%。qPH-3位于3号染色体RM231-RM489标记区间,LOD值为2.4,贡献率为9%。对2010年在北京种植的群体进一步分析,结果表明,在这一环境中检测到3个QTL,分别位于1,2,3号染色体上。qPH-1位于1号染色体RM319-RM3290标记区间,LOD值为4.4,贡献率为16%。qPH-2-2位于2号染色体RM450-RM318标记区间,LOD值为2.6,贡献率为10%。qPH-3位于3号染色体RM231-RM489标记区间,LOD值为2.5,贡献率为10%。
综合以上结果表明,qPH-1为主效QTL,qPH-1和qPH-3可以在 3个不同的生长环境中稳定表达。其他的位点均表现为只在某一环境中表达,稳定性差。各QTL均表现为来自亲本Ch5-10的基因增加株高。
剑叶宽:只在一种环境中检测到QTL(2009年北京),qFLW-1位于1号染色体RM283-RM522标记区间,LOD值为3.2,贡献率为12%。表现为来自亲本Ch5-10的基因增加剑叶宽。
2.2 CA和CB群体水旱不同栽培条件下连续选择的影响
2.2.1CA和CB亲本及群体的表型
由表4可以看出,在最高分蘖数方面,双亲在2009年北京地区种植差异达到极显著水平,Ch5-10和Ch6-11分别为19.8个和14.0个。在有效分蘖数方面,双亲在2009年北京地区种植差异不显著,Ch5-10和Ch6-11分别为8.6个和8.1个,其他性状双亲之间差异水平同表1。群体中各性状基本符合正态分布,CA群体和CB群体间做t检验检测,发现2009年安徽始穗期和2009年北京剑叶宽差异不显著,其他性状差异均达到极显著水平。

表4 亲本及CA、CB群体的表型统计结果
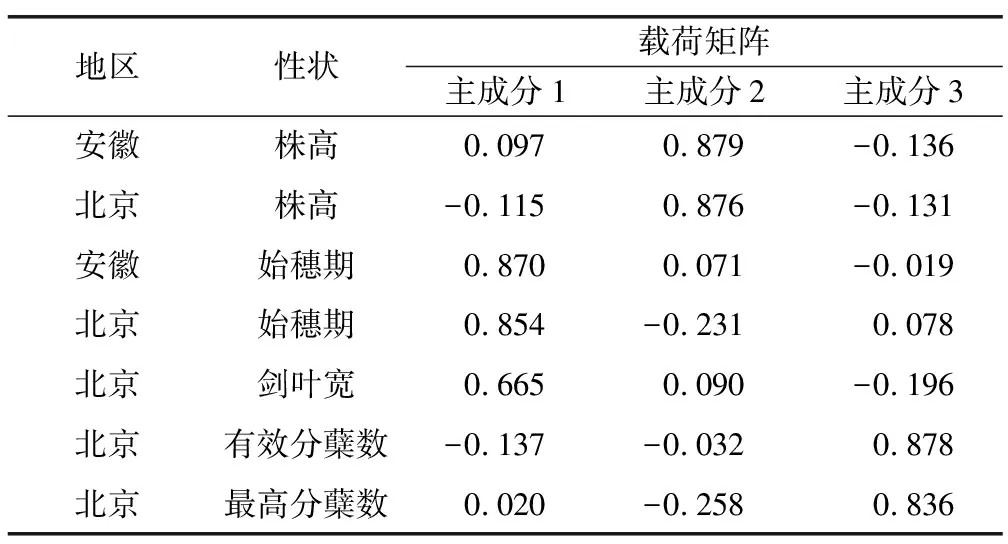
对CA群体和CB群体各性状平均数作比较,北京最高分蘖数:CA>CB;北京始穗期:CA>CB;北京有效分蘖数:CA>CB;北京剑叶宽:CA=CB;北京株高:CA 2.2.2农艺性状的因子分析 本研究运用SPSS17.0软件进行农艺性状的因子分析。按照特征根大于1.00的原则,提取3个主因子,其累积贡献率达74.2%,即反映了原有信息的74.2%。将这3个主因子作为评价综合变量。贡献率分别为28.1%,23.9%和22.1%。 表5是因子旋转成分矩阵,提取方法为主成分分析法,旋转法为具有Kaiser标准化的正交旋转法。第一个因子对北京始穗期、安徽始穗期和北京剑叶宽有绝对值较大的负荷系数,第二个因子对北京株高和安徽株高有绝对值较大的负荷系数,第三个因子对北京最高分蘖数和北京有效分蘖数有绝对值较大的负荷系数。根据因子分析结果作散点图(图2),可以看出CA、CB两群体经过5代的水、旱田筛选,最高分蘖数、始穗期、有效分蘖数、剑叶宽、株高有明显差异。 图2 CA、CB群体的因子分析 表5 旋转成分矩阵 2.2.3基因型分析 根据RIL9群体的QTL定位结果[21],选择qSLtr-2、qSLratio-2、qRL28-2-1和qRL28-4附近的标记RM166、RM5472、RI05751和RM5979,及未定位到QTL的对照标记RM259对CA群体和CB群体进行基因型分析,两群体间t检验结果表明,RM5472、RI05751和RM5979均达到极显著水平,RM259和RM166差异不显著。其中,RM166差异不显著,可能是RM166-RM5472区间较大,RM166与QTL位点距离较远所致。 本研究以来自云南的地方品种Ch5-10(母本、水稻、偏籼性)和Ch6-11(父本、陆稻、偏粳性)为亲本,后代用SSD方法而得到的RIL9群体为试验材料,对水陆稻差异性状(始穗期、剑叶宽、株高等)进行了考察。并利用SSR分子标记构建连锁图谱,进行了上述性状的QTL定位。双亲杂交得到F2代,后代用Bulk方法进行繁殖。F4代开始对Bulk群体进行水、旱田两种条件种植。经过5代的筛选后,繁殖成株系,分别得到CA、CB两个群体。对水陆稻差异性状(始穗期、最高分蘖数、有效分蘖数、剑叶宽、株高)进行考察。 水稻株高的QTL定位研究表明,1号染色体上与sd1基因处于相同位置的QTL最常被检测到,效应值也较大[1-3]。本研究株高主效QTL:qPH-1与sd1位置相近。Liu等[4]利用一套来源于籼粳组合IR64×Azucena的DH群体定位到的株高QTL位点qPH-2-11与本研究qPH-2位置一致。王韵等[5]利用粳稻Lemont和籼稻特青相互导入构建的遗传背景基本一致的双向回交导入系群体,定位到的株高QTL位点qPH-2与本研究qPH-2位置一致。冯玉涛等[6]利用金23B和CR071构建的BC3F1群体及其衍生的 BC3F2群体,定位到的株高QTL位点qPH9与本研究结果qPH-9位置一致。侯磊磊等[7]利用粳型品种月之光与籼型品种明恢 63的F2群体定位到的株高QTLqPH1和qPH3与本研究结果qPH-1和qPH-3位置一致。彭伟业等[8]以粳稻魔王谷和籼稻 CO39 组配的 280 个重组自交系为材料,定位到的株高QTLqPH1与本研究结果qPH-1位置一致。姜雪等[22]利用矮秆品种“V20B”和高秆品种“CPSLO17”作为亲本,构建了150个重组自交系家系(RILs),结合SLAF_seq技术构建的水稻高密度遗传图谱,对水稻株高进行QTL检测。在1号和6号染色体各检测到2个株高QTL位点,其中qPH1-2对表型变异的贡献率为22.2%,位于sd1基因附近,与本研究qPH-1位置一致。 冯玉涛等[6]利用金23B和CR071构建的BC3F1群体及其衍生的 BC3F2群体,定位到的抽穗期QTL位点qHD7与本研究结果qHD-7位置相近。Lin等[9]利用BC4F2群体和BC4F3群体精细定位到影响抽穗期的QTL:Hd9与本研究QTL定位结果qHD-3-1位置一致;谭禄宾等[10]以中国云南元江普通野生稻与籼稻品种特青构建的BC3F2群体和BC3F3群体,检测到的位于3号染色体RM22附近的抽穗期QTL位点与本研究QTL定位结果qHD-3-1位置一致。赵凌等[23]利用抽穗期存在明显差异的粳稻TD70和籼稻Kasalath杂交衍生的包含186个家系的重组自交系群体,构建了基于深度重测序的高密度Bin遗传图谱,对控制水稻抽穗期的QTL进行鉴定,其中qHD3与本研究qHD-3-1位置一致。 董国军等[11]以剑叶角度差异显著的籼稻窄叶青8号和粳稻京系17构建DH群体为材料,检测到的剑叶角度QTL的qFLA-1与本研究剑叶宽QTL定位结果qFLW-1位置相近。王小雷等[24]利用籼稻昌恢121和粳稻Koshihikari构建的208个染色体片段置换系对控制株高、剑叶形态和分蘖数的QTL进行检测,其中株高QTL的qPH-1-2与本研究qPH-1位置一致,叶宽QTL的qFLW-1与本研究qFLW-1位置一致。 陆稻亲本Ch6-11始穗期较短、分蘖数较少、剑叶宽偏窄、株高偏低,这些都表现为植株对干旱环境的一种适应性和积极的反应。从表2可以看出,始穗期的QTL的加性效应有正有负,表明了来自双亲的等位基因对这些性状值都有增效作用。等位基因聚合在一起互作,部分解释了群体的超亲分离现象。两个性状双亲的等位基因分布于不同染色体上,这为进行标记辅助选择和基因聚合育种提供了条件。聚集在一起的QTL可能表现为“紧密连锁”,也可能表现为“一因多效”[25]。例如3号染色体的RM231-RM489的区间内聚集着株高和始穗期在不同环境下检测到的5个QTL。CA、CB群体经过5代的水、旱田环境筛选之后,最高分蘖数、有效分蘖数、始穗期、株高、剑叶宽有明显差异。说明两群体的表型和基因型都受到了人工选择的影响。
3 结论与讨论
猜你喜欢
中国水稻科学(2021年6期)2021-11-18
东坡赤壁诗词(2020年5期)2020-11-06
农村百事通(2019年17期)2019-10-08
浙江农业科学(2019年1期)2019-01-25
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
小学生时代·综合版(2016年4期)2016-11-19
扬子江(2016年1期)2016-05-19
化工进展(2015年3期)2015-11-11
